Демонстративное поведение школьников: причины, проявления, способы коррекции
Демонстративное поведение школьников: причины, проявления, способы коррекции
АКТУАЛЬНОСТЬ ЭТОЙ ТЕМЫ заключается в том, что демонстративность, как нарушение поведения — это очень часто встречающаяся, как личная особенность подрастающего поколения нашего времени, которая может как помочь личности в достижении значимых для неё целей, так и создать множество проблем в социальных и межличностных отношениях, в частности в образовательных учреждениях и семье.
Демонстративное поведение – подчеркнуто-выразительное поведение с осознаваемой или неосознанной целью привлечь к себе особое внимание окружающих без учёта их потребностей и желаний.
Демонстративность- личная особенность. В её основе лежит повышенная потребность во внимании к себе, стремление всегда быть в центре внимания.

Причины возникновения: (по слайду)
Недостаток внимания взрослых к детям
По наблюдениям и диагностическим данным за детьми можно выделить четыре основных мотива » демонстративного » поведения.
Очень часто причиной такого поведения учеников бывает потребность в особом внимании к себе.
Мы все нуждаемся во внимании со стороны окружающих людей и не хотим чувствовать себя «пустым местом», мы хотим ощущать свой вклад в группу, к которой себя причисляем. Однако есть дети, которые своим поведением все время стремятся привлечь внимание и никогда не удовлетворены получаемым вниманием, требуя его все больше и больше.
Возникает вопрос, что чувствует учитель, наблюдающий это на своём уроке. Что чувствует нормальный человек, которого всё время отвлекают? Да, совершенно верно — раздражение. Из него легко рождается негодование и возмущение.
Что чувствует нормальный человек, которого всё время отвлекают? Да, совершенно верно — раздражение. Из него легко рождается негодование и возмущение.
Отсюда следующий вопрос — что хочется сделать? Сделать замечание. Иногда прочитать нотацию.
Какова же ответная реакция ученика. Временно прекращает выходку (обратите на это внимание, если ученик не прекращает вертеться или стучать после Вашего замечания — мотив другой), их мы рассмотрим дальше.
Итак, дети привлекают внимание на уроке и основная трудность педагога — как дать внимание (ведь они имеют на него право), не подкрепляя при этом условный рефлекс «хочешь внимания — делай плохо».
Постарайтесь до урока подчеркнуть громко «что-то» хорошее в этом ученике, предложить выполнить поручение: достать мел, написать число т. п. (полить цветок) и похвалить.
Следующий мотив — власть
Одна из главных причин такого поведения — мода на «сильную» личность, на «крутых». Дети и подростки видят только внешнюю эффективную сторону властолюбивого поведения. Они ещё не знают того, что знают люди мудрые: против лома нет приёма, окромя другого лома. Страх — оборотная сторона власти.
Дети и подростки видят только внешнюю эффективную сторону властолюбивого поведения. Они ещё не знают того, что знают люди мудрые: против лома нет приёма, окромя другого лома. Страх — оборотная сторона власти.
Многие молодые родители полагают, что «крутой» и есть сильная личность. А сильная личность — это человек, который принимает решения за других.
Однако сильная личность ничего не решает за других, а просто не позволяет другим решать за неё. Для этого не нужно быть агрессивным, достаточно быть уверенным в себе.
Рассмотрим чувства учителя, наблюдающего властолюбивое поведение: гнев, негодование, возмущение, часто также страх.
Действия учителя — что же он хочет сделать? Этот момент особенно важен для различия власти и привлечение внимания. Властолюбца хочется «убить своими руками», то есть, говоря методически правильно, немедленно прекратить выходку. Иными словами, когда кулаки сжимаются — имеешь дело с мотивом «власть».
Какова ответная реакция ученика. Эта реакция также весьма характерна — ученик прекратит выходку, когда сам решит. Он провоцирует Вас, вы попадаетесь на провокацию, выходите из себя — именно этого он и добивался.
Он победил и наслаждается минутами торжества. Почему же он должен прекращать?
Конечно, можно забыть про гуманизм и унизить или напугать его. Но тогда Вы сделаете из него мстителя. А «месть» — более тяжёлый, чем «власть» мотив «плохого» поведения.Месть
Месть похожа на власть, но причины «плохого» поведения здесь глубже. Если властолюбец просто испытывает вас, то мститель изживает внутреннюю боль, которую нанесли, может быть, вы сами, не заметив этого, а скорее всего, кто- то другой.
Непрофессиональное и неэтическое поведение педагога часто также является причиной мести.
Первоклассник полгода отчаянно мстил хорошей учительнице, пока она не решилась выяснить причину его ненависти. Он сказал: » Вы всех посылали за журналом, даже Мишку, хоть он глупее меня. А меня не разу!».
Он сказал: » Вы всех посылали за журналом, даже Мишку, хоть он глупее меня. А меня не разу!».
Мстители делают не просто пакости, они выбирают то, что произведёт впечатление именно на этого человека. Если учительница любит цветы можно «нечаянно» уронить цветочный горшок, остричь их ножницами.
Если требует пунктуальности — мститель будет методично опаздывать на каждый урок. Если учительница много говорит о приличном внешнем виде, можно специально соорудить костюмчик, увидеть который, она не останется равнодушной.
Каковы здесь чувства учителя. Это растерянность, гнев, негодование, обида, страх. «Как это вообще могло произойти?»
Что хочется сделать учителю? «В этой школе, дорогой, останешься либо ты, либо я».
Ответно мстить или убегать — древняя, первобытная формула. Она включается в момент опасности, и изменить её крайне трудно.
Учитель в эти минуты забывает о гуманизме, о том, что перед ним ещё ребёнок, о том, что из двоих участников конфликта именно он профессиональный воспитатель с дипломом.
Какова ответная реакция ученика. Она та же, что и у властолюбца.
Особая проблема — как изменить поведение мстителей? Личная проблема учителя: как научиться прощать учеников, которые им мстят, как вообще относится к ним?
Одна из психологических тонкостей здесь следующая. Нужно разделять личность (в данном случае человека, который пытается выжить, выражая внутреннюю боль с помощью агрессии) и поведение, то есть человека, и то что он сказал. Проступок, очевидно, нуждается в санкциях, а вот личность — в поддержке.
Избегание неудачи
Эти тихие на уроках дети вполне послушны, только не выполняют заданий. Происходит это потому, что они боятся вновь столкнуться с ощущением полного ничтожества: ведь получается у всех кроме меня!
У детей этот мотив формируется в первые годы обучения в школе. Постоянное ощущение неуспеха, что ты не оправдал чьих — то ожиданий: «Опять не успел сделать примеры! Почему «4» , а не «5»? Только ты один, Петров, опоздал в школу!» и т.
Какие чувства возникают у учителя в данной ситуации? Сначала это жалость, затем, когда раз за разом попытки позаниматься дополнительно, объяснить материал ещё раз ни к чему не приводят, возникает ощущение профессиональной беспомощности.
Что хочется сделать учителю, чтобы исправить ситуацию?
Хочется отвести ребёнка к психологу и получить ответ на вопрос «Почему он не тянет?». А психолог пишет: «Интеллектуальное развитие соответствует возрасту». А вот мотивация и эмоциональная сторона — запущены в процессе обучения.
Какова ответная реакция ученика. Ученик впадает в зависимость. Он готов на всё: мыть класс, носить парты, и т. д., но он по-прежнему не выполняет задания.
Сильные и слабые стороны демонстративного поведения (слайд)
Как и у любой личностной черты человека, у демонстративности есть свои сильные и слабые стороны. Давайте вместе подумаем, какие сильные стороны у такого поведения, а какие слабые?
Рассмотрим принципы профилактики демонстративного поведения (слайд)
Работа с учащимися (слайд)
Работа с родителями (слайд)
Работа с учителями (слайд)
Можно выделить несколько стратегий педагогического воздействия на учащегося с демонстративным поведением.
1 Стратегия «Минимизация внимания»
Это парадоксально: естественным образом реагируя на выходки учеников, целью которых является привлечение внимания, мы как бы даем им положительное подкрепление в виде нашего внимания, закрепляя тем самым их плохое поведение. Они начинают ощущать себя частью учебного процесса, чувствовать принадлежность к группе (классу) только когда получают много замечаний, а наши гневные проповеди и угрозы они воспринимают как вид особого внимания к себе. Зачем же поддерживать с помощью внимания такое поведение, которое нарушает ведение урока и отвлекает других учеников? Стратегия минимизации внимания содержит множество приемов, призванных уменьшить демонстративное поведение, поскольку оно будет оставаться незамеченным.
Они начинают ощущать себя частью учебного процесса, чувствовать принадлежность к группе (классу) только когда получают много замечаний, а наши гневные проповеди и угрозы они воспринимают как вид особого внимания к себе. Зачем же поддерживать с помощью внимания такое поведение, которое нарушает ведение урока и отвлекает других учеников? Стратегия минимизации внимания содержит множество приемов, призванных уменьшить демонстративное поведение, поскольку оно будет оставаться незамеченным.
Игнорируйте такое поведение. Часто лучший способ прекратить демонстративное поведение – перестать на него реагировать. «Нет ответа» — значит, цель не достигается с помощью этого действия. Когда ученик ведет себя демонстративно, задайте себе вопрос: «Что будет, если я проигнорирую его выходку?» Если вы ответите себе, что ничего не случится, кроме того, что он лишится вашего внимания, смело применяйте прием игнорирования. Через несколько попыток ученик прекратит вести себя подобным образом.
Установите зрительный контакт. «Пристально посмотрите на них, — советуют опытные учителя. — Они знают, что они делают. Они знают, что я знаю, что они делают. И они знают, что этот взгляд означает «хватит». Пристальный взгляд (без осуждения) — это все внимание, которое они «выручили» за свою проделку. Никаких слов — только взгляд».
Встаньте рядом. Физическое приближение — другой инструмент, помогающий минимизировать поведение, направленное на привлечение внимания. Продолжая вести урок, просто подойдите и встаньте рядом с учеником. Без контакта глазами и без слов. Дети начинают понимать, что делают что-то не то, когда учитель стоит так близко.
Упоминайте имя ученика. Этот прием позволяет одновременно дать минимум внимания «в награду» за демонстративное поведение и рекомендовать ученику присоединиться к тому, что вы объясняете. Учитель делает это, периодически вставляя имя ученика в контекст урока. Это может звучать так: «Таким образом, квадрат гипотенузы, Вова, равен сумме…» Или: «Тогда, Игорь, Петр Первый издал указ о…».
Эти простые приемы могут быть легко использованы с учениками, часто нарушающими поведение.
Пошлите «секретный сигнал». Вы можете использовать какие-то жесты, смысл которых известен детям. Например, приложить палец к губам и сказать: «Тсс». Вот интересный рассказ одной учительницы.
Однажды, разговаривая с второклассником Борей, который очень раздражал меня на уроке, я поинтересовалась: «Что мне сделать, чтобы ты спокойно сидел, пока я занята другими детьми?» Боря поднял над головой скрещенные руки, как это делает баскетбольный судья. «Что это значит?» — спросила я. «Это означает, что игра окончена», — ответил Боря.
Всегда и везде теперь, когда Боря шалил, я делала ему этот знак — и он помогал! К этому основному сигналу я часто добавляла подмигивание, и Боря улыбался и успокаивался, удовлетворенный тем, что получил хотя бы две секунды моего внимания только себе. Прошло время, Боря вырос. Но где бы мы ни встречались с ним, в коридоре или на школьном дворе, я подмигивала ему и скрещивала руки — это были наши особые отношения.
Делайте письменные замечания. Если вы знаете, что завтра у вас урок в классе, где будет ученик с поведением, направленным на привлечение внимания, приготовьте заранее стопку одинаковых записок с таким содержанием: «Пожалуйста, престань делать то, что ты сейчас делаешь». Просто кладите записку на парту ученику, когда он «разошелся». Не надо ничего говорить – все написано. Этот прием, конечно, работает с учениками, которые хорошо и быстро читают.
Формулируйте «Я-высказывания». Бывают ситуации, когда нервы не выдерживают и просто хочется крикнуть ученику, который заводит класс: «Прекрати немедленно!»
Психологи советуют в этих случаях применять «Я-высказывание». Это словесное утверждение дает в специфической форме информацию о нарушении поведения и о том, какое оно производит на вас впечатление. Вот пример: «Катя, когда ты шепчешься с Леной во время моего объяснения, я чувствую сильное раздражение, потому что теряю мысль. Пожалуйста, перестань!»
Пожалуйста, перестань!»
«Я-высказывание» состоит из трех частей:
Часть 1. Содержит объективное описание плохого поведения, которое имеет место здесь и сейчас: «Когда ты шепчешься с Леной во время объяснения…»
Часть 2. Называет чувства учителя в этот момент: «…я чувствую сильное раздражение…»
Часть 3. Описывает эффект от плохого поведения: «…потому что я теряю мысль…»
«Я-высказывание» сообщает ученикам только то, что мы чувствуем. Если, используя «Я-высказывание», вы будете искренни на словах и в интонациях, вы сможете подействовать на многих учеников.
2 Стратегия. «Разрешающее поведение»
Запретный плод всегда сладок. Это свойство человеческой натуры известно со времен Адама и Евы. Поэтому вторая стратегия заключается не в том, чтобы запретить есть яблоко, а в том, чтобы уничтожить всю прелесть содеянного, объявив яблоко не запретным.
Вот конкретные приемы стратегии разрешающего поведения.
Стройте урок на основе вопиющего поведения. Вот пример такого урока математики.
Ученики четвертого класса помешались на плевках из трубочек. Они не поддавались никаким воспитательным методам. Плевались на уроках и переменах. Тогда учитель математики сказал, что раз они так любят плевки, он собирается учить их математике исключительно на плевках. Он сделал мишень и движущийся контур, начертил на полу зоны, обозначающие разную степень успешности, и велел тщательно записывать результаты своих достижений. Довольно скоро дети больше не хотели и слышать о плевках.
Этот прием хорошо работает с ругательствами, которые могут быть изучены в теме «Сленг», с надписями на английском языке (названия рок-групп и молодежных лозунгов) — в английском, с выкриками запомнившихся рекламных слоганов — в обучении орфографии, пунктуации и практической стилистике.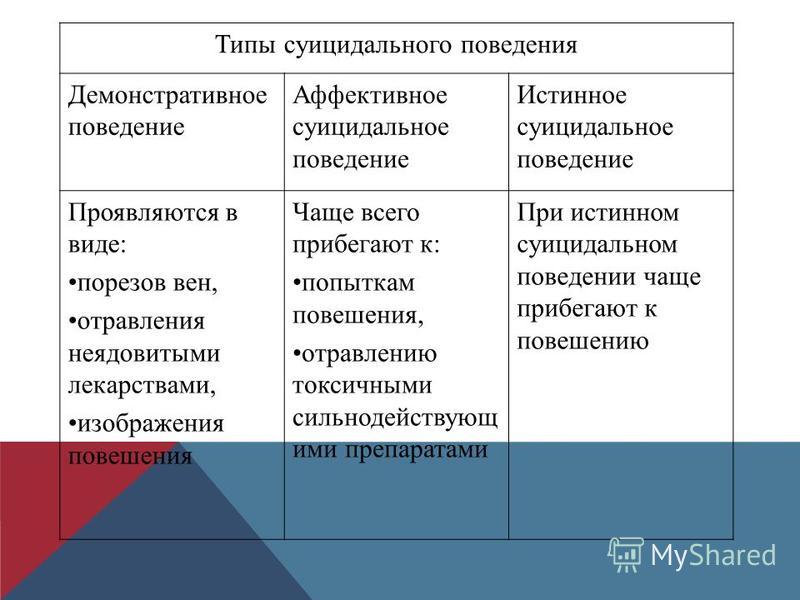
Доведите до абсурда демонстративную выходку. А вот другой прием, используемый в той же ситуации массовой эпидемии плевания.
Провинившиеся ученики получали задание плюнуть 500 раз. После первой сотни плевков они уже мечтали только об одном — никогда больше не видеть трубочек для плевания, ибо это было скучное и однообразное занятие, от которого сохло горло.
Этот прием годится для многих хамских выходок: учеников, которые не хотят сидеть на своем месте, заставьте стоять весь урок, ученики, которые постоянно болтают или издают неприличные звуки, должны записать полную часовую аудиокассету своих речей или звуковых упражнений и принести завтра в класс и т.п.
Ира без конца стучит ручкой по парте. Учитель: «Ну, хорошо. Класс, возьмите свои ручки и три минуты стучите ими по парте. Начали!» Весь класс присоединяется к выходке. Ира стучит со всеми и не получает какого-то особого внимания. И интерес к этому виду деятельности у Иры пропадает.
Используйте «разрешенную квоту». Этот прием рекомендует доктор Рудольф Дрейкурс в книге «Психология в классе». Прием состоит в том, что какое-то нарушение поведения разрешается, уж если оно появилось, но только в том объеме, который оговорен заранее и с условием, что ежедневно объем этот будет уменьшаться.
В качестве примера того, как работает этот прием, Дрейкурс описывает следующий случай.
Джонни громко икает на каждом уроке гражданского права по меньшей мере десять раз. Однажды утром, перед началом занятий, учитель договорился с Джонни, какое количество «иканий» будет позволено ему на каждом уроке, а также, что каждый день он будет икать меньшее число раз, чем вчера. А уж учитель проследит за этим. Каждый раз, когда Джонни икал, учитель просто улыбался ему и делал пометку мелом в углу классной доски. Когда «лимит» на сегодня был исчерпан, учитель говорил Джонни: «На сегодня — все!» Так постепенно число громких иканий было сведено на нет.
Скептики, возможно, скажут: «А что будет, если ученик продолжит выходку после слов учителя «На сегодня — все»?» Если это случится, оставьте эту технику и используйте другую технику из этой главы. Или же проанализируйте еще раз это нарушение поведения с точки зрения его цели — возможно, дело в том, что истинная цель не привлечение вашего внимания, а, например, власть. Техника «разрешенной квоты» работает только в отношении поведения, направленного на привлечение внимания.
Учителя, которые использовали эту технику, утверждают, что ученики соблюдают правило «разрешенной квоты». Почему? Потому что особые’ отношения между вами и учеником, ваши улыбки, отметки мелом — все это знаки внимания, которые так нужны ученику. Ведь эти ученики не нуждаются в утверждении своей власти над нами, они не хотят свергать ничьих авторитетов. Им нужно лишь немного внимания, чтобы не чувствовать себя «пустым местом».
3 Стратегия. «Поступайте неожиданно!»
Часто мы можем прекратить «плохое» поведение учеников, действуя неожиданно. Когда мы что-то неожиданно «выкидываем», мы как бы говорим: «Я все вижу и знаю, что ты делаешь, но не собираюсь играть в твою игру». Игра требует по меньшей мере двух участников. Когда же учитель отказывает в игре, лучше делать это необычным способом. Скажем, короткие вспышки хохота лучше, чем что-либо другое, могут разрядить атмосферу в классе. Чем больше юмора в вашем классе в момент нарушения поведения, тем быстрее оно прекратится. Вот некоторые конкретные приемы данной стратегии.
Когда мы что-то неожиданно «выкидываем», мы как бы говорим: «Я все вижу и знаю, что ты делаешь, но не собираюсь играть в твою игру». Игра требует по меньшей мере двух участников. Когда же учитель отказывает в игре, лучше делать это необычным способом. Скажем, короткие вспышки хохота лучше, чем что-либо другое, могут разрядить атмосферу в классе. Чем больше юмора в вашем классе в момент нарушения поведения, тем быстрее оно прекратится. Вот некоторые конкретные приемы данной стратегии.
Выключите свет. Это старый испытанный способ, который учителя используют давно. Когда кто-то из учеников или весь класс стал неуправляемым, просто поверните выключатель, подождите тишины несколько минут. Нет необходимости в очередной раз читать обвинительную речь или лекцию о том, что ученики плохо себя ведут, ведь они и сами прекрасно понимают, что нарушают правила поведения. В этой ситуации молчание дает больший эффект, чем громкие слова.
Используйте музыкальные инструменты. Учителя музыки часто исполняют аккорд или несколько аккордов на пианино в момент нарушения поведения. Вместо пианино может быть использован любой музыкальный инструмент, и не только на занятиях по музыке.
Учителя музыки часто исполняют аккорд или несколько аккордов на пианино в момент нарушения поведения. Вместо пианино может быть использован любой музыкальный инструмент, и не только на занятиях по музыке.
Начните говорить тихим голосом. Известны результаты социологического опроса учеников разных классов. На вопрос «Что вам больше всего не нравится в школе?» большинство учеников ответили: «Учителя, которые орут».
Учительские крики не уменьшают уже существующий беспорядок и сильно снижают самоуважение и внутреннюю свободу учеников. Когда мы начинаем говорить тише, ученики, наоборот, прислушиваются и обращают на нас внимание, а это отвлекает их от нарушения поведения. Когда мы говорим спокойно, они тоже говорят спокойно.
Измените манеру речи. Используйте необычную манеру говорить, измените произношение, ударения или начните вдруг говорить на другом языке, может быть, даже на бессмысленном. Шепчите или пойте слова, говорите монотонно, высоким или низким голосом, меняйте тон. Какие-то из этих вокализаций отвлекут учеников от того, чем они занимаются, нарушая правила, и заставят обратить на вас внимание.
Какие-то из этих вокализаций отвлекут учеников от того, чем они занимаются, нарушая правила, и заставят обратить на вас внимание.
Говорите со стеной или «с портретом Пушкина». Этот прием хорошо работает в 5—7 классах. Когда один или несколько ваших учеников плохо ведут себя, повернитесь к стене и начните монолог: «Уважаемая стена, (Александр Сергеевич!) ты не поверишь, что происходит в моем классе сейчас. Одни выкрикивают ответы, не подняв руки, другие сидят ко мне спиной. Не хочешь ли посмотреть на ученика, который пускает бумажные самолетики с задней парты? Вот оно, племя младое, незнакомое!»
Решив попробовать этот прием, заранее предупредите об этом директора, в противном случае он может решить, что вы — кандидат в психиатрическую клинику.
Временно прекратите вести урок. Ученики знают, что учитель в школе, чтобы учить. Когда вы прерываете урок и «ничего не делаете» несколько минут, вы посылаете ученикам впечатляющий сигнал о том, что пора прекратить такое поведение. «Ничего не делать» можно стоя у доски или присев у стола. «Дайте мне знать, когда будете готовы продолжать урок», — вот все, что вам нужно сказать. Ненавязчивое давление старшего вскоре подействует, мир и порядок будут быстро восстановлены.
«Ничего не делать» можно стоя у доски или присев у стола. «Дайте мне знать, когда будете готовы продолжать урок», — вот все, что вам нужно сказать. Ненавязчивое давление старшего вскоре подействует, мир и порядок будут быстро восстановлены.
4 Стратегия. «Отвлеките ученика»
Никто долго не может делать два дела одновременно. А именно это и происходит, когда ученик плохо ведет себя. Поэтому вы можете просто отвлечь его, сфокусировав его внимание на чем-либо другом. Как это сделать практически?
Задавайте прямые вопросы. В критический момент полезно задать ему прямой вопрос: «Роман, какое задание я только что дала?», или: «Миша, что ты думаешь об этой физической проблеме?» Такие вопросы и отвлекают от «плохого» поведения, и направляют внимание ученика на тот урок, на котором он сейчас находится. Попросите об одолжении. «Саша, собери, пожалуйста, сочинения!», «Маша, не могла бы ты отнести эту тетрадь в учительскую прямо сейчас?», «Гриша, сходи к Марье Ивановне в 8 «А» и спроси, не могла бы она дать нам цветной мел?»
Только не используйте этот прием часто, так как демонстративные дети могут решить, что их «плохое» поведение награждается особыми поручениями. Но как единовременная мера он работает очень хорошо.
Но как единовременная мера он работает очень хорошо.
Измените деятельность. Если сразу много учеников «бесятся», чтобы привлечь ваше внимание, резко измените их деятельность, отвлекая от нарушения поведения. Попросите их освободить парты для нового задания, вынуть другие книги, послушать новое задание и так далее.
5 Стратегия. «Обращайте внимание класса на примеры хорошего поведения»
Гораздо полезнее уделить внимание кому-то из учеников, ведущих себя хорошо, чем указывать ученику-нарушителю на его «плохое» поведение. Этим вы подчеркнете, что внимания учителя и его уважения заслуживает хорошее, а не «плохое» поведение.
Благодарите учеников. Благодарите и отмечайте тех учеников, которые делают то, что вы просили сделать: «Спасибо тебе, Саша, что ты нашел нужную страницу в учебнике и внимательно смотришь на доску!», «Спасибо, Оля, что твои руки на парте, а ноги — под партой». Такие утверждения, обращенные к соседу по парте или приятелю ученика-нарушителя, должны точно описывать то самое поведение, которое мы ждем от шалуна.
Этот прием работает, только если мы описываем желаемое поведение в объективных терминах. Общие и неконкретные утверждения типа: «Спасибо, Юля, что ты была такой доброй» или «Спасибо, Саша, что ты делаешь то, что я ожидал», — абсолютно неэффективны, так как неясны эти ваши ожидания. Будьте внимательны, избегайте благодарить одних и тех же учеников слишком часто, дабы не представить их «любимчиками» и не навлечь насмешки класса.
6 Стратегия. «Пересаживайте учеников»
Ученики, которые нуждаются во внимании, как правило, нуждаются и в аудитории. Когда вы отсаживаете таких учеников от их зрителей, вы лишаете их главной награды, и это заставляет учеников успокаиваться. Есть два приема, подходящих для пересаживания.
Просите ученика сесть на другое место. Иногда этого вполне достаточно: «Игорь, пересядь, пожалуйста, на свободное место в третьем ряду», — и продолжайте вести урок, пока Игорь пересаживается. Так он не получит желаемого внимания. А возможно, что такой вид внимания с вашей стороны окажется вполне достаточным и будет воспринят как награда.
Так он не получит желаемого внимания. А возможно, что такой вид внимания с вашей стороны окажется вполне достаточным и будет воспринят как награда.
«Стул размышлений». Некоторые учителя в своем классе ставят специальный «стул размышлений» в отдалении от остального класса (он не должен находиться в поле зрения остальных учеников). Этот стул должен отличаться от остальных. Его можно купить в комиссионке и перекрасить, например, в ярко-красный цвет. Подойдет любой стул, непохожий на остальные.
Этот стул — место, где нарушитель может подумать о том, как он будет вести себя по-другому, вернувшись на свое место. Пять минут на этом стуле — достаточное время. Остальной класс должен понимать, что нельзя беспокоить сидящего на «стуле размышлений».
Но может быть так, что, когда вы говорите ученику-нарушителю: «Женя, пожалуйста, сядь на стул размышлений», он не идет туда. Это не означает, что плох прием, просто цель ученика другая — власть, а не внимание.
Теперь мы предлагаем вам пройти диагностику, которая позволяет судить о психолого-педагогической готовности педагога объективно реагировать на демонстративное поведение ученика.
Методика автора «Педагогические ситуации» состоит из 14 вопросов. Сами вопросы представлены на слайде. На каждый их вопросов необходимо выбрать один из предложенных вариантов ответов.
Теперь оцените свои ответы в соответствии с предложенным ключом.
Спасибо за внимание!
агрессия / Selfire — блог с историями
В природе одни виды неизбежно нападают на другие — называется это межвидовой агрессией. Но не всякое нападение этолог (зоолог, изучающий инстинкты животных) назовет агрессией. Когда волк ловит зайца — это не агрессия, а охота. Точно так же, когда охотник стреляет уток или рыбак ловит рыбу, — это не агрессивное поведение. Ведь все они не испытывают к жертве ни неприязни, ни страха, ни гнева, ни ненависти.
Агрессивное же поведение вызывается этими эмоциями. Вот когда собака выгоняет со двора незнакомого человека — это агрессия. И когда он в ответ бросает в нее камни — тоже агрессия, ибо оба они в этот момент ненавидят и боятся друг друга. Когда дрозд прогоняет от гнезда кота — это агрессия со стороны дрозда. И когда огромный самец лебедя шипением отгоняет от своего гнезда мелких водоплавающих птиц — тоже агрессия. Животное ведет себя по отношению к животному другого вида агрессивно потому, что оно его чем-то раздражает — либо угрожает ему самому (а также его территории, гнезду, детенышам), либо просто незнакомо, подозрительно выглядит.
Объект агрессии может быть крупнее субъекта, а может и меньше. У нас вспышку агрессивности может вызвать и загородившая дорогу корова, и вьющаяся вокруг головы муха. Польза межвидовой агрессии вроде бы ясна: дрозду выгоднее атаковать кошку, чем позволить ей съесть птенцов. Ясно и то, что такая агрессия сопровождается страхом: кошка далеко не беззащитна, и атака может кончиться для дрозда плачевно. Нападая на вьющееся вокруг головы насекомое, мы тоже побаиваемся — ведь среди них есть жалящие.
Нападая на вьющееся вокруг головы насекомое, мы тоже побаиваемся — ведь среди них есть жалящие.
Внутривидовая агрессия. Казалось бы, без нее природа могла обойтись. Но это не так. Особи одного вида неизбежно вступают в конфликт. Можно не поделить пищу или удобное для отдыха место. Живущие каждый на своей территории виды должны изгонять конкурентов. Неизбежны конфликты из-за самки, дупла, норы и многих других причин.
Появление или приближение другой особи с неясными намерениями неизбежно вызывает настороженность (а это легкая форма страха). Если намерения не проясняются, зачастую ничего другого не остается, как либо убежать, либо напасть первым. То же происходит и с приближающимся животным. Вступая в конфликт, оба животных испытывают страх. И вместе с ним — приступ агрессивности.
Агрессия и страх — близнецы. Все мы с яростью и бесстрашием хлопаем укусившего нас комара, бесстрашно можем оттолкнуть от себя или прогнать прочь и животное покрупнее, но заведомо безобидное. Однако при встрече с животным незнакомым или способным как-то за себя постоять, не говоря уже о действительно опасном, наша агрессия сопровождается заметным для нас самих приступом страха. Вспомните, как нервозно люди отгоняют паука или ловят забежавшую в комнату мышь. Изгнание пчелы или осы сопровождается уже явным приступом страха. Конфликт с маленькой собачкой может довести до сердцебиения.
Однако при встрече с животным незнакомым или способным как-то за себя постоять, не говоря уже о действительно опасном, наша агрессия сопровождается заметным для нас самих приступом страха. Вспомните, как нервозно люди отгоняют паука или ловят забежавшую в комнату мышь. Изгнание пчелы или осы сопровождается уже явным приступом страха. Конфликт с маленькой собачкой может довести до сердцебиения.
Читатель, видимо, согласится с выводом этологов, что агрессия и страх взаимосвязаны. Агрессия всегда сопровождается приступом страха, а страх может перерастать в агрессию. Самые разнообразные опыты на животных показали, что это так. Если на группу животных нагонят страх, они становятся агрессивнее. То же происходит и с толпой людей или обществом в целом. Агрессивно-трусливое состояние — самое опасное.
В. Р. Дольник, доктор биологических наук, профессор, главный научный сотрудник Зоологического института РАН. Вице-президент Российского орнитологического общества, почетный член орнитологических обществ США, Германии, Нидерландов. Научные интересы связаны с экологией и поведением животных.
Научные интересы связаны с экологией и поведением животных.
Демонстративное поведение педагога как инструмент воздействия на окружающих
О. А. Петрова, г. Соликамск
Демонстративное поведение – один из многочисленных инструментов воздействия на окружающих с целью удовлетворения тех или иных потребностей, поскольку в социальных условиях жизни удовлетворение многих потребностей человека зависит от отношения к нему других людей, на изменение которого, осознанно или нет, направлены его демонстрации.
На современном этапе развития общества нельзя не согласиться, что некоторые социальные ситуации навязывают необходимость демонстративного поведения. Экономические и политические события в России последних лет повлекли за собой социально-психологические изменения, которые выражаются в частности, в изменении социально-одобряемого поведения. Поворот в сторону капиталистического пути развития общества вызвал у большей части населения потребность в поиске работы. Одновременно с этим появилось большое количество новых профессий. В этих условиях огромное значение для многих людей приобрело умение демонстрировать себя, то есть умение произвести благоприятное впечатление о себе.
Одновременно с этим появилось большое количество новых профессий. В этих условиях огромное значение для многих людей приобрело умение демонстрировать себя, то есть умение произвести благоприятное впечатление о себе.
Умение и желание привлекать внимание к себе – непременное условие
многих публичных профессий, в том числе и профессии педагога. Вступая
во взаимоотношения с учащимися, педагог предлагает себя как партнёр по
общению. Это предполагает определённую активность учителя. Желательно, чтобы он создал позитивное впечатление о себе в глазах учеников. Эта
способность «вмешательства» живого объекта восприятия в процессе формирования своего образа у собеседника некоторыми авторами называется
демонстративностью [5]. А. А. Леонтьев выделяет три основных мотива демонстративности: стремление к развитию отношений, самоутверждение
личности, необходимость профессионального плана [7,10].
Таким образом, демонстративное поведение может быть использовано
для реализации фактически любых социальных потребностей. Анализируя различные классификации мотивов демонстративности, таких авторов как: Р. Аркин, Э. Гофман, У. Джеймс, Е. Е.Джонсс, Р. М.Ковальски, М. Р.Лири, Т. С.Питтман [1,2,6,9], появляется возможность систематизировать представления об инструментальном использовании демонстративного поведения:
Анализируя различные классификации мотивов демонстративности, таких авторов как: Р. Аркин, Э. Гофман, У. Джеймс, Е. Е.Джонсс, Р. М.Ковальски, М. Р.Лири, Т. С.Питтман [1,2,6,9], появляется возможность систематизировать представления об инструментальном использовании демонстративного поведения:
1. Инрациация – наиболее распространённый вид самопрезентации, цель которой – выглядеть привлекательным, вызывать расположение, втереться в доверие. Успешность этого вида самопрезентации зависит от видимой искренности субъекта. Возможные негативные результаты этого вида самопрезентации — оценка инграциатора как льстеца, подхалима, конформиста и т. д. В своей работе Е. Е.Джонсс (1964) различает 3 стратегий инграциации: представление себя в выгодном свете, самовосхваление; лесть и похвалы адресату самопрезентации. Инграциация может быть следствием поиска одобрения и попытки избежать неодобрения, принятия со стороны других и т. д. [9].
2. Запугивание – демонстрация силы. Этот вид самопрезентации чаще всего встречается в отношениях сверху – вниз (начальник – подчиненный, родитель – ребенок). Эта стратегия может быть использована и в противоположную сторону.
Эта стратегия может быть использована и в противоположную сторону.
3. Самореклама – человек старается выглядеть компетентным в той или иной области, что даёт ему «власть эксперта». При разоблачении или отсутствии убедительных доказательств компетентности он – мошенник, тщеславный, хвастун и пр. В мотивации этого вида самопрезентации принимают участие потребности социального признания, признание компетентности, нужности.
4. Примерность – требует от демонстранта постоянных усилий. Всё поведение и сама жизнь такого человека демонстрирует достойность, чистоту помыслов, порядочность, что даёт ему право вести за собой, распоряжаться судьбами других. Подобное демонстративное поведение может определяться потребностями самоуважения, самопринятия, самоопределения и пр.
5. Самоуничижение – создаёт образ несчастного, обездоленного, беспомощного человека которому необходима помощь. Такой человек «замечателен» своими изъянами. Наряду с чисто манипулятивными использованиями самоуничижения в основе этого поведения в равной степени могут быть потребности самооправдания, призыв о помощи и пр.
В педагогическом же общении демонстративность является одним из профессионально значимых качеств учителя, так как она помогает самовыражению как учителя, так и ученика. Успех во всевозможных конкурсах педагогического мастерства, при приёме на работу, а чаще всего в повседневных ситуациях совместной деятельности с родителями, учениками, коллегами, ситуациях знакомства, различных общественных мероприятиях также предполагает умение привлечь внимание к себе и создать о себе определённое впечатление, чаще всего благоприятное.
Способность «работать на публику» или демонстративность является
основным профессионально важным качество талантливого педагога любого «ранга»: от воспитателя детского сада до профессора университета.
Такой человек по своей природе нуждается в том, чтобы на него смотрели, его слушали – но при этом он сам обладает достаточным умением привлекать чужое внимание, яркой образной речью, и артистичностью в принципе. Такой педагог выложится, что называется, до конца – только бы заинтересовать учеников, чтобы они увидели своими глазами то, что он рассказывает, и увлеклись этим.
Люди, умеющие красиво и образно рассказывать, были учителями ещё в первобытные времена, когда не было ни досок, ни тетрадей, ни карт и атласов – и только демонстративная личность, имеющая, прежде всего, актёрские задатки и образную речь могла достаточно наглядно передать ученикам необходимые знания и навыки. И сегодня, именно демонстративный учитель способен ярко и эмоционально излагать предмет, быть искренне увлечённым и увлекать слушателей, являться самым результативным педагогом. Демонстративный учитель способен к любым перевоплощениям. Если перед учениками он может быть и строгим наставником, то вне класса он вполне способен перевоплотиться в ровесника своих подопечных. Именно в силу его актёрских, демонстративных способностей у него прекрасно срабатывает один из основных принципов успешного обучения «Учиться играя». Учитель с такими качествами часто тяготеет к различного рода экспериментам, которые всегда имеют одну основную цель – привлечение внимания к предмету, повышение интереса учеников [8]. Но когда артистичность начинает определять стиль поведения, и основной целью педагога становится привлечение внимания не к предмету, а к своей персоне, то это снижает качество педагогической деятельности и становится средством самоуважения и самопрезентации педагога.
Но когда артистичность начинает определять стиль поведения, и основной целью педагога становится привлечение внимания не к предмету, а к своей персоне, то это снижает качество педагогической деятельности и становится средством самоуважения и самопрезентации педагога.
Такое проявление демонстративности является одной из основных профессиональных деформаций личности педагога. Данная тенденция реализуется в оригинальном поведении, демонстрации своего превосходства, преувеличениях, расцвечивании своих переживаний, в позах, поступках, рассчитанных на внешний эффект.
Демонстративность как профессионально нежелательное качество заключается в чрезмерной эмоциональности и самопрезентации, она проявляется на профессионально-типологическом уровне профессиональной деформации (на уровне способностей). Демонстративность как деформация, по мнению Э. Ф. Зеера, — это разновидность психологической защиты — индифферентности, которая проявляется в завышенной самооценке
«Я-образа» и эгоцентризме [5].
В профессиональной деятельности демонстративность проявляется в стремление добиваться для себя льгот и послаблений за счет коллег, семьи
и т. д. Попытки реализации идут по двум направлениям: во-первых, предпринимаются шаги с целью вызвать по отношению к себе как можно больше симпатии, уважения, восхищения и т. д.; во-вторых, если не срабатывает первый способ, со стороны окружающих стимулируются чувства сострадания и сочувствия. Трудности демонстративной личности проявляются в том, что человек не в состоянии оказаться за пределами эгоцентрического уровня личностно-смысловой сферы и низкого уровня самосознания. Если он еще в какой-то мере способен осознавать себя в когнитивном аспекте профессионального самосознания, то его Я-отношение обнаруживает явную деформацию в силу резко завышенной самооценки.
Педагоги с «негативной» демонстративностью, как правило, начинают чрезвычайно ярко выражать свои агрессивные эмоции – кричат, громко ругаются, разбрасывают вещи, их поведение становится направленным на получение эмоционального отклика от окружающих. Они активно стремятся к контактам с окружающими, а добившись внимания от партнёра по общению успокаиваются и прекращают свои вызывающие действия. Педагог с данной тенденцией также часто тяготеет к различного рода экспериментам, которые не всегда вызывают удовольствие у окружающих, но всегда имеют основную цель – привлечение внимания. Им свойственна мания собственного «Я», эгоцентричность [5].
Они активно стремятся к контактам с окружающими, а добившись внимания от партнёра по общению успокаиваются и прекращают свои вызывающие действия. Педагог с данной тенденцией также часто тяготеет к различного рода экспериментам, которые не всегда вызывают удовольствие у окружающих, но всегда имеют основную цель – привлечение внимания. Им свойственна мания собственного «Я», эгоцентричность [5].
Отношения с коллегами у демонстративной личности часто нарушаются, что связанно с неспособностью понимать других, во главу ставятся только собственные интересы, а интересы других не учитываются, другие люди воспринимаются как часть самого себя. Для такой личности характерно стремление манипулировать людьми, относиться к ним свысока, пренебрегать другим, что часто производит отталкивающее впечатление. Демонстративный педагог, показывая своё превосходство над учащимися, часто злоупотребляет запретами и отказами. Попадая в конфликтную ситуацию между спонтанным интересом и педагогическим «нельзя», ребёнок невольно испытывает сильнейшую депривацию – ограничение возможности удовлетворения своих потребностей. Другими словами, приводит к состоянию фрустрации. Ребёнок воспринимает эту ситуацию как акт отвержения со стороны педагога. Невозможность решения данной ситуации приводит к тому, что в нём просыпается злость, отчаяние, агрессивность. В целом личность с демонстративным поведением относится к окружающему миру с подозрением, недоверием. У них легко возникает мысль о том, что все их ненавидят, стараются помешать их успеху, стремятся обмануть. Такие оценки возникают тогда, когда они встречаются с объективными трудностями, критикой, замечаниями, даже если последние носят доброжелательный характера.
Другими словами, приводит к состоянию фрустрации. Ребёнок воспринимает эту ситуацию как акт отвержения со стороны педагога. Невозможность решения данной ситуации приводит к тому, что в нём просыпается злость, отчаяние, агрессивность. В целом личность с демонстративным поведением относится к окружающему миру с подозрением, недоверием. У них легко возникает мысль о том, что все их ненавидят, стараются помешать их успеху, стремятся обмануть. Такие оценки возникают тогда, когда они встречаются с объективными трудностями, критикой, замечаниями, даже если последние носят доброжелательный характера.
Существует два вида демонстративного самопредъявления: негативное и позитивное. У педагога с негативным самопредъявлением как правило складывается самосознание «нигилиста», чья заметность в обществе достигается экстравагантностью и демонстративным противопоставлением себя окружающим. Синдром позитивного самопредъявления близок к синдрому негативного самопредъявления с тем отличием, что в этом случае внимание привлекается не по средством нарушения норм и правил, а напротив,
посредством их подчёркнутого соблюдения. Педагог характеризуется самосознанием «образцового члена общества». Разумеется, формы внимания, получаемого при позитивном самопредъявлении, более привлекательны, чем при негативном, оно проявляется в восхищении и умилении окружающими. Демонстративность часто перерастает в личностную особенность, черту характера, которая приносит массу негативных переживаний человеку. Основным мотивом действия педагога с позитивным самопредъявлением, становится положительная оценка окружающих, с помощью которой он удовлетворяет собственную потребность в самоутверждении. Совершая добрый поступок, личность делает это не ради другого, а ради того, чтобы продемонстрировать окружающим собственную доброту [3;205].
Педагог характеризуется самосознанием «образцового члена общества». Разумеется, формы внимания, получаемого при позитивном самопредъявлении, более привлекательны, чем при негативном, оно проявляется в восхищении и умилении окружающими. Демонстративность часто перерастает в личностную особенность, черту характера, которая приносит массу негативных переживаний человеку. Основным мотивом действия педагога с позитивным самопредъявлением, становится положительная оценка окружающих, с помощью которой он удовлетворяет собственную потребность в самоутверждении. Совершая добрый поступок, личность делает это не ради другого, а ради того, чтобы продемонстрировать окружающим собственную доброту [3;205].
Представители различных теоретических направлений отмечают многообразие причин, обуславливающих формирование педагогической демонстративности как профессионально обусловленной деформации. A. M. Павлова, А. П. Зольнико, Э. Ф. Зеер выделяют следующие психологические факторы, способствующие появлению демонстративного поведения у педагога:
1. Сверхнормативная активность как следствие неудовлетворенности своим положением, своим статусом – в этом случае педагог начинает повышать свое положение через преувеличение своих способностей демонстрацию себя как самого «умного» и «незаменимого» работника.
Сверхнормативная активность как следствие неудовлетворенности своим положением, своим статусом – в этом случае педагог начинает повышать свое положение через преувеличение своих способностей демонстрацию себя как самого «умного» и «незаменимого» работника.
2. Вступление в новую должность (а также – участие в конкурсах на замещение, в аттестациях и т. д.) – педагог стремится к победе, похвале и тем самым старается привлечь к себе внимание по средствам эмоционально окрашенного поведения, демонстрацию своего превосходства.
3. Акцентуации характера (профессиональные акцентуации – это чрезмерное усиление некоторых черт характера, а также отдельных профессионально обусловленных свойств и качеств личности) – педагогам присуща демонстративность в поведении, но когда демонстративность выходит за рамки разумного можно говорить об акцентуации данного поведения.
4. Образование стереотипов профессионального поведения [4].
Таким образом, демонстративное поведение может быть инструментом воздействия на окружающих людей в процессе взаимодействия для решения каких либо социальных задач. Демонстративность не считается отрицательным и, собственно, проблемным качеством. Более того, в настоящее время некоторые особенности, присущие демонстративным людям, напротив, являются социально одобряемыми: настойчивость, здоровый эгоизм, способность добиться своего, стремление к признанию, честолюбие считают залогом успешной жизненной позиции. Однако при этом не учитывается, что противопоставление себя другому, болезненная потребность в признании и самоутверждении являются зыбким фундаментом психологического комфорта и мотивации тех или иных поступков. Не насыщаемая потребность в похвале, в превосходстве над другими становится главным мотивом всех действий и поступков. Такой педагог постоянно боится оказаться хуже других, что порождает тревожность, неуверенность в себе, которая компенсируется хвастовством и
Демонстративность не считается отрицательным и, собственно, проблемным качеством. Более того, в настоящее время некоторые особенности, присущие демонстративным людям, напротив, являются социально одобряемыми: настойчивость, здоровый эгоизм, способность добиться своего, стремление к признанию, честолюбие считают залогом успешной жизненной позиции. Однако при этом не учитывается, что противопоставление себя другому, болезненная потребность в признании и самоутверждении являются зыбким фундаментом психологического комфорта и мотивации тех или иных поступков. Не насыщаемая потребность в похвале, в превосходстве над другими становится главным мотивом всех действий и поступков. Такой педагог постоянно боится оказаться хуже других, что порождает тревожность, неуверенность в себе, которая компенсируется хвастовством и
подчёркиванием своих преимуществ и как следствие приводит к повышению тревожности и агрессивности учащихся. Поэтому важно вовремя выявить проявления демонстративности, как личного качества и помочь педагогу в преодолении такой конкурентной позиции.
(Материал взят из книги Антропологические основы современной теории и практики образования: Материалы V Международной научно-практической конференции)
Трудное поведение ребенка
Трудное поведение ребенка
Часто приемные родители сталкиваются с проблемами в поведении приемного ребенка. Многие из нас не знают, как вести себя в таких ситуациях, как правильно реагировать. Сегодня в статье попробуем разобраться учиться управлять проблемным поведением приемных детей.
Какое поведение ребенка — трудное?
Родителям трудно принять поведение ребенка, если оно социально неприемлемо. Также иногда поведение ребенка может не особенно беспокоить окружающих, но при этом нести непосредственную опасность для его жизни и здоровья. Наконец, «трудным» может стать поведение, которое, не являясь чем-то негативным с общепринятой точки зрения, разрушает семейную атмосферу и выводит из себя родителей.
Если поведение ребенка «странно» для родителей и при этом они не могут найти разумных объяснений его действиям, то оно может вызывать сильное беспокойство, казаться «ненормальным».
Таким образом, по-настоящему «трудным» оказывается то поведение ребенка, которое:
· неприемлемо для конкретной семьи,
· неприемлемо с социальной точки зрения,
· непонятны причины этого поведения,
· поведение опасно или вредно для самого ребенка.
Важно отметить, что под словами «трудное поведение» здесь не подразумевается криминальное поведение или поступки, совершенные вследствие диагностированных психических нарушений.
Нередко трудное поведение возникает не из глобальных расхождений во взглядах на жизнь, а накапливается как снежный ком из мелких недопониманий и в конце концов приводит к бурным конфликтам. Кроме причин, которые действительно важно понять, есть тема «немедленного реагирования» на конкретные поступки ребенка. Родители могут растеряться как раз в тот момент, когда им нужно действовать быстро и желательно разумно. Вот идеи, которые помогут родителям разработать для себя подходящие способы «быстрого реагирования» в сложных ситуациях.
Родители могут растеряться как раз в тот момент, когда им нужно действовать быстро и желательно разумно. Вот идеи, которые помогут родителям разработать для себя подходящие способы «быстрого реагирования» в сложных ситуациях.
Он просто подражает
Попадая в любую новую ситуацию, человек многим вещам учится, наблюдая за окружающими и повторяя их действия. Очень часто негативное поведение детей является не просто спонтанным выражением чувств, а воспроизведением примеров плохого поведения, которые ребенок наблюдал в своей жизни. Если в кровной семье или детском доме взрослые позволяли себе нецензурно ругаться, бить детей за нарушение правил, то ребенок может потом в приемной семье вести себя подобным же образом.
К счастью, усвоение моделей поведения — вещь универсальная. Усваиваются не только плохие, но и хорошие модели. Когда действия ребенка в определенной ситуации неприемлемы, можно предложить ему другие варианты действий на конкретном примере, и поощрить ребенка, чтобы он пытался действовать такими же приемлемыми способами. Для этого подходят как реальные примеры из жизни, так и сказки, фильмы, метафоры и т.д.
Для этого подходят как реальные примеры из жизни, так и сказки, фильмы, метафоры и т.д.
«Побей меня, я буду хорошо себя вести»
В некоторых случаях для взрослых трудным становится поведение, которым дети провоцируют их на плохое обращение. Так бывает, когда в жизни ребенка была только одна модель отношений со взрослыми: нежелательное поведение пресекалось агрессией и побоями. Соответственно, для ребенка это привычное и понятное ему взаимодействие, которого он будет ожидать.
Если взрослые ведут себя иначе, ребенок перестает понимать, что происходит. Тогда он может начать добиваться своими «безобразиями» привычных тумаков, некоторые даже формулируют словами: «А ты побей меня, я буду хорошо себя вести». Взрослые могут поддаться на эти провокации, исходя из логики, что раз действует, то надо использовать этот метод воспитания. Тогда для ребенка подтверждается его опыт, и агрессивные взаимодействия остаются для него предпочтительным способом общения с другими людьми.
Правильное решение — прерывание цепочки негативного взаимодействия, поиск и формирование других, альтернативных способов поведения и для родителей, и для детей. Принципиальная вещь — отказ родителей от битья и агрессии, на которые их провоцирует ребенок. Сдерживать и выстраивать границы, отстаивать свой авторитет можно и без физической агрессии.
Тайм-аут
Многие взрослые используют тайм-аут как способ самоконтроля: стремятся уединиться, когда нужно выйти из напряженной ситуации и спокойно обдумать создавшееся положение. Кому-то из людей необходимо уединиться и побыть в тишине, кому-то — наоборот, пройтись, пробежаться.
Вне зависимости от специфики ситуации общая идея — приостановка нарастающего напряжения, которое грозит завершиться взрывом. Маленькие дети не могут контролировать себя сами, когда их переполняют эмоции. Им в этом нужна помощь взрослых, которые не только влияют на поведение ребенка, но и своими действиями предоставляют ребенку образцы того, как он в дальнейшем может научиться управлять собою сам. Ребенку говорят: «Побудь в своей комнате, успокойся, а потом мы поговорим»; «Я не буду разговаривать с тобой в таком тоне, я выслушаю тебя, когда ты будешь не кричать и браниться, а сможешь сказать, что ты хочешь, и мы это обсудим»; «Ты идешь „вразнос“, посиди на табуретке до тех пор, пока ты не придешь в себя».
Хвалить или ругать ребенка?
Позитивное подкрепление побуждает детей продолжать и повторять совершенные ими действия. Формы позитивного подкрепления могут быть самые разные: похвала, признание успехов и способностей ребенка, благодарность, материальные поощрения (призы, символические награды и т.д.). Безусловно, имеет значение, от кого исходит одобрение: чем более значимый человек благосклонно реагирует на действия ребенка, тем более вдохновляющим это становится для него.
Часто родителям кажется, что то, что ребенок делает правильно, — это естественно, поэтому они сами не особенно обращают на это внимание, да и ребенка стараются лишний раз не хвалить, чтобы «не загордился». А вот ругать и стыдить за ошибки считается очень действенным. Но на самом деле любой человек с большим удовольствием делает то, с чем у него связаны приятные чувства. И похвала естественным образом закрепляет правильное поведение ребенка.
Привлекать внимание — приемлемым способом
В некоторых случаях негативное поведение ребенка является демонстративной попыткой привлечь к себе внимание. Если взрослый человек ясно понимает, что безобразие, творимое ребенком, имеет целью вызвать бурную эмоциональную реакцию у взрослых, то имеет смысл игнорировать подобное поведение, не предоставляя ребенку желаемой реакции. Стоит пояснить ребенку происходящее: «Когда ты делаешь то-то и то-то, я не хочу на тебя смотреть и разговаривать с тобой. Мне это неинтересно».
Если ожидаемой ребенком реакции взрослого не следует, ребенок поначалу может усиливать демонстративное поведение. Но если взрослый продолжает игнорирование и старается переключить активность ребенка на что-то другое (например, предлагает заняться чем-то еще), то информация, которую получает ребенок, — «такая форма поведения не действует на взрослого». При этом полезно предлагать ребенку альтернативы: «Я буду обращать на тебя внимание / мне будет интересно с тобой / я с удовольствием посмотрю, как ты делаешь то-то и то-то». Цель состоит не в том, чтобы доставлять друг другу неприятности, а в том, чтобы научить ребенка получать внимание приемлемым способом.
Что такое тактика малых шагов?
Зачастую бывает так, что уставшие от плохого поведения ребенка и других жизненных трудностей родители начинают обобщать проступки ребенка, выстраивая для себя и для него единую картину «глобальной испорченности». Взрослые объединяют проступки ребенка разного срока давности, разной степени тяжести, вызванные разными причинами. В эти моменты родителями движут отчаяние, страхи, ощущение собственной некомпетентности или бессилия. Им кажется, что они сделали все, что могли, а ребенок продолжает вести себя неправильно.
Сама по себе тенденция к обобщению связана с попыткой систематизировать опыт и выделить в нем главное, существенное. Но в отношениях, особенно в проблемных, эта тенденция вредна и ошибочна.
Во-первых, в кризисных ситуациях основой для изменений служит принципиальная надежда на то, что изменения к лучшему возможны. Негативные обобщения эту надежду убивают. Во-вторых, объединение явлений разного порядка и значения по сути является не обобщением, а «сваливанием в одну кучу», и имеет под собой только эмоциональное основание, далекое от справедливости.
Важно помнить: негативные обобщения являются способом сбросить напряжение, «поругаться». Но никакого положительного влияния на процесс воспитания ребенка это не оказывает.
Сохранить отношения помогает то, что называется «тактикой малых шагов»:
· Для начала стоит выделить какую-то одну форму негативного поведения ребенка и начать целенаправленно работать именно с ней.
· Далее важно ставить конкретные и реалистичные цели. Например, не просто «я хочу, чтобы мой ребенок хорошо учился», а я хочу, чтобы вместо «2» по биологии к концу года была «3» (это не обязательно является конечной целью, дальше может быть «4» или «5»).
· Постановка целей должна происходить совместно с ребенком: «Ты знаешь, у нас очень много проблем сейчас в жизни. И чтобы улучшить нашу жизнь, давай начинать с чего-то конкретного. Я очень волнуюсь за твою учебу, и мне хотелось бы начать изменения к лучшему с этой области. А как ты думаешь?». Если ребенок не хочет начинать с учебных проблем, то стоит вместе поискать что-то другое. В этом случае будет удовлетворена потребность ребенка в самостоятельности и в том, что его мнение уважается. В то же время работа над любой целью, которая будет поставлена совместно, будет учить ребенка последовательным усилиям, умению ставить цели и достигать их, сотрудничеству со взрослым и т.д.
· Важно не сравнивать ребенка с другими, а сравнивать с ним самим на предыдущих жизненных этапах. Например: «Смотри, два года назад ты не умел читать-писать, а сейчас ты умеешь это делать»; «Ты боишься, что у тебя не получится, и не хочешь это делать. С тобой так бывало и раньше, но потом ты справился… (конкретное перечисление достижений ребенка)». Важно уметь видеть и помнить прошлые успехи ребенка, потому что именно они являются аргументом в пользу возможности изменений к лучшему.
· Собственно тактика малых шагов заключается в том, чтобы взрослые обращали внимание на любой, даже самый малый прогресс в делах ребенка, хвалили за предпринятые усилия, даже небольшие, обращали его внимание на положительные изменения. В этом процессе важна последовательность и терпение взрослых, которым нужно убедить ребенка в том, что он способен добиваться хороших результатов и что это на самом деле происходит, а также что это важно для его близких людей, которые в него верят. Например: «Я вижу, что на этой неделе ты ни разу не прогулял биологию, выполнял домашние задания. У тебя нет отметок, но отсутствие двоек — это тоже положительный результат. А хорошие отметки появятся позже, я уверена»; «Сегодня ты делал аппликацию, ты вырезал все детальки сам, и тебе удалось гораздо ровнее разместить их на листе, и только половина стола запачкалась клеем. Все это тебе удалось лучше, чем две недели назад, помнишь? Ты молодец».
Очевидно, что когда люди хронически расстроены и раздражены, им трудно видеть позитив и серьезным образом следовать этой тактике. Поэтому родителям необходимо иметь возможность получать индивидуальную поддержку и помощь у друзей, родственников и специалистов.
Опубликована монография Л.А. Зеленской «Тихоокеанская чайка»
Опубликована монография Л.А. Зеленской «Тихоокеанская чайка»
- Подробности
- Создано 15.10.2008 15:36
- Обновлено 18.11.2013 06:27
Зеленская Л. А. Тихоокеанская чайка (Larus schistisagus Stejneger, 1884). Магадан: СВНЦ ДВО РАН, 2008. 213 с.
Тихоокеанская чайка — самая многочисленная из крупных чаек российских дальневосточных морей. Данные по размножению этого вида, питанию, поведению, взаимоотношению с хищниками и растительностью на гнездовьях приведены в основном по результатам полевых работ автора в 1986-2006 гг. В книге обобщены сведения по таксономии, гибридизации, возрастным нарядам, распространению, миграциям и морфометрии и дополнены материалами автора. Приводятся оригинальные данные по биотопическим предпочтениям, формированию и росту колоний, гнездовой биологии, росту, развитию и «усыновлению» птенцов. По результатам суточных наблюдений рассчитан бюджет времени и энергии в период инкубации. Описываются состав кормов и особенности поведения чаек при поиске и добывании пищи как в естественных экосистемах, так и на свалке пищевых отходов. Показаны особенности экологии чаек, гнездящихся на крышах зданий в городе. Изложены данные по суточной и сезонной активности чаек, проанализировано их демонстративное поведение в репродуктивный период. Прослеживаются взаимосвязи с растительностью на гнездовых колониях и белоплечим орланом, для которого тихоокеанская чайка часто является основной добычей в ключевые моменты его гнездования.
Для биологов, преподавателей зоологии и экологии, любителей природы.
Ил. 93. Табл. 55. Библиогр.: 497 назв.
Ключевые слова: тихоокеанская чайка, численность, распространение, биология размножения, экология питания, кормодобывание, демонстративное поведение, биотопическое распределение, рост птенцов, «усыновление» птенцов.
Утверждено к печати ученым советом ИБПС ДВО РАН.
Ответственный редактор д. б. н. А. В. Андреев.
Рецензенты: к. б. н. В. А. Зубакин, к. б. н. Г. И. Атрашкевич
Zelenskaya L. A. Slaty-backed Gull (Larus schistisagus Stejneger, 1884). Magadan: NESC FEB RAS,2008.213p.
Slaty-backed Gulls (Larus schistisagus) are the most numerous of all the large gulls of the Far-Eastern seas of Russia. Data on breeding, feeding, behavior, interactions with predators and association with vegetation at nesting sites, as well as characteristics of urbanized population ecology, are presented mainly according to the results of the author’s fieldwork from 1986-2006. Information on taxonomy, hybridization, age-related plumage, distribution, migration and morphometrics is summarized in the text. Comparative analysis is made with corresponding literature. Data are also provided on biotopical preferences, formation and expansion of colonies, nesting biology, and growth and development of chicks. Food composition and characteristics of foraging behavior are described both for free-ranging and roof-nesting urban populations. Data on daily and seasonal activity of the gulls are presented; display behavior during the reproductive period is analyzed. Interrelationships with vegetation at the nesting colonies and with the Steller’s Sea Eagle (Haliaeetuspelagicus), for which the Slaty-backed Gull is the primary prey during key phases of the eagles’ nesting period are documented.
For biologists, teachers of zoology and ecology and naturalists.
Ills. 93. Tables 55. Refs.: 497 titles.
Key words: Slaty-backed Gull, numbers, distribution, breeding biology, feeding ecology, foraging behavior, displays, habitat distribution, chick growth, adoption.
Edited by Dr. A. V. Andreev
Reviewer: Dr. V. A. Zubakin, Dr. G. I. Atrashkevich
Приспособительные особенности строения окраски и поведения. Приспособительные особенности строения окраски тела и поведения животных. Совместная постановка целей урока
Вопрос 1. Приведите примеры приспособленности организмов к условиям существования на основании собственных наблюдений.
В ходе эволюции организмы приобретают различные свойства, позволяющие им более успешно приспосабливаться к условиям обитания. Например, мех животных севера (песцов, медведей) имеет белую окраску, делая их практически незаметными на фоне снега. Насекомые, питающиеся цветочным нектаром, имеют строение и длину хоботка, идеально подходящие для этого. Ласты тюленей, преобразованные из лап их сухопутных предков, прекрасно приспособлены к передвижению в воде. Жирафы живут в саванне и поедают листья деревьев на большой высоте, в чем им помогает длинная шея.
Таких примеров можно привести множество, так как каждое живое существо обладает большим количеством признаков, приобретенных в процессе приспособления к конкретным условиям обитания.
Вопрос 2. Почему у некоторых животных яркая, демаскирующая окраска, а у других, наоборот, покровительственная?
Двум типам окраски соответствуют два варианта стратегии поведения. При одном из них животное стремится остаться незамеченным, пытаясь избежать встречи с хищником или подкрадываясь к жертве. Для этого используется покровительственная окраска, позволяющая сливаться с фоном. С другой стороны, животные, которые являются опасными или ядовитыми, часто всячески это подчеркивают. Они используют яркую демаскирующую окраску, предостерегающую: «не ешь меня». Кроме ядовитых организмов, эту стратегию применяют мимикрирующие под них безобидные виды. Организмы могут иметь демаскирующую окраску и по совершенно иной причине — в связи со стремлением привлечь партнера для размножения (яркая окраска многих самцов птиц, рыб, рептилий, бабочек и т. д.). В этом случае задача продолжения рода приходит в противоречие с инстинктом самосохранения, но оказывается для организма более значимой.
Вопрос 3. В чём сущность мимикрии? Сравните мимикрию и маскировку. В чём их принципиальные отличия? Чем они схожи?
Сущность мимикрии (от греч. mimikos — подражательный) заключается в том, что безобидные животные в процессе эволюции приобретают сходство с опасными (ядовитыми) видами. Это позволяет им избегать нападения хищников. Примером служат некоторые неядовитые змеи: существует вид полоза, который расцветкой похож на смертельно опасного аспида и отличается от него лишь чередованием полос. Кроме окраски мимикрирующие животные обладают характерным поведением: мухи-журчалки ведут себя подобно осам, имитируя агрессию.
Вопрос 4. Распространяется ли действие естественного отбора на поведение животных? Приведите примеры.
Естественный отбор влияет не только на внешние признаки организма, но и на поведение. Это относится, прежде всего, к врожденным (инстинктивным) формам поведения. Такие формы очень разнообразны: способы добывания пищи, проявления страха и агрессии, половое поведение, родительское поведение и т. д. Паук плетет сеть, пчела строит соты, кошка в момент опасности принимает угрожающую позу, бурундуки делают запасы и впадают в спячку на зиму и т. д. Очень сложны брачные ритуалы, строгое следование которым является для животных одним из путей предотвращения межвидового скрещивания.
Вопрос 5. Каковы биологические механизмы возникновения приспособительной (скрывающей и предупреждающей) окраски у животных?
Биологическим механизмом, обеспечивающим возникновение приспособительной окраски, является естественный отбор. В процессе эволюции в популяции, которая в силу разнообразия генофонда отличалась очень широким спектром окраски, преимущественно выживали и оставляли потомство те особи, которые были менее заметны на фоне окружающей среды. В результате доля соответствующих генотипов постоянно росла. В дальнейшем данный фенотип, а следовательно, и генотип закреплялся в популяции с помощью стабилизирующего отбора. В случае предостерегающей окраски происходили аналогичные процессы. Например, птицы исходно легче находят и поедают ярких насекомых. Если эти насекомые оказываются ядовитыми, то птицы быстро обучаются их не трогать и предпочитают более скромно окрашенную добычу. Тем самым особи с яркой окраской, которую легко идентифицировать как ядовитую, сохраняются и оставляют потомство. Со временем данный признак закрепляется в популяции.
Вопрос 6. Существуют ли живые организмы, не имеющие приспособительных особенностей строения? Ответ обоснуйте.
Адаптация –совокупность особенностей строения, физиологии и поведения живых организмов к конкретным условиям, в которых они могут нормально существовать и оставлять потомство.
Возникновение приспособленности к среде — основной результат эволюции. Поэтому эволюцию можно рассматривать как процесс возникновения приспособлений или адаптаций.
Организмы не сумевшие приспособиться к окружающему миру вымерли.
Приспособительные особенности строения, окраски тела и поведения животных (сопроводительный материал к уроку общей биологии по учебнику Н. И. Сонина В. Б. Захарова. 9 класс).
Приспособленность. Форма тела. ВИДЫ ПРИСПОСОБЛЕННОСТИ ЗНАЧЕНИЕ ПРИМЕРЫ Форма тела: — торпедообразная — сучковидная, листовидная — причудливая Способствует избежанию образования завихрений Акулы, дельфины потоков воды при движении Делает организм незаметным среди тех или иных предметов Палочники, гусеницы пядениц среды Скрывает среди водорослей, коралловых полипов Морские коньки, удильщики
Форма тела — Торпедообразная форма Способствует образованию завихрений потоков воды при движении. акулы дельфины
Обтекаемая форма тела Сокол-сапсан Обтекаемая форма тела способствует быстрому передвижению в воздушной среде. орел пингвин Утка лебедь дятел Птицы: соколсапсан, орел, пингвин, утка, лебедь, дятел.
Форма тела Палочники сверчок цикады кобылка -Сучковидная и листовидная форма тела делает организм незаметным среди объектов окружающей среды.
Форма тела -Причудливая рыба-клоун удильщик морская игла форма тела Скрывает среди водорослей, коралловых полипов представителей морских глубин. рыба-жаба.
Приспособленность. Окраска тела. ВИДЫ ПРИСПОСОБЛЕННОСТИ ЗНАЧЕНИЕ ПРИМЕРЫ Окраска тела: — покровительственная Скрывает на фоне окружающей среды Заяц-беляк, белая куропатка, зеленый кузнечик, тли — расчленяющая То же на фоне полос света и тени Зебры, тигры — предостерегающая Сохранение численности видов, обладающих ядовитыми, обжигающими, жалящими свойствами Пчелы, осы, жуки-нарывники, гусеницы, божья коровка Мимикрия (подражание беззащитных животных хорошо Защита от истребления защищенным и обладающим предостерегающей окраской) Иглы, колючки, кристаллы щавелевокислого калия, Защита от поедания травоядными накапливающегося в колючках или животными листьях растений Твердые покровы тела Защита от поедания плотоядными животными Осовидки, пчеловидки, шмелевидки; яйца, откладываемые кукушкой Кактусы, шиповник, боярышник, крапива Жуки, крабы, двустворчатые моллюски, черепахи, броненосцы
Меняющаяся и расчлененная окраска тигры хамелеон осьминог Зебры кальмар камбала — Меняющаяся покровительственная окраска Скрывает на фоне окружающей среды. — Расчленяющая Скрывает в окружающей среде на фоне полос света и тени.
Однотонная окраска зеленый кузнечик Гнездо малого зуйка тля антилопа лев Однотонная покровительственная окраска Скрывает на фоне окружающей среды.
Приспособительная окраска меняющаяся покровительстве нная окраска Скрывает на фоне окружающей среды, меняется в зависимости от сезона. песец горностай заяц
Предостерегающая окраска Сохранение численности видов, обладающих ядовитыми, обжигающими, жалящими свойствами. Пчелы божья коровка жуки нарывники гусеницы клоп-солдатик
Мимикрия Защита от истребления Оса — Осовидки пчела шмель Бабочка данаида пчеловидки шмелевидки Бабочка нимфали
Приспособленность. Защитные приспособления ВИДЫ ПРИСПОСОБЛЕННОСТИ ЗНАЧЕНИЕ ПРИМЕРЫ Защитные приспособления: Иглы, колючки, кристаллы щавелевокислого калия, Защита от поедания травоядными Кактусы, шиповник, накапливающегося в колючках или животными боярышник, крапива листьях растений Твердые покровы тела Иглы Жуки, крабы, двустворчатые Защита от поедания плотоядными моллюски, черепахи, животными броненосцы Защита от поедания плотоядными животными Ехидны, дикобразы, ежи
Защитные приспособления Кактусы шиповник крапива боярышник — Иглы, колючки, кристаллы щавелевокислого калия, накапливающегося в колючках или листьях растений Служат защитой от поедания травоядными животными.
Защитные приспособления. Иглы. ежи дикобразы Ехидны Рыба-еж — Иглы, колючки, часто накапливающие ядовитые вещества. Защита от поедания плотоядными животными.
Твердые покровы Твердые покровы тела Защита от поедания плотоядными животными. Жуки крабы черепахи двустворчатые моллюски броненосцы
Приспособленность. Приспособительное поведение ВИДЫ ПРИСПОСОБЛЕННОСТИ ЗНАЧЕНИЕ ПРИМЕРЫ Приспособительное поведение: — замирание — угрожающая поза — запасание корма Защита от поедания травоядными животными То же Переживание бескормицы Опоссумы, некоторые жуки, земноводные, птицы Бородатая ящерица, ушастая круглоголовка Кедровка, сойка, бурундук, белка, пищуха (сеноставка)
Замирающая поза Опоссумы жуки -Замирание это имитация ранения или смерти. жаба серая тритон гребенчатый выпь дроффа
Угрожающая поза Бородатая ящерица ушастая круглоголовка Скунс Богомол — Угрожающая поза отпугивающее поведение, часто очень характерно для ядовитых и жалящих форм.
Приспособленность. Забота о потомстве. ВИДЫ ПРИСПОСОБЛЕННОСТИ ЗНАЧЕНИЕ ПРИМЕРЫ Забота о потомстве: — вынашивание икры в ротовой полости, Сохранение потомства в складке кожи на животе Самцы тиляпии, морского сомика, морского конька — постройка гнезда и выведение в нем Сохранение потомства Некоторые рыбы (колюшки, петушки, макроподы), птицы, белки, мыши-малютки — выкармливание потомства, Сохранение потомства обеспечение будущего потомства пищей Птицы, млекопитающие, жуки скарабеи, наездники.
Вынашивание потомства — Вынашивание икры в ротовой полости, в складке кожи на животе Сохранение потомства морской сомик Самцы тиляпии морской конек
Выведение потомства иволга петушки — Постройка гнезда и выведение в нем потомства. Сохранение потомства. колюшки трясогузка аист белки мыши-малютки
Выкармливание потомства львы лебедь — Выкармливание потомства, обеспечение будущего потомства пищей. Енот-полоскун Жуки -скарабеи. фламинго наездники.
Слайд 1
Слайд 2
Слайд 3
Слайд 4
Слайд 5
Слайд 6
Слайд 7
Слайд 8
Слайд 9
Слайд 10
Слайд 11
Слайд 12
Слайд 13
Слайд 14
Слайд 15
Слайд 16
Слайд 17
Слайд 18
Слайд 19
Слайд 20
Слайд 21
Слайд 22
Слайд 23
Слайд 24
Слайд 25
Слайд 26
Слайд 27
Слайд 28
Слайд 29
Слайд 30
Слайд 31
Слайд 32
Слайд 33
Презентацию на тему «Приспособленность организмов» можно скачать абсолютно бесплатно на нашем сайте. Предмет проекта: Биология. Красочные слайды и иллюстрации помогут вам заинтересовать своих одноклассников или аудиторию. Для просмотра содержимого воспользуйтесь плеером, или если вы хотите скачать доклад — нажмите на соответствующий текст под плеером. Презентация содержит 33 слайд(ов).
Слайды презентации
Слайд 1
Приспособленность организмов к условиям внешней среды как результат действия естественного отбора
Составитель Большаков С. В.
Слайд 2
Виды растений и животных удивительно приспособлены к условиям среды, в которых они обитают. Известно огромное количество самых разнообразных особенностей строения, обеспечивающих высокий уровень приспособленности вида к среде. В понятие «приспособленность вида» входят не только внешние признаки, но и соответствие строения внутренних органов выполняемым ими функциям, например длинный и сложно устроенный пищеварительный тракт животных, питающихся растительной пищей (жвачные). Соответствие физиологических функций организма условиям обитания, их сложность и разнообразие также входят в понятие приспособленности.
Слайд 3
Приспособительные особенности строения, окраски тела и поведения животных. У животных приспособительной является форма тела. Хорошо известен облик водного млекопитающего дельфина. Его движения легки и точны. Самостоятельная скорость движения в воде достигает 40 км/ч. Нередко описывают случаи, как дельфины сопровождают быстроходные морские суда, например эсминцы, движущиеся со скоростью 65 км/ч..
http://www.botik.ru/~yz/rrp/puzlyary/prize/index.koi8.html
Слайд 4
Объясняется это тем, что дельфины пристраиваются к носу судна и используют гидродинамическую силу корабельных волн. Но это не их естественная скорость. Плотность воды в 800 раз выше плотности воздуха. Как дельфину удается преодолеть её? Помимо других особенностей строения идеальной приспособленности дельфина к среде обитания и образу жизни способствует форма тела. Торпедовидная форма тела позволяет избежать образования завихрения потоков воды, обтекающих дельфина.
http://desktop.kazansoft.ru/preview/cat1-117.html
Слайд 5
Слайд 6
Обтекаемая форма тела способствует быстрому передвижению животных и в воздушной среде. Маховые и контурные перья, покрывающие тело птицы, полностью сглаживают его форму. Птицы лишены выступающих ушных раковин, в полёте они обычно втягивают ноги. В результате птицы по быстроте намного превосходят всех других животных. Например, сокол-сапсан пикирует на свою жертву со скоростью до 290 км/ч.
Слайд 7
Птицы быстро двигаются даже в воде. Наблюдали антарктического пингвина, плывущего под водой со скоростью около 35 км/ч.
Пингвин Адели
Слайд 8
У животных ведущих скрытный, затаивающийся образ жизни, полезными оказываются приспособления, придающие им сходство с предметами окружающей среды. Причудливая форма тела у рыб, обитающих в зарослях водорослей, помогает им успешно скрываться от врагов.
http://forum.allgaz.ru/showthread.php?t=10009&page=4
Слайд 9
Сходство с предметами среды обитания широко распространено у насекомых. Известны жуки, своим внешним видом напоминающие лишайники, цикады, сходные с типами тех кустарников, среди которых они живут. Насекомые-палочники похожи на небольшую бурую или зеленую веточку, а прямокрылые насекомые имитируют лист.
Палочники http://macroid.ru/showphoto.php?photo=11879
Слайд 10
Слайд 11
Средством защиты от врагов служит и покровительственная окраска. Птицы, насиживающие яйца на земле, сливаются с окружающим фоном. Мало заметны и их яйца, имеющие пигментированную скорлупу, и вылупляющиеся из них птенцы. Защитный характер пигментации яиц подтверждается тем, что у видов, чьи яйца недоступны для врагов — крупных хищников, или у птиц, откладывающих яйца на скалах или закапывающих их в землю, покровительственная окраска скорлупы не развивается.
http://kizhi.karelia.ru/gallery/life_moment/index_e.php?i=16
Слайд 12
Покровительственная окраска широко распространена среди самых различных животных. Гусеницы бабочек часто зеленые, под цвет листьев, или темные, под цвет коры или земли. Донные рыбы обычно окрашены под цвет песчаного дна (скаты и камбалы). При этом камбалы способны еще менять окраску в зависимости от цвета окружающего фона.
Камбала полярная
Слайд 13
Способность менять окраску путем перераспределения пигмента в покровах тела известна и у наземных животных (хамелеон).
Хамелеоны http://ru.wikipedia.org/wiki/Хамелеоны
Слайд 14
Животные пустынь имеют, как правило, желто-бурую или песочно-желтую окраску.
Королевская змея пустынная (Lampropeltis getula… http://www.terrariy.ru/Anim/Snake/Desert_p.htm
Слайд 15
Слайд 16
Если фон среды не остается постоянным в зависимости от сезона года, многие животные меняют окраску. Например, обитатели средних и высоких широт (песец, заяц, горностай, белая куропатка) зимой имеют белую окраску, что делает их незаметными на снегу.
Слайд 17
Однако нередко у животных наблюдается не скрывающая окраска тела, а, напротив, привлекающая внимание, демаскирующая. Такая окраска свойственна ядовитым, обжигающим или жалящим насекомым: пчелам, осам, жукам-нарывникам.
Пчела медоносная
Слайд 18
Божью коровку, очень заметную, птицы никогда не склевывают из-за выделяемого насекомым ядовитого секрета.
Фотографии божьих коровок фото 14 http://basik.ru/macro/1778/
Слайд 19
Яркую предупреждающую окраску имеют несъедобные гусеницы, многие ядовитые змеи. Яркая окраска заранее предупреждает хищника о бесполезности и опасности нападения. Путем «проб и ошибок» хищники быстро приучаются избегать нападения на жертву с предупреждающей окраской.
Ядовитая змея кобра. http://900igr.net/Detskie_prezentatsii/Biologija.Morskie_zhiteli/Zmei_1.files/detskie_kartinki_zhivotnykh_020_JAdovitaja_zmeja_kobra_vsta.html
Слайд 20
Защитное действие покровительственной или предупреждающей окраски повышается при сочетании ее с соответствующим поведением. Например, выпь гнездится в камышах. В минуты опасности она вытягивает шею, поднимает вверх голову и замирает. В такой позе ее трудно обнаружить даже на близком расстоянии.
Выпь большая
Слайд 21
Многие другие животные, не обладающие средствами активной защиты, в случае опасности принимают позу покоя и замирают (насекомые, рыбы, амфибии, птицы). Предостерегающая окраска у животных, наоборот, сочетается с демонстративным поведением, отпугивающим хищника. Эффективность предостерегающей окраски явилась причиной очень интересного явления — подражания, или мимикрии. Мимикрией называется сходство беззащитного или съедобного вида с одним или несколькими неродственными видами, хорошо защищенными и обладающими предостерегающей окраской. С божьей коровкой размерами, формой тела и распределением пигментных пятен очень сходен один из видов тараканов.
Слайд 22
Некоторые съедобные бабочки подражают формой тела и окраской ядовитым бабочкам, мухи — осам. Возникновение мимикрии связано с накоплением под контролем естественного отбора мелких удачных мутаций у съедобных видов в условиях их совместного обитания с несъедобными.
Пример мимикрии: муха семейства журчалок… http://www.enci.ru/Мимикрия
Слайд 23
Понятно, что подражание одних видов другим оправдано: истреблению подвергается значительно меньшая часть особей и того вида, который послужил моделью, и вида-подражателя. Необходимо, однако, чтобы численность вида подражателя была значительно меньше численности модели. В противном случае мимикрия не приносит пользы: у хищника не будет вырабатываться стойкий условный рефлекс на форму или окраску, которую следует избегать. Каким же образом численность вида-имитатора поддерживается на низком уровне? Оказалось, что генофонд этих видов насыщен летальными мутациями. В гомозиготном состоянии эти мутации вызывают гибель насекомых, в результате чего высокий процент особей не доживает до половозрелого состояния.
Яйцо кукушки в кладке синего соловья. http://kniiekotija.ucoz.ru/forum/58-145-3
Слайд 25
Кроме защитной окраски, у животных и растений наблюдаются и другие средства защиты. У растений нередко образуются иглы и колючки, защищающие их от поедания травоядными животными (кактусы, шиповник, боярышник, облепиха и др.).
http://www.tiensmed.ru/news/shipovnik-wkti/
Слайд 26
Такую же роль играют ядовитые вещества, обжигающие волоски, например у крапивы. Кристаллы щавелевокислого кальция, накапливающиеся в шипах некоторых растений предохраняют их от поедания гусеницами, улитками и даже грызунами.
Крапива двудомная
Слайд 27
Образования в виде твердого хитинового покрова у членистоногих (жуки, крабы), раковин у моллюсков, чешуи у крокодилов, панциря у броненосцев и черепах хорошо защищают их от многих врагов. Этому же служат иглы у ежа и дикобраза. Все эти приспособления могли появиться лишь в результате естественного отбора, т. е. преимущественного выживания лучше защищенных особей.
Черепаха слоновая
Слайд 28
Для выживания организмов в борьбе за существование большое значение имеет приспособительное поведение. Помимо затаивания или демонстративного, отпугивающего поведения при приближении врага существует много других вариантов приспособительного поведения, обеспечивающего выживаемость взрослых особей или молоди. Сюда относится запасание корма на неблагоприятный сезон года. Особенно это относится к грызунам. Например, полевка-экономка, распространенная в таежной зоне, собирает зерна злаков, сухую траву, корешки — всего до 10 кг.
Полевка-экономка — Microtus oeconomus (Pallas http://www.apus.ru/site.xp/049051056048124053054050052.html
Слайд 29
Роющие грызуны (слепыши и др.) накапливают кусочки корней дуба, желуди, картофель, степной горошек — до 14 кг.
Thread. skajazz. слепыши. http://fon-shcmal.livejournal.com/1840.html
Слайд 30
Большая песчанка, живущая в пустынях Средней Азии, в начале лета срезает траву и затаскивает ее в норы или оставляет на поверхности в виде стожков. Корм этот используется во второй половине лета, осенью и зимой.
Большие песчанки- типичные обитатели пустыни. http://elementy.ru/news/430180
Слайд 31
Речной бобр собирает обрубки деревьев, веток и пр., которые складывает в воду возле своего жилища. Склады эти могут достигать объема 20 метров куб.
Бобры являются самыми известными «строителями» дамб в реках и потоках, и… http://www.ff18.ru/bobry/bobry.html
Советы как сделать хороший доклад презентации или проекта
- Постарайтесь вовлечь аудиторию в рассказ, настройте взаимодействие с аудиторией с помощью наводящих вопросов, игровой части, не бойтесь пошутить и искренне улыбнуться (где это уместно).
- Старайтесь объяснять слайд своими словами, добавлять дополнительные интересные факты, не нужно просто читать информацию со слайдов, ее аудитория может прочитать и сама.
- Не нужно перегружать слайды Вашего проекта текстовыми блоками, больше иллюстраций и минимум текста позволят лучше донести информацию и привлечь внимание. На слайде должна быть только ключевая информация, остальное лучше рассказать слушателям устно.
- Текст должен быть хорошо читаемым, иначе аудитория не сможет увидеть подаваемую информацию, будет сильно отвлекаться от рассказа, пытаясь хоть что-то разобрать, или вовсе утратит весь интерес. Для этого нужно правильно подобрать шрифт, учитывая, где и как будет происходить трансляция презентации, а также правильно подобрать сочетание фона и текста.
- Важно провести репетицию Вашего доклада, продумать, как Вы поздороваетесь с аудиторией, что скажете первым, как закончите презентацию. Все приходит с опытом.
- Правильно подберите наряд, т.к. одежда докладчика также играет большую роль в восприятии его выступления.
- Старайтесь говорить уверенно, плавно и связно.
- Старайтесь получить удовольствие от выступления, тогда Вы сможете быть более непринужденным и будете меньше волноваться.
Растения и животные приспособлены к условиям среды, в которых они обитают. В понятие «приспособленность вида» входит не только внешние признаки, но и соответствие строения внутренних органов выполняемым ими функциям (например, длинный и сложно устроенный пищеварительный тракт жвачных животных, питающихся растительной пищей). Соответствие физиологических функций организма условиям их обитания, их сложность и разнообразие также входят в понятие приспособленности.
Не вызывает сомнений и согласованность деятельности отдельных частей и систем внутри самого организма. Долгое время подобная целесообразность строения служила аргументом в пользу божественного происхождения живой природы. Но теория эволюции Дарвина сумела объяснить это с материалистических позиций. В настоящее время эволюционный подход к рассмотрению биологических закономерностей служит естественнонаучной базой для объяснения целесообразности строения живых организмов и их приспособленности к условиям обитания.
Приспособительные особенности строения, окраски тела и поведения животных
Обтекаемая форма тела — приспособление к преодолению сопротивления воздуха (для птиц) и воды (для водных животных) при передвижении в этих средах. Такая форма позволяет развивать большую скорость движения и экономить при этом энергию.
Покровительственная окраска и форма тела — окраска и форма тела животного, способствующие сохранению его жизни в борьбе за существование. Покровительственная окраска и форма тела очень разнообразны и встречаются среди многих групп беспозвоночных и позвоночных животных. Различают 3 типа покровительственной окраски и формы тела: маскировку , демонстрацию и мимикрию .
Маскировка — приспособление, при котором форма тела и окраска животного сливаются с окружающими предметами. Например, гусеницы некоторых бабочек по форме тела и окраске напоминают веточки.
Животные, обитающие в траве, имеют зелёную окраску: ящерицы, кузнечики, гусеницы , обитатели пустынь — жёлтую или бурую: пустынная саранча, ушастая круглоголовка, сайгак .
Некоторые животные меняют окраску в течение онтогенеза (детёныши и взрослые особи тюленей), в различные сезоны года (песец, заяц-беляк, белка и многие другие).
Некоторые животные способны изменять окраску в соответствии с фоном, что достигается перераспределением пигментов в хроматофорах покровов тела (каракатицы, камбалы, агамы и др.). Маскирующая окраска обычно сочетается с позой покоя.
Маскировка способствует успеху в борьбе за существование.
Расчленяющая окраска (дизруптивная окраска) — окраска с наличием контрастных полос или пятен, разбивающих контур тела на отдельные участки, благодаря чему животное становится незаметным на окружающем фоне.
Расчленяющая окраска часто сочетается с подражающей поверхности и фону и встречается у многих животных: жираф, зебр, бурундуков , у некоторых рыб, земноводных, пресмыкающихся, из насекомых— у саранчовых, многих бабочек и их гусениц.
Скрадывающая окраска основана на эффекте противотени: наиболее ярко освещаемые участки тела окрашены темнее менее освещаемых: при этом окраска кажется более монотонной, а очертания животного сливаются с фоном. Такая окраска (тёмная спина — светлое брюхо) характерна для большинства рыб и других обитателей толщи воды, для многих птиц и некоторых млекопитающих (олени, зайцы ).
Предупреждающая окраска — вид покровительственной окраски и формы, при которой несъедобные животные обладают яркой, броской, порой — пёстрой, окраской. Эти животные хорошо заметны контрастным сочетаниям цветов (чёрного, красного, белого; оранжевого, белого, чёрного и т. д.). Многие насекомые обладают предупреждающей окраской, например клопы-солдатики, божьи коровки, бронзовки, листоеды, нарывники , различные бабочки — пестрянки, медведицы и др.
Среди позвоночных животных предупреждающая окраска наблюдается у рыб, саламандр, жаб жерлянок, у некоторых птиц (дронго ), а среди млекопитающих — например, у американского скунса . Заметность животных с предупреждающей окраской — их преимущество, поскольку они неузнаваемы и не подвергаются нападению хищников. Предупреждающая окраска способствует выживаемости вида в борьбе за существование и является результатом действия естественного отбора.
Мимикрия (греч. мимикос — подражательный) — подражательное сходство незащищённого организма с защищённым или несъедобным.
У животных мимикрия способствует выживанию в борьбе за существование. Мимикрия может быть не только направлена на пассивную защиту, но и служить орудием нападения, подманивания добычи.
Демонстративное поведение — одно из средств общения у животных. Совершая различные телодвижения, например, птицы в брачный период показывают друг другу те или иные участки оперения, с яркой сигнальной окраской, которая несет информацию.
Демонстративное поведение используется для привлечения брачных партнеров, при ухаживании, конфликтах с соперниками, защите гнезд, общении с птенцами, захвате и отстаивании территорий, а также в качестве средства предупреждения об опасности.
Большое значение для выживания организмов имеет приспособительное поведение . Сезонные миграции животных — пример приспособительного поведения.
Сезонная линька связана с сезонными изменениями условий существования животных. У животных, не впадающих в зимнюю спячку, ежегодно наблюдается осенняя и весенняя линька.
При осенней линьке теплопроводный волосяной покров сменяется густым, тёплым мехом. При весенней линьке одновременно с заменой покрова у многих животных происходит слущивание верхней части рогового слоя эпидермиса.
Запасание кормов животными — важный инстинкт, наиболее развитый у обитателей холодных и умеренных широт с резкими сезонными изменениями кормовых условий. Наблюдается у многих беспозвоночных, у некоторых птиц и особенно часто у млекопитающих. Из беспозвоночных запасают корм некоторые пауки, крабы, раки и многие насекомые.
Из птиц корма запасают только зимующие. Большинство птиц использует запасы зимой как дополнительное питание.
Из млекопитающих запасают корм некоторые хищники, пищухи и многие грызуны. Запасы используются зимой или весной после пробуждения от спячки или зимнего сна.
Степной хорь складывает в нору сусликов, горностай — водяных крыс, мышей, лягушек, ласка — мелких грызунов. Многие пищухи заготовляют сено, складывая его в стожки или в щели между камнями. Белка запасает грибы, орехи и жёлуди. Бурундук затаскивает к себе в нору орехи, зёрна, лесная мышь — семена, речной бобр — ветви и корневища, погружая их в воду около входа в нору.
Конспект урока по биологии 9 класс
Тема: «Приспособительные особенности строения, окраски тела и поведения животных»
Цель: познакомиться с разными видами приспособленности живых организмов к среде обитания, понять относительный характер приспособленности.
Задачи:
Обучающие:
Сформировать понятие о механизмах возникновения приспособленности как результата эволюции;
Продолжить развитие умений использовать знания теоретических закономерностей для объяснения явлений, наблюдаемых в живой природе;
Сформировать конкретные знания о приспособительных особенностях строения, окраски тела и поведения животных, раскрыть относительный характер приспособлений
Развивающие:
Развивать интерес к изучению биологии, расширять кругозор о закономерностях в природе с помощью ситуативного общения;
Развивать творческие способности учащихся посредством самостоятельного создания компьютерной презентации используя найденный в Интернете иллюстративный материал.
развивать интеллектуальную сферу: внимание, память, речь, мышление;
Воспитательные:
продолжить формирование у школьников экологической культуры, убеждения в необходимости сохранения видового многообразия растений и животных.
подвести к выводам о естественных причинах формирования приспособлений, с использованием учения о движущих силах эволюции;
расширить кругозор учащихся.
№ урока | Тема урока | УУД | Предметные результаты | Метапредметные результаты | Личностные результаты |
Приспособленность организмов к условиям внешней среды как результат действия естественного отбора | Информационная компетентность извлечение первичной информации (1уровень), Установление причинно-следственных связей между приспособлениями организмов и среды обитания. (2 уровень) | формировать понятие об адаптациях организмов, научить называть и определять разнообразные адаптации развить умения применять полученные знания для определения приспособленности организмов. | Продолжить освоение важнейшими предметными умениями (аргументировать свой ответ, давать определение терминам, анализировать, обобщать полученную информацию) продолжить формировать развитие умений работать с различными информационными источниками и объектами. | Обеспечение формирования критического мышления через чтение научно-популярной литературы, способствовать развитию умения выражать собственное мнение, воспитывать ценностное отношение к живой природе. |
ход урока
1. Актуализация прежних знаний
К.О.З.
С какими силами эволюции мы познакомились?
Какую силу эволюции Ч.Дарвин считал основной?
Какие организмы выживают и дают потомство в результате естественного отбора?
2. Изучение нового материала.
В настоящее время на нашей планете обитает несколько миллионов видов живых организмов, каждый из которых по-своему уникальный. Давайте же выясним, что такое приспособленность организмов к среде обитания.
Совместная постановка целей урока
В ходе беседы выясняем понятие приспособленности, оно появляется на сладе, ребята записывают его в тетрадь (это же определение на информационной карте)
Приспособленность организмов, или адаптации (от лат. adaptatio — приспособление, прилаживание), представляют собой совокупности тех особенностей строения, физиологии и поведения, которые обеспечивают для данного вида возможность специфического образа жизни в определенных условиях внешней среды.
К.О.З.
Как вы думаете с помощью чего можно приспособиться к среде обитания?
У животных приспособительной является форма тела. Хорошо известен облик водного млекопитающего дельфина. Его движения легки и точны, скорость движения в воде достигает 40 км/ч. Плотность воды в 800 раз выше плотности воздуха. Как дельфину удаётся преодолеть её? Торпедовидная обтекаемая форма тела, отсутствие ушных раковин позволяют избежать завихрения потоков воды, окружающих дельфина, снижают трение. Подобная форма тела у многих водных животных: акул, китов, тюленей. Обтекаемая форма тела способствует быстрому передвижению животных и в воздушной среде. Маховые и контурные перья, покрывающие тело птицы, полностью сглаживают его форму. Птицы лишены выступающих ушных раковин, в полёте они обычно втягивают ноги. В результате птицы по быстроте намного превосходят всех других животных. Птицы быстро двигаются даже в воде. Наблюдали арктического пингвина, плывущего под водой со скоростью 35 км/ч.
Организменные адаптации – запись в тетради.
К.О.З. Еще Ч. Дарвин подчеркивал, что все приспособления, сколько бы совершенными они не были, носят относительный характер, т.е. полезны лишь в типичной среде обитания.
Например, дятел легко перемещается по стволам деревьев, но его конечности плохо приспособлены для перемещения по поверхности почвы.
Водоплавающие птицы плохо перемещаются по суше.
К.О.З.
Давайте вспомним о такой силе эволюции как борьба за существование. Какие формы борьбы за существование вы знаете?
В чем особенность межвидовой борьбы за существование, между кем она происходит?
как должны приспособиться хищники и их жертвы?
СЛАЙДЫ №
ученики записывают определения, делают выводы об относительном характере любой адаптации.
покровительственная окраска
сплошная
разбитая
изменение окраски тела
предостерегающая окраска
Однако нередко у животных наблюдается не скрывающая окраска тела, а, напротив, привлекающая внимание, демаскирующая. Такая форма приспособления называется предупреждающая окраска. Она свойственна большинству жалящих, выделяющих ядовитые вещества, отвратительно пахнущих или омерзительных на вкус животных. Подобно стоп-сигналам, эти рисунки и сочетания цветов должны легко распознаваться животными. Они означают: “Опасно!”, “Не подходи!”, “Со мной лучше не связываться!”. Божью коровку, очень заметную, птицы никогда не склёвывают из-за выделяемого насекомым ядовитого секрета. Яркую предупреждающую окраску имеют несъедобные гусеницы, многие ядовитые змеи. Среди земноводных есть настоящие щёголи. Они эффектно окрашены, нередко медлительны, ведут дневной образ жизни и даже не пытаются прятаться от хищников в отличие от своих более многочисленных замаскированных родичей, которые отправляются на поиски корма по ночам, когда они менее заметны. Наиболее своеобразны среди земноводных щеголей, пожалуй, древолазы, обитатели Центральной и Южной Америки. Их кожные железы вырабатывают мощные парализующие яды, так что хищник, попробовавший закусить такой лягушкой и оставшийся в живых, ассоциирует пережитые неприятные минуты с её яркой расцветкой и в дальнейшем старательно избегает ей подобных. Среди примерно ста тысяч видов, составляющих отряд чешуекрылых, или бабочек, медведицы принадлежат не только к наиболее привычным, но и к самым красивым. У неё на редкость эффективная предупреждающая окраска – оранжево-чёрная и жёлто-чёрная с узорами из пятен и полосок. Медведица очень хороша собой, но ядовита. Специальные железы вырабатывают сильные токсины, поступающие в кровоток бабочки. Другие железы содержат жидкость с неприятным предостерегающим запахом. В тропических прибрежных водах Австралии, Новой Гвинеи, Индонезии и Филиппин обитает маленький (до 20 см. в длину вместе со щупальцами) синекольчатый осьминог. Ярко-оранжевые круглые пятна окаймлены характерными синими кольцами. Как и все представители рода, синекольчатый осьминог обладает удивительной способностью к регенерации, и, потеряв в бою одну или несколько из своих восьми щупалец, довольно быстро может отрастить новые. Насколько этот осьминожка красив, настолько же и ядовит. В слюне животного содержится сильнейшийнейротоксин. Укус синекольчатого осьминога смертелен. Яд практически моментально парализует нервную систему любого живого существа, и противоядия от него не существует.
мимикрия
Эффективность предостерегающей окраски явилась причиной очень интересного явления – подражания, или мимикрии. Мимикрия – это подражание менее защищённого организма одного вида более защищённому организму другого вида. Это подражание может проявляться в форме тела, окраске и т.д. Покрытая предостерегающими полосками, но совершенно безобидная муха журчалка извлекает нектар из цветка, как и медоносные пчелы, обладающие грозным жалом. Мимикрия журчалки не ограничивается окраской, но включает и поведение. Журчалки подражают звукам, издаваемым пчёлами и осами, и, если их потревожить, угрожающе жужжат. Всё это вместе гарантирует журчалке неприкосновенность. Красивая бабочка данаида обязана своей несъедобностью тому, что её гусеницы питаются листьями ядовитого латука, опасного для скота и других позвоночных. Крылатые хищники быстро научились не трогать данаид, а заодно и их подражательницу, одну из нимфалид – лишь слегка невкусную. Бабочка-стеклянница удивительно похожа на осу. Крылья её совершенно прозрачны, так как чешуек, покрывающих крылья бабочек, у неё нет. При полёте она жужжит подобно осам, и летает так же стремительно и беспокойно, как и они. Уж имитирует окраску гадюки, его выдают только жёлтые пятна на голове. Немало подражателей приобрели ядовитые коралловые змеи. Например, аризонская королевская змея, которая не ядовита.
маскировка
У животных ведущих скрытый, затаивающийся образ жизни, полезными оказываются приспособления, придающие им сходство с предметами окружающей среды – маскировка. Например, гусеницы бабочки пяденицы по форме тела и окрасу напоминают сучки. Насекомые-палочники похожи на небольшую бурую или зелёную веточку, некоторые бабочки напоминают сухие листья, а пауки подражают колючкам. Великие мастера маскировки своим успехом во многом обязаны способности замереть в момент, когда им грозит нападение или они сами готовятся схватить добычу. Среди животных особенно разнообразны те, кто так или иначе подражает цветкам. Например, цветочные богомолы, настолько уподобляются тем или иным частям растения, что обманутые сходством другие насекомые опускаются прямо на них и попадают в объятия хищника.
Учащиеся записывают определения и делают выводы об относительном характере любой адаптации .
К.О.З. Как же возникают столь совершенные адаптации?
Ключ к разгадке лежит в сложнейшем процессе естественного отбора. Например, дальний пращур бабочки, теперь почти неотличимой от сухого листа, появился на свет со случайным набором генов, придавшим ему чуть большее сходство с сухим листком. А потому птицам было несколько труднее обнаружить эту бабочку среди сухих листьев, и в результате она и ей подобные особи выживали в большем количестве. Следовательно, они оставляли больше потомства. А признак “сухой лист” становился всё более чётким и распространённым. Любые признаки возникают в результате мутаций. Может произойти одна крупная мутация, а может огромное множество мелких, что бывает гораздо чаще. Те из них, которые повышают жизнеспособность, передаются последующим поколениям, закрепляются и становятся приспособлениями. Каждое приспособление вырабатывается на основе наследственной изменчивости в процессе борьбы за существование и отбора в ряду поколений.
Какие выводы можно сделать из всего вышеизложенного?
1.Общая приспособленность организмов к условиям среды складывается из множества отдельных адаптаций самого разного масштаба.
2.Все приспособления возникают в ходе эволюции в результате естественного отбора.
3.Любое приспособление относительно.
Таким образом, приспособленность – это относительная целесообразность строения и функций организма, являющаяся результатом естественного отбора.
Сайт учителей биологии МБОУ Лицей № 2 города Воронежа
Растения и животные приспособлены к условиям среды, в которых они обитают. В понятие «приспособленность вида» входит не только внешние признаки, но и соответствие строения внутренних органов выполняемым ими функциям (например, длинный и сложно устроенный пищеварительный тракт жвачных животных, питающихся растительной пищей). Соответствие физиологических функций организма условиям их обитания, их сложность и разнообразие также входят в понятие приспособленности.
Не вызывает сомнений и согласованность деятельности отдельных частей и систем внутри самого организма. Долгое время подобная целесообразность строения служила аргументом в пользу божественного происхождения живой природы. Но теория эволюции Дарвина сумела объяснить это с материалистических позиций. В настоящее время эволюционный подход к рассмотрению биологических закономерностей служит естественнонаучной базой для объяснения целесообразности строения живых организмов и их приспособленности к условиям обитания.
Приспособительные особенности строения, окраски тела и поведения животных
Обтекаемая форма тела — приспособление к преодолению сопротивления воздуха (для птиц) и воды (для водных животных) при передвижении в этих средах. Такая форма позволяет развивать большую скорость движения и экономить при этом энергию.
Покровительственная окраска и форма тела — окраска и форма тела животного, способствующие сохранению его жизни в борьбе за существование. Покровительственная окраска и форма тела очень разнообразны и встречаются среди многих групп беспозвоночных и позвоночных животных. Различают 3 типа покровительственной окраски и формы тела: маскировку, демонстрацию и мимикрию.
Маскировка — приспособление, при котором форма тела и окраска животного сливаются с окружающими предметами. Например, гусеницы некоторых бабочек по форме тела и окраске напоминают веточки.
Животные, обитающие в траве, имеют зелёную окраску: ящерицы, кузнечики, гусеницы, обитатели пустынь — жёлтую или бурую: пустынная саранча, ушастая круглоголовка, сайгак.
Некоторые животные меняют окраску в течение онтогенеза (детёныши и взрослые особи тюленей), в различные сезоны года (песец, заяц-беляк, белка и многие другие).
Некоторые животные способны изменять окраску в соответствии с фоном, что достигается перераспределением пигментов в хроматофорах покровов тела (каракатицы, камбалы, агамы и др.). Маскирующая окраска обычно сочетается с позой покоя.
Маскировка способствует успеху в борьбе за существование.
Расчленяющая окраска (дизруптивная окраска) — окраска с наличием контрастных полос или пятен, разбивающих контур тела на отдельные участки, благодаря чему животное становится незаметным на окружающем фоне.
Расчленяющая окраска часто сочетается с подражающей поверхности и фону и встречается у многих животных: жираф, зебр, бурундуков, у некоторых рыб, земноводных, пресмыкающихся, из насекомых— у саранчовых, многих бабочек и их гусениц.
Скрадывающая окраска основана на эффекте противотени: наиболее ярко освещаемые участки тела окрашены темнее менее освещаемых: при этом окраска кажется более монотонной, а очертания животного сливаются с фоном. Такая окраска (тёмная спина — светлое брюхо) характерна для большинства рыб и других обитателей толщи воды, для многих птиц и некоторых млекопитающих (олени, зайцы).
Предупреждающая окраска — вид покровительственной окраски и формы, при которой несъедобные животные обладают яркой, броской, порой — пёстрой, окраской. Эти животные хорошо заметны контрастным сочетаниям цветов (чёрного, красного, белого; оранжевого, белого, чёрного и т. д.). Многие насекомые обладают предупреждающей окраской, например клопы-солдатики, божьи коровки, бронзовки, листоеды, нарывники, различные бабочки — пестрянки, медведицы и др.
Среди позвоночных животных предупреждающая окраска наблюдается у рыб, саламандр, жаб жерлянок, у некоторых птиц (дронго), а среди млекопитающих — например, у американского скунса. Заметность животных с предупреждающей окраской — их преимущество, поскольку они неузнаваемы и не подвергаются нападению хищников. Предупреждающая окраска способствует выживаемости вида в борьбе за существование и является результатом действия естественного отбора.
Мимикрия (греч. мимикос — подражательный) — подражательное сходство незащищённого организма с защищённым или несъедобным.
У животных мимикрия способствует выживанию в борьбе за существование. Мимикрия может быть не только направлена на пассивную защиту, но и служить орудием нападения, подманивания добычи.
Гнездовой паразитизм — тип паразитизма, при котором паразит использует хозяина, которым может являться особь как своего (внутривидовой паразитизм), так и другого вида (межвидовой паразизм), с целью выращивания своего потомства. В результате такого взаимодействия паразит избавлен от необходимости строить гнездо и выкармливать потомство, а может тратить больше времени на добычу пищи и размножение.
Гнездовой паразитизм встречается у птиц, рыб, насекомых, но в целом является малораспространённым явлением.
Демонстративное поведение — одно из средств общения у животных. Совершая различные телодвижения, например, птицы в брачный период показывают друг другу те или иные участки оперения, с яркой сигнальной окраской, которая несет информацию.
Демонстративное поведение используется для привлечения брачных партнеров, при ухаживании, конфликтах с соперниками, защите гнезд, общении с птенцами, захвате и отстаивании территорий, а также в качестве средства предупреждения об опасности.
Большое значение для выживания организмов имеет приспособительное поведение. Сезонные миграции животных — пример приспособительного поведения.
Сезонная линька связана с сезонными изменениями условий существования животных. У животных, не впадающих в зимнюю спячку, ежегодно наблюдается осенняя и весенняя линька.
При осенней линьке теплопроводный волосяной покров сменяется густым, тёплым мехом. При весенней линьке одновременно с заменой покрова у многих животных происходит слущивание верхней части рогового слоя эпидермиса.
Запасание кормов животными — важный инстинкт, наиболее развитый у обитателей холодных и умеренных широт с резкими сезонными изменениями кормовых условий. Наблюдается у многих беспозвоночных, у некоторых птиц и особенно часто у млекопитающих. Из беспозвоночных запасают корм некоторые пауки, крабы, раки и многие насекомые.
Из птиц корма запасают только зимующие. Большинство птиц использует запасы зимой как дополнительное питание.
Из млекопитающих запасают корм некоторые хищники, пищухи и многие грызуны. Запасы используются зимой или весной после пробуждения от спячки или зимнего сна.
Степной хорь складывает в нору сусликов, горностай — водяных крыс, мышей, лягушек, ласка — мелких грызунов. Многие пищухи заготовляют сено, складывая его в стожки или в щели между камнями. Белка запасает грибы, орехи и жёлуди. Бурундук затаскивает к себе в нору орехи, зёрна, лесная мышь — семена, речной бобр — ветви и корневища, погружая их в воду около входа в нору.
< Предыдущая страница «Борьба за существование»Примерные и окончательные причины поведения — Биология 2e
Цели обучения
К концу этого раздела вы сможете делать следующее:
- Сравнить врожденное и усвоенное поведение
- Обсудить, как движения и миграция являются результатом естественного отбора
- Обсудить различные способы общения членов населения друг с другом
- Приведите примеры того, как виды используют энергию для демонстрации спаривания и других форм ухаживания.
- Различия между различными системами спаривания
- Опишите различные способы обучения видов
Поведение — это изменение активности организма в ответ на раздражитель.Поведенческая биология — это изучение биологических и эволюционных основ таких изменений. Идея о том, что поведение возникло в результате давления естественного отбора, не нова. На протяжении десятилетий поведение животных изучали несколько ученых. Так поступают биологи в этологии; психологи в области сравнительной психологии; и другие ученые в области нейробиологии. Первые два, этология и сравнительная психология, имеют наибольшее значение для изучения поведенческой биологии.
Одна из целей поведенческой биологии — это врожденное поведение, которое имеет сильный генетический компонент и в значительной степени не зависит от влияний окружающей среды, от приобретенного поведения, которое является результатом условий окружающей среды. Врожденное поведение или инстинкт важны, потому что нет риска научиться неправильному поведению. Они «жестко подключены» к системе. С другой стороны, усвоенное поведение, хотя и более рискованное, является гибким, динамичным и может изменяться в соответствии с изменениями в окружающей среде.
Врожденное поведение: движение и миграция
Врожденное или инстинктивное поведение зависит от реакции на стимулы. Самый простой пример — рефлекторное действие, непроизвольная и быстрая реакция на раздражитель. Чтобы проверить рефлекс «коленного рефлекса», врач ударяет резиновым молотком по сухожилию надколенника ниже коленной чашечки. Стимуляция нервов приводит к рефлексу разгибания ноги в колене. Это похоже на реакцию человека, который прикасается к горячей плите и инстинктивно убирает руку.Даже люди, обладающие огромной способностью к обучению, по-прежнему демонстрируют множество врожденных форм поведения.
Кинезис и такси
Другой вид деятельности или движения врожденного поведения — кинезис, или ненаправленное движение в ответ на стимул. Ортокинез — это повышенная или пониженная скорость движения организма в ответ на раздражитель. Например, мокрицы увеличивают скорость передвижения при воздействии высоких или низких температур. Это движение, хотя и случайное, увеличивает вероятность того, что насекомое будет меньше времени проводить в неблагоприятной среде.Другой пример — клинокинез, усиление поворотов. Это проявляется бактериями, такими как E. coli , которые в сочетании с ортокинезом помогают организмам случайным образом находить более благоприятную среду.
Похожая, но более направленная версия кинезиса — это такси: направленное движение к стимулу или от него. Это движение может происходить в ответ на свет (фототаксис), химические сигналы (хемотаксис) или гравитацию (геотаксис) и может быть направлено (положительно) или от источника (отрицательно) от источника стимула.Пример положительного хемотаксиса демонстрирует одноклеточное простейшее Tetrahymena thermophila . Этот организм плавает, используя реснички, иногда двигаясь по прямой линии, а иногда делая повороты. Привлекающий хемотаксический агент изменяет частоту поворотов по мере того, как организм движется прямо к источнику, следуя возрастающему градиенту концентрации.
Шаблоны с фиксированным действием
Паттерн фиксированного действия — это серия движений, вызванных стимулом, так что даже когда стимул убран, паттерн продолжается до завершения.Пример такого поведения наблюдается у трехиглой колюшки, небольшой пресноводной рыбы ((рисунок)). Самцы этого вида развивают красный живот во время сезона размножения и в это время проявляют инстинктивную агрессивность по отношению к другим самцам. В лабораторных экспериментах исследователи подвергали таких рыб воздействию предметов, которые по форме никак не напоминали рыбу, но были окрашены в красный цвет на нижней половине. Самцы колюшки агрессивно реагировали на предметы, как если бы они были настоящими самцами колюшки.
Самцы трехиглой колюшки демонстрируют фиксированный рисунок движения. Во время брачного сезона самцы, у которых появляется ярко-красный живот, сильно реагируют на предметы с красным дном, никак не похожие на рыб.
Миграция
Миграция — это сезонное перемещение животных на большие расстояния. Это развитая, адаптированная реакция на изменение доступности ресурсов, и это обычное явление, обнаруживаемое у всех основных групп животных. Птицы летят на юг на зиму, чтобы попасть в более теплый климат с достаточным количеством пищи, а лосось мигрирует к своим нерестилищам.В популярном документальном фильме 2005 года «Марш пингвинов « »прослеживается 62-мильная миграция императорских пингвинов через Антарктиду, чтобы принести пищу к месту их размножения и к их детенышам. Гну ((Рисунок)) ежегодно мигрируют на расстояние более 1800 миль в поисках новых лугов.
Гну ежегодно мигрируют по часовой стрелке на расстояние более 1800 миль в поисках созревшей дождем травы. (кредит: Эрик Инафуку)
Хотя миграция считается врожденным поведением, только некоторые мигрирующие виды всегда мигрируют (облигатная миграция).Животные, которые демонстрируют факультативную миграцию, могут выбирать мигрировать или нет. Кроме того, у некоторых животных мигрирует только часть популяции, тогда как остальная часть не мигрирует (неполная миграция). Например, совы, обитающие в тундре, могут мигрировать в годы, когда их источник пищи, мелкие грызуны, относительно мало, но не мигрировать в годы, когда грызунов много.
Собирательство
Собирательство — это поиск и использование пищевых ресурсов. Кормление, которое максимизирует прирост энергии и сводит к минимуму расход энергии, называется оптимальным поведением при кормлении, и им отдается предпочтение естественным путем.Раскрашенный аист, например, использует свой длинный клюв, чтобы искать на дне пресноводных болот крабов и другую пищу ((Рисунок)).
Раскрашенный аист добывает корм своим длинным клювом. (кредит: J.M. Garg)
Врожденное поведение: жизнь в группах
Не все животные живут группами, но даже те, которые живут относительно уединенно, за исключением тех, которые могут размножаться бесполым путем, должны спариваться. Спаривание обычно заключается в том, что одно животное сигнализирует другому, чтобы сообщить о своем желании спариться.Есть несколько типов энергоемкого поведения или проявлений, связанных со спариванием, которые называются брачными ритуалами. Другое поведение, обнаруживаемое в популяциях, которые живут в группах, описано с точки зрения того, какое животное получает выгоду от поведения. В эгоистичном поведении выигрывает только рассматриваемое животное; при альтруистическом поведении действия одного животного приносят пользу другому; Кооперативное поведение описывает, когда оба животного получают выгоду. Все эти формы поведения предполагают своего рода общение между членами населения.
Общение внутри вида
Животные общаются друг с другом с помощью стимулов, известных как сигналы. Примером этого является трехиглая колюшка, где визуальный сигнал красной области в нижней половине рыбы сигнализирует самцам о необходимости стать агрессивными, а самкам — к спариванию. Другие сигналы — химические (феромоны), слуховые (звуковые), визуальные (ухаживания и агрессивные проявления) или тактильные (прикосновения). Эти типы общения могут быть инстинктивными, усвоенными или сочетанием того и другого.Это не то же самое, что общение, которое мы связываем с языком, которое наблюдается только у людей и, возможно, у некоторых видов приматов и китообразных.
Феромон — это секретируемый химический сигнал, используемый для получения ответа от другого человека того же вида. Цель феромонов — вызвать определенное поведение у принимающего человека. Феромоны особенно распространены среди социальных насекомых, но они используются многими видами для привлечения противоположного пола, для подачи сигналов тревоги, для обозначения следов еды и для вызова других, более сложных форм поведения.Считается, что даже люди реагируют на определенные феромоны, называемые подмышечными стероидами. Эти химические вещества влияют на человеческое восприятие других людей, и в одном исследовании они отвечали за синхронизацию менструальных циклов у группы женщин. Роль феромонов в общении между людьми до конца не изучена и продолжает изучаться.
Песни — это пример звукового сигнала, который должен быть услышан получателем. Возможно, самые известные из них — это песни птиц, которые определяют вид и используются для привлечения самок.Другие хорошо известные песни — это песни китов, которые настолько редки, что могут преодолевать большие расстояния под водой. Виды дельфинов общаются друг с другом (а иногда и с другими видами дельфинов), используя самые разные вокализации. Сверчки-самцы издают щебечущие звуки, используя специальный орган, чтобы привлечь партнера, оттолкнуть других самцов и объявить об успешном спаривании.
Демонстрация ухаживания — это серия ритуализированных визуальных форм поведения (сигналов), предназначенных для привлечения и убеждения представителя противоположного пола к спариванию.Эти показы повсеместны в животном мире. Часто эти отображения включают в себя серию шагов, включая начальное отображение одного члена, за которым следует ответ от другого. Если в какой-то момент изображение будет отображаться неправильно или не будет получен надлежащий ответ, брачный ритуал будет отменен и попытка брачного союза будет неудачной. Брачный дисплей обыкновенного аиста показан на (Рисунок).
Агрессивные проявления также обычны в животном мире. Например, собака обнажает зубы, когда хочет, чтобы другая собака отступила.Предположительно, эти проявления передают не только готовность животного к драке, но и его боевые способности. Хотя эти дисплеи действительно сигнализируют об агрессии со стороны отправителя, считается, что эти дисплеи на самом деле являются механизмом для уменьшения количества реальных боев, которые происходят между представителями одного и того же вида: они позволяют людям оценить боевые способности своего противника. и, таким образом, решить, «стоит ли оно того, чтобы бороться». Проверка некоторых гипотез с использованием теории игр привела к выводу, что некоторые из этих отображений могут преувеличивать фактические боевые способности животного и используются для «блефа» оппонента.Этот тип взаимодействия, даже если он «нечестный», будет одобрен естественным отбором, если он будет успешным в большинстве случаев.
Выставка ухаживания этого аиста предназначена для привлечения потенциальных партнеров. (Источник: Линда «jinterwas» / Flickr)
Отвлечение внимания наблюдается у птиц и некоторых рыб. Они созданы для того, чтобы отвести хищника от гнезда. Это пример альтруистического поведения: оно приносит пользу молодежи больше, чем человеку, выполняющему показ, который тем самым подвергает себя риску.
Многие животные, особенно приматы, общаются с другими членами группы посредством прикосновений. Такие действия, как уход, прикосновение к плечу или корню хвоста, объятия, контакт губ и церемонии приветствия наблюдались у индийских лангуров, обезьян Старого Света. Подобное поведение наблюдается и у других приматов, особенно у человекообразных обезьян.
Ссылка на обучение
Птица-убийца отвлекает хищников от своих яиц, изображая сломанное крыло на этом видео, сделанном в Бойсе, штат Айдахо.
Альтруистическое поведение
Поведение, которое снижает физическую подготовку человека, но увеличивает физическую форму другого человека, называется альтруистическим. Примеры такого поведения широко распространены в животном мире. Социальные насекомые, такие как рабочие пчелы, не способны к размножению, но они поддерживают королеву, чтобы она могла заселить улей своим потомством. Сурикаты держат часового на страже, чтобы предупредить остальную часть колонии о вторжении, даже если часовой подвергает себя опасности.Волки и дикие собаки приносят мясо членам стаи, отсутствующим на охоте. Лемуры заботятся о младенцах, не связанных с ними. Хотя на первый взгляд такое поведение кажется альтруистическим, на самом деле все не так просто.
Было много дискуссий о том, почему существует альтруистическое поведение. Приводит ли такое поведение к общим эволюционным преимуществам для их вида? Помогают ли они альтруисту передать собственные гены? А как насчет такой деятельности между не связанными между собой людьми? Одно объяснение поведения альтруистического типа можно найти в генетике естественного отбора.В книге « Эгоистичный ген» 1976 года ученый Ричард Докинз попытался объяснить многие, казалось бы, альтруистические формы поведения с точки зрения самого гена. Хотя ген, очевидно, не может быть эгоистичным в человеческом смысле, это может выглядеть так, если жертва индивида приносит пользу людям, имеющим общие гены, идентичные по происхождению (присутствующие у родственников из-за общего происхождения). Родители млекопитающих приносят эту жертву, чтобы заботиться о своем потомстве. Императорские пингвины мигрируют на многие мили в суровых условиях, чтобы принести пищу своим детенышам.Теория эгоистичного гена вызвала споры на протяжении многих лет и до сих пор обсуждается учеными в смежных областях.
Даже люди с меньшим родством, с меньшей генетической идентичностью, чем у родителей и потомков, извлекают выгоду из, казалось бы, альтруистического поведения. Действия социальных насекомых, таких как пчелы, осы, муравьи и термиты, являются хорошими примерами. Бесплодные рабочие в этих обществах заботятся о королеве, потому что они тесно связаны с ней, и поскольку королева имеет потомство, она косвенно передает гены от рабочих.Таким образом, для рабочего выгодно содержать матку, не имея прямого шанса передать ее гены из-за ее бесплодия. Понижение индивидуальной приспособленности для повышения репродуктивной приспособленности родственника и, следовательно, индивидуальной приспособленности происходит через родственный отбор. Этот феномен может объяснить многие на первый взгляд виды альтруистического поведения, наблюдаемые у животных. Однако такое поведение нельзя по-настоящему определить как альтруизм в этих случаях, потому что субъект на самом деле увеличивает свою приспособленность либо напрямую (через собственное потомство), либо косвенно (через инклюзивную приспособленность, которую он приобретает через родственников, которые имеют общие с ним гены).
Неродственные люди также могут действовать альтруистично друг другу, и это, кажется, бросает вызов объяснению «эгоистичного гена». Пример этого наблюдается у многих видов обезьян, когда обезьяна показывает свою спину обезьяне, не являющейся родственницей, чтобы та выбрала паразитов из ее шерсти. По прошествии определенного времени роли меняются, и первая обезьяна ухаживает за второй обезьяной. Таким образом, в поведении присутствует взаимность. Оба получают выгоду от взаимодействия, и их приспособленность повышается больше, чем если бы они не сотрудничали или если бы один сотрудничал, а другой не сотрудничал.Такое поведение все же не обязательно является альтруизмом, поскольку «отдающее» поведение актера основано на ожидании того, что он будет «получателем» поведения в будущем, что называется реципрокным альтруизмом. Взаимный альтруизм требует, чтобы люди неоднократно встречались друг с другом, часто в результате проживания в одной социальной группе, и чтобы мошенники (те, кто никогда не «сдавались») были наказаны.
Эволюционная теория игр, модификация классической теории игр в математике, показала, что многие из этих так называемых «альтруистических моделей поведения» вовсе не альтруистичны.Определение «чистого» альтруизма, основанное на человеческом поведении, — это действие, которое приносит пользу другому без какой-либо прямой выгоды для себя. Похоже, что большинство описанных ранее форм поведения не удовлетворяют этому определению, и теоретики игр умеют находить в них «эгоистичные» компоненты. Другие утверждали, что при обсуждении поведения животных следует полностью отказаться от терминов «эгоистичный» и «альтруистический», поскольку они описывают поведение человека и могут не иметь прямого отношения к инстинктивной активности животных.Ясно, однако, что наследственное поведение, которое увеличивает шансы передачи своих генов или части генов, поддерживается естественным отбором и будет сохраняться в будущих поколениях до тех пор, пока это поведение дает преимущество в приспособленности. Это инстинктивное поведение затем может быть применено, в особых обстоятельствах, к другим видам, если оно не снижает приспособленность животного.
В поисках сексуальных партнеров
Не все животные размножаются половым путем, но многие из них сталкиваются с одной и той же проблемой: им нужно найти подходящую пару и часто приходится соревноваться с другими особями, чтобы получить ее.Значительная энергия тратится на поиск, привлечение и совокупление полового партнера. В ходе этого процесса происходят два типа отбора: интерсексуальный отбор, когда особи одного пола выбирают себе партнеров другого пола, и внутриполовой отбор, конкуренция за спаривание между представителями одного пола. Интерсексуальный отбор часто бывает сложным, потому что выбор партнера может основываться на множестве визуальных, слуховых, тактильных и химических сигналов. Пример интерсексуального отбора — это когда самка павлина выбирает спариваться с самцом с самым ярким оперением.Этот тип отбора часто приводит к чертам выбранного пола, которые не повышают выживаемость, но являются теми чертами, которые наиболее привлекательны для противоположного пола (часто за счет выживания). Внутрисексуальный отбор включает в себя брачные демонстрации и агрессивные брачные ритуалы, такие как бода баранов головами — в этих битвах побеждает тот, кто способен спариваться. Многие из этих ритуалов отнимают много энергии, но приводят к выбору самых здоровых, сильных и / или доминирующих особей для спаривания.
В популяциях животных наблюдаются три основные системы спаривания, все из которых связаны с врожденным поведением, а не с усвоенным: моногамным, полигамным и полиандрическим.
Ссылка на обучение
Посетите этот веб-сайт, чтобы увидеть информативные видеоролики о сексуальном выборе.
В моногамных системах один самец и одна самка спариваются по крайней мере на один сезон размножения. У некоторых животных, таких как серый волк, эти ассоциации могут длиться намного дольше, даже на всю жизнь. Несколько теорий могут объяснить этот тип брачной системы.«Гипотеза защиты партнера» утверждает, что самцы остаются с самкой, чтобы не дать другим самцам спариваться с ней. Такое поведение выгодно в таких ситуациях, когда помощников мало и их трудно найти. Другое объяснение — это «гипотеза помощи самцам», согласно которой самцы, которые помогают охранять и воспитывать своих детенышей, будут иметь более здоровое потомство. Моногамия наблюдается во многих популяциях птиц, где, помимо родительской заботы со стороны самки, самец также является основным источником родительской заботы о птенцах.Третье объяснение эволюционных преимуществ моногамии — это «гипотеза женского принуждения». В этом сценарии самка гарантирует, что у самца не будет другого потомства, которое могло бы конкурировать с ее собственным, поэтому она активно вмешивается в сигналы самца, чтобы привлечь других самцов.
Полигинное спаривание означает спаривание одного самца с несколькими самками. В этих ситуациях самка должна нести большую часть родительской заботы, поскольку одинокий самец не способен заботиться о таком большом количестве потомков.В полигинии, основанной на ресурсах, самцы соревнуются за территории с лучшими ресурсами, а затем спариваются с самками, которые проникают на территорию, привлеченные ее богатством ресурсов. Самка получает выгоду от спаривания с доминантным, генетически подходящим самцом; однако это происходит за счет отсутствия помощи мужчин в уходе за потомством. Пример можно увидеть в медогиде с желтым крестцом, птице, самцы которой защищают ульи, потому что самки питаются их воском. По мере приближения самок самец, защищающий гнездо, спаривается с ними.Брачные структуры гаремов — это тип полигинной системы, в которой определенные самцы доминируют при спаривании, контролируя территорию с ресурсами. Спаривание гаремов происходит у морских слонов, где альфа-самец доминирует при спаривании внутри группы. Третий тип многоженства — это система лек. Здесь есть общая площадка для ухаживаний, где несколько самцов демонстрируют замысловатые представления для самок, а самки выбирают себе пару из этой группы. Такое поведение наблюдается у нескольких видов птиц, включая шалфейный тетерев и степной цыпленок.
В полиандрических системах спаривания одна самка спаривается со многими самцами. Эти типы систем гораздо реже, чем моногамные и полигамные брачные системы. У морских игл и морских коньков самцы получают яйца от самок, оплодотворяют их, защищают в сумке и дают потомство ((Рисунок)). Таким образом, самка может дать яйца нескольким самцам без бремени вынашивания оплодотворенных яиц.
Спаривание с полиандроусом, при котором одна самка спаривается со многими самцами, происходит у морского конька (а) и рыбы-иглы (b).(кредит А: модификация работы Брайана Грэтвика; кредит В: модификация работы Стивена Чайлдса)
Простое выученное поведение
Большинство описанных ранее форм поведения были врожденными или, по крайней мере, имели врожденный компонент (можно изучить варианты врожденного поведения). Они передаются по наследству, и поведение не меняется в ответ на сигналы из окружающей среды. И наоборот, усвоенное поведение, даже если оно может иметь инстинктивные компоненты, позволяет организму адаптироваться к изменениям в окружающей среде и модифицируется предыдущим опытом.Простые усвоенные формы поведения включают в себя привыкание и импринтинг — оба они важны для процесса созревания молодых животных.
Привыкание
Привыкание — это простая форма обучения, при которой животное перестает реагировать на раздражитель после периода многократного воздействия. Это форма неассоциативного обучения, поскольку стимул не связан ни с каким наказанием или вознаграждением. Луговые собачки обычно издают тревожный сигнал, когда им угрожает хищник, но они привыкают к звуку человеческих шагов, когда с этим звуком не связано никакого вреда, поэтому они больше не реагируют на них тревожным звонком.В этом примере привыкание относится к звукам шагов человека, поскольку животные по-прежнему реагируют на звуки потенциальных хищников.
Печать
Импринтинг — это тип обучения, который происходит в определенном возрасте или на определенном этапе жизни, который является быстрым и независимым от участвующих видов. Птенцы утки узнают первого взрослого, которого они увидят, свою мать, и завязывают с ней связь. Привычное зрелище — утята, идущие или плывущие вслед за матерью ((рисунок)). Это еще один тип неассоциативного обучения, но он очень важен в процессе созревания этих животных, поскольку побуждает их оставаться рядом с матерью, чтобы они были защищены, что значительно увеличивает их шансы на выживание.Однако, если новорожденные утки увидят человека до того, как увидят свою мать, они запечатлеваются на человеке и будут следовать за ним точно так же, как они следовали бы за своей настоящей матерью.
Привязанность утят к матери — пример импринтинга. (кредит: модификация работы Марка Харкина)
Ссылка на обучение
Международный фонд журавлей помог увеличить мировую популяцию американских журавлей с 21 особи до примерно 600.Отпечаток вылупившихся птенцов был ключом к успеху: биологи носят полные костюмы журавля, чтобы птицы никогда не «видели» людей. Посмотрите это видео, чтобы узнать больше.
Условное поведение
Условное поведение — это типы ассоциативного обучения, при котором стимул ассоциируется со следствием. Во время оперантного обусловливания поведенческая реакция модифицируется ее последствиями, касающимися ее формы, силы или частоты.
Классический кондиционер
В классическом кондиционировании реакция, называемая условной реакцией, связана со стимулом, с которым она ранее не была связана, — условным раздражителем.Реакция на исходный безусловный раздражитель называется безусловной реакцией. Наиболее цитируемым примером классической обусловленности являются эксперименты Ивана Павлова с собаками ((Рисунок)). В экспериментах Павлова безусловной реакцией было выделение у собак слюноотделения в ответ на безусловный стимул видеть или чувствовать запах их пищи. Обусловливающим стимулом, который исследователи связали с безусловной реакцией, был звонок в колокольчик. Во время кондиционирования каждый раз, когда животному давали пищу, звонили в звонок.Это повторилось во время нескольких испытаний. Через некоторое время собака научилась ассоциировать звон колокольчика с едой и реагировать выделением слюны. По окончании периода кондиционирования собака реагировала выделением слюны на звонок, даже когда безусловный стимул, еда, отсутствовала. Таким образом, звон колокольчика стал условным раздражителем, а слюноотделение — условной реакцией. Хотя некоторые ученые считают, что безусловная и условная реакции идентичны, даже Павлов обнаружил, что слюна у условных собак имеет характерные отличия по сравнению с безусловной собакой.
В классическом павловском ответе собака начинает ассоциировать звонок в колокольчик с едой.
Некоторые ученые полагали, что для этого типа кондиционирования требуется многократное воздействие парных стимулов и ответов, но теперь известно, что это не обязательно во всех случаях, и что определенному условию можно научиться в одном парном эксперименте. Классическая обусловленность — главный принцип бихевиоризма, ветви психологической философии, которая предполагает, что все действия, мысли и эмоции живых существ являются поведением, которое можно лечить путем модификации поведения и изменений в окружающей среде.
Оперативное кондиционирование
При оперантном обусловливании условное поведение постепенно модифицируется его последствиями по мере того, как животное реагирует на раздражитель. Основным сторонником такой обусловленности был психолог Б.Ф. Скиннер, изобретатель ящика Скиннера. Скиннер поместил крыс в свои ящики, в которых был рычаг, который выдавал крысе пищу при нажатии. Хотя изначально крыса несколько раз нажимала на рычаг случайно, в конечном итоге нажатие на рычаг ассоциировалось с получением еды.Этот тип обучения является примером оперантного обусловливания. Оперантное обучение — основа большинства дрессировок животных. Обусловленное поведение постоянно изменяется с помощью положительного или отрицательного подкрепления, часто в виде награды, такой как еда или какой-либо вид наказания, соответственно. Таким образом, животное приучают ассоциировать определенный тип поведения с наказанием или наградой, и со временем его можно побудить к поведению, которое они не стали бы делать в дикой природе, например, к «трюкам» дельфинов. показывает морской парк аттракционов ((Рисунок)).
Дрессировка дельфинов путем вознаграждения их пищей является примером положительного подкрепления оперантной обусловленности. (кредит: Роланд Танглао)
Когнитивное обучение
Классическая и оперантная обусловленность — неэффективные способы обучения людей и других разумных животных. Некоторые приматы, в том числе люди, могут учиться, подражая поведению других и следуя инструкциям. Развитие сложного языка людьми сделало когнитивное обучение, манипулирование информацией с помощью разума, наиболее известным методом человеческого обучения.Фактически, именно так студенты учатся прямо сейчас, читая эту книгу. По мере чтения учащиеся могут создавать мысленные образы объектов или организмов, воображать изменения в них или их поведение и предвидеть последствия. Помимо визуальной обработки, когнитивное обучение также улучшается за счет запоминания прошлого опыта, прикосновения к физическим объектам, слушания звуков, дегустации еды и множества других сенсорных входов. Когнитивное обучение настолько мощно, что его можно использовать для детального понимания обусловленности.В обратном сценарии кондиционирование не может помочь кому-то узнать о познании.
Классическая работа по когнитивному обучению была проделана Вольфгангом Кёлером с шимпанзе. Он продемонстрировал способность этих животных к абстрактному мышлению, показав, что они могут научиться решать головоломки. Когда банан был подвешен в их клетке слишком высоко, чтобы они могли достать до них, и несколько ящиков были случайным образом расставлены на полу, некоторые шимпанзе могли складывать ящики друг на друга, взбираться на них и подниматься. банан.Это означает, что они могли визуализировать результат складывания ящиков еще до того, как они выполнили действие. Этот тип обучения гораздо более мощный и универсальный, чем кондиционирование.
Когнитивное обучение не ограничивается приматами, хотя они наиболее эффективно его используют. Эксперименты по бегу в лабиринте, проведенные на крысах Х. Блоджетт в 1920-х годах был первым, кто продемонстрировал когнитивные способности простого млекопитающего. Мотивом для животных проложить себе путь через лабиринт был кусок еды на его конце.В этих исследованиях животных в группе I запускали по одной пробе в день, и по завершении пробежки они получали пищу каждый день ((рисунок)). Крыс группы II не кормили в лабиринте в течение первых шести дней, а затем проводили последующие прогоны с пищей в течение нескольких дней. Крысы группы III получали пищу на третий день и каждый последующий день. В результате контрольные крысы, группа I, быстро научились и поняли, как пройти лабиринт за семь дней. Группа III не многому научилась в течение трех дней без еды, но быстро догнала контрольную группу, когда ей дали пищевое вознаграждение.Группа II училась очень медленно в течение шести дней без вознаграждения, чтобы мотивировать их, и они не начали догонять контрольную группу до тех пор, пока не был получен дневной корм, а затем потребовалось на два дня больше, чтобы выучить лабиринт.
Группа I (сплошная зеленая линия) находила пищу в конце каждого испытания, группа II (синяя пунктирная линия) не находила пищу в течение первых 6 дней, а группа III (красная пунктирная линия) не находила пищу в течение работает в первые три дня. Обратите внимание, что крысы, которым давали корм раньше, учились быстрее и в конечном итоге догнали контрольную группу.Оранжевые точки на линиях групп II и III показывают дни, когда в лабиринты добавлялись пищевые награды.
Может быть не сразу очевидно, что этот тип обучения отличается от кондиционирования. Хотя у кого-то может возникнуть соблазн поверить в то, что крысы просто научились находить путь через условную серию поворотов вправо и влево, Е.К. Толмен через десять лет доказал, что крысы в своем сознании создавали представление о лабиринте, что он назвал «познавательная карта.«Это была ранняя демонстрация силы когнитивного обучения и того, что эти способности не ограничиваются только людьми.
Непосредственные и окончательные причины поведения — Биология
OpenStaxCollege
[latexpage]
Цели обучения
К концу этого раздела вы сможете:
- Сравнить врожденное и усвоенное поведение
- Обсудить, как движения и миграция являются результатом естественного отбора
- Обсудить различные способы общения членов населения друг с другом
- Приведите примеры того, как виды используют энергию для демонстрации спаривания и других форм ухаживания.
- Различия между различными системами спаривания
- Опишите различные способы обучения видов
Поведение — это изменение активности организма в ответ на раздражитель.Поведенческая биология — это изучение биологических и эволюционных основ таких изменений. Идея о том, что поведение возникло в результате давления естественного отбора, не нова. Поведение животных десятилетиями изучается биологами в области этологии, психологами в области сравнительной психологии и учеными многих дисциплин, занимающихся нейробиологией. Хотя эти дисциплины частично совпадают, ученые в этих областях поведения используют разные подходы.Сравнительная психология — это продолжение работы, проделанной в области психологии человека и поведенческой психологии. Этология — это расширение генетики, эволюции, анатомии, физиологии и других биологических дисциплин. Тем не менее, нельзя изучать поведенческую биологию, не затрагивая как сравнительную психологию, так и этологию.
Одна из целей поведенческой биологии — отделить врожденное поведение, которое имеет сильный генетический компонент и в значительной степени не зависит от влияний окружающей среды, от усвоенного поведения, которое является результатом условий окружающей среды.Врожденное поведение или инстинкт важны, потому что нет риска научиться неправильному поведению. Они «жестко подключены» к системе. С другой стороны, усвоенное поведение, хотя и более рискованное, является гибким, динамичным и может изменяться в соответствии с изменениями в окружающей среде.
Врожденное или инстинктивное поведение зависит от реакции на стимулы. Самый простой пример — рефлекторное действие, непроизвольная и быстрая реакция на раздражитель. Чтобы проверить рефлекс «коленного рефлекса», врач ударяет резиновым молотком по сухожилию надколенника ниже коленной чашечки.Стимуляция нервов приводит к рефлексу разгибания ноги в колене. Это похоже на реакцию человека, который прикасается к горячей плите и инстинктивно убирает руку. Даже люди, обладающие огромной способностью к обучению, по-прежнему демонстрируют множество врожденных форм поведения.
Кинезис и такси
Другой вид деятельности или движения врожденного поведения — кинезис, или ненаправленное движение в ответ на стимул. Ортокинез — это повышенная или пониженная скорость движения организма в ответ на раздражитель.Например, мокрицы увеличивают скорость передвижения при воздействии высоких или низких температур. Это движение, хотя и случайное, увеличивает вероятность того, что насекомое будет меньше времени проводить в неблагоприятной среде. Другой пример — клинокинез, усиление поворотов. Это проявляется бактериями, такими как E. coli , которые в сочетании с ортокинезом помогают организмам случайным образом находить более благоприятную среду.
Похожая, но более направленная версия кинезиса — это такси: направленное движение к стимулу или от него.Это движение может происходить в ответ на свет (фототаксис), химические сигналы (хемотаксис) или гравитацию (геотаксис) и может быть направлено (положительно) или от источника (отрицательно) от источника стимула. Пример положительного хемотаксиса демонстрирует одноклеточное простейшее Tetrahymena thermophila . Этот организм плавает, используя реснички, иногда двигаясь по прямой линии, а иногда делая повороты. Привлекающий хемотаксический агент изменяет частоту поворотов по мере того, как организм движется прямо к источнику, следуя возрастающему градиенту концентрации.
Шаблоны с фиксированным действием
Паттерн фиксированного действия — это серия движений, вызванных стимулом, так что даже когда стимул убран, паттерн продолжается до завершения. Пример такого поведения наблюдается у трехиглой колюшки, небольшой пресноводной рыбы ([ссылка]). Самцы этого вида развивают красный живот во время сезона размножения и в это время проявляют инстинктивную агрессивность по отношению к другим самцам. В лабораторных экспериментах исследователи подвергали таких рыб воздействию предметов, которые по форме никак не напоминали рыбу, но были окрашены в красный цвет на нижней половине.Самцы колюшки агрессивно реагировали на предметы, как если бы они были настоящими самцами колюшки.
Самцы трехиглой колюшки демонстрируют фиксированный рисунок движения. Во время брачного сезона самцы, у которых появляется ярко-красный живот, сильно реагируют на предметы с красным дном, никак не похожие на рыб.
Миграция
Миграция — это сезонное перемещение животных на большие расстояния. Это развитая, адаптированная реакция на изменение доступности ресурсов, и это обычное явление, обнаруживаемое у всех основных групп животных.Птицы летят на юг на зиму, чтобы попасть в более теплый климат с достаточным количеством пищи, а лосось мигрирует к своим нерестилищам. В популярном документальном фильме 2005 года «Марш пингвинов « »прослеживается 62-мильная миграция императорских пингвинов через Антарктиду, чтобы принести пищу к месту их размножения и к их детенышам. Гну ([ссылка]) ежегодно мигрируют более чем на 1800 миль в поисках новых лугов.
Гну ежегодно мигрируют по часовой стрелке на расстояние более 1800 миль в поисках созревшей дождем травы.(кредит: Эрик Инафуку)
Хотя миграция считается врожденным поведением, только некоторые мигрирующие виды всегда мигрируют (облигатная миграция). Животные, которые демонстрируют факультативную миграцию, могут выбирать мигрировать или нет. Кроме того, у некоторых животных мигрирует только часть популяции, тогда как остальная часть не мигрирует (неполная миграция). Например, совы, обитающие в тундре, могут мигрировать в годы, когда их источник пищи, мелкие грызуны, относительно мало, но не мигрировать в годы, когда грызунов много.
Собирательство
Собирательство — это поиск и использование пищевых ресурсов. Кормление, которое максимизирует прирост энергии и сводит к минимуму расход энергии, называется оптимальным поведением при кормлении, и им отдается предпочтение естественным путем. Раскрашенный аист, например, использует свой длинный клюв, чтобы искать на дне пресноводных болот крабов и другую пищу ([ссылка]).
Раскрашенный аист добывает корм своим длинным клювом. (кредит: J.M. Garg)
Не все животные живут группами, но даже те, которые живут относительно уединенно, за исключением тех, которые могут размножаться бесполым путем, должны спариваться.Спаривание обычно заключается в том, что одно животное сигнализирует другому, чтобы сообщить о своем желании спариться. Есть несколько типов энергоемкого поведения или проявлений, связанных со спариванием, которые называются брачными ритуалами. Другое поведение, обнаруживаемое в популяциях, которые живут в группах, описано с точки зрения того, какое животное получает выгоду от поведения. В эгоистичном поведении выигрывает только рассматриваемое животное; при альтруистическом поведении действия одного животного приносят пользу другому; Кооперативное поведение описывает, когда оба животного получают выгоду.Все эти формы поведения предполагают своего рода общение между членами населения.
Общение внутри вида
Животные общаются друг с другом с помощью стимулов, известных как сигналы. Примером этого является трехиглая колюшка, где визуальный сигнал красной области в нижней половине рыбы сигнализирует самцам о необходимости стать агрессивными, а самкам — к спариванию. Другие сигналы — химические (феромоны), слуховые (звуковые), визуальные (ухаживания и агрессивные проявления) или тактильные (прикосновения).Эти типы общения могут быть инстинктивными, усвоенными или сочетанием того и другого. Это не то же самое, что общение, которое мы связываем с языком, которое наблюдается только у людей и, возможно, у некоторых видов приматов и китообразных.
Феромон — это секретируемый химический сигнал, используемый для получения ответа от другого человека того же вида. Цель феромонов — вызвать определенное поведение у принимающего человека. Феромоны особенно распространены среди социальных насекомых, но они используются многими видами для привлечения противоположного пола, для подачи сигналов тревоги, для обозначения следов еды и для вызова других, более сложных форм поведения.Считается, что даже люди реагируют на определенные феромоны, называемые подмышечными стероидами. Эти химические вещества влияют на человеческое восприятие других людей, и в одном исследовании они отвечали за синхронизацию менструальных циклов у группы женщин. Роль феромонов в общении между людьми все еще остается спорной и продолжает изучаться.
Песни — это пример звукового сигнала, который должен быть услышан получателем. Возможно, самые известные из них — это песни птиц, которые определяют вид и используются для привлечения самок.Другие хорошо известные песни — это песни китов, которые настолько редки, что могут преодолевать большие расстояния под водой. Дельфины общаются друг с другом, используя самые разные вокализации. Сверчки-самцы издают щебечущие звуки, используя специальный орган, чтобы привлечь партнера, оттолкнуть других самцов и объявить об успешном спаривании.
Демонстрация ухаживания — это серия ритуализированных визуальных форм поведения (сигналов), предназначенных для привлечения и убеждения представителя противоположного пола к спариванию. Эти показы повсеместны в животном мире.Часто эти отображения включают в себя серию шагов, включая начальное отображение одного члена, за которым следует ответ от другого. Если в какой-то момент изображение будет отображаться неправильно или не будет получен надлежащий ответ, брачный ритуал будет отменен и попытка брачного союза будет неудачной. Брачный дисплей обыкновенного аиста показан в [ссылка].
Агрессивные проявления также обычны в животном мире. Примером может служить собака, обнажающая зубы, когда она хочет, чтобы другая собака отступила.Предположительно, эти проявления передают не только готовность животного к драке, но и его боевые способности. Хотя эти дисплеи действительно сигнализируют об агрессии со стороны отправителя, считается, что эти дисплеи на самом деле являются механизмом для уменьшения количества реальных боев, которые происходят между представителями одного и того же вида: они позволяют людям оценить боевые способности своего противника. и, таким образом, решить, «стоит ли оно того, чтобы бороться». Проверка некоторых гипотез с использованием теории игр привела к выводу, что некоторые из этих отображений могут преувеличивать фактические боевые способности животного и используются для «блефа» оппонента.Этот тип взаимодействия, даже если он «нечестный», будет одобрен естественным отбором, если он будет успешным в большинстве случаев.
Выставка ухаживания этого аиста предназначена для привлечения потенциальных партнеров. (Источник: Линда «jinterwas» / Flickr)
Отвлечение внимания наблюдается у птиц и некоторых рыб. Они созданы для того, чтобы отвести хищника от гнезда, в котором находится их детеныш. Это пример альтруистического поведения: оно приносит пользу молодежи больше, чем человеку, выполняющему показ, который тем самым подвергает себя риску.
Многие животные, особенно приматы, общаются с другими членами группы посредством прикосновений. Такие действия, как уход, прикосновение к плечу или корню хвоста, объятия, контакт губ и церемонии приветствия наблюдались у индийских лангуров, обезьян Старого Света. Подобное поведение наблюдается и у других приматов, особенно у человекообразных обезьян.
Ссылка на обучение
Птица-убийца отвлекает хищников от своих яиц, изображая сломанное крыло на этом видео, сделанном в Бойсе, штат Айдахо.
Альтруистическое поведение
Поведение, которое снижает физическую подготовку человека, но увеличивает физическую форму другого человека, называется альтруистическим. Примеры такого поведения широко распространены в животном мире. Социальные насекомые, такие как рабочие пчелы, не способны к размножению, но они поддерживают королеву, чтобы она могла заселить улей своим потомством. Сурикаты держат часового на страже, чтобы предупредить остальную часть колонии о вторжении, даже если часовой подвергает себя опасности.Волки и дикие собаки приносят мясо членам стаи, отсутствующим на охоте. Лемуры заботятся о младенцах, не связанных с ними. Хотя на первый взгляд такое поведение кажется альтруистическим, все может быть не так просто.
Было много дискуссий о том, почему существует альтруистическое поведение. Приводит ли такое поведение к общим эволюционным преимуществам для их вида? Помогают ли они альтруисту передать собственные гены? А как насчет такой деятельности между не связанными между собой людьми? Одно объяснение поведения альтруистического типа можно найти в генетике естественного отбора.В книге « Эгоистичный ген» 1976 года ученый Ричард Докинз попытался объяснить многие, казалось бы, альтруистические формы поведения с точки зрения самого гена. Хотя ген, очевидно, не может быть эгоистичным в человеческом смысле, это может выглядеть так, если жертва индивида приносит пользу людям, имеющим общие гены, идентичные по происхождению (присутствующие у родственников из-за общего происхождения). Родители млекопитающих приносят эту жертву, чтобы заботиться о своем потомстве. Императорские пингвины мигрируют на многие мили в суровых условиях, чтобы принести пищу своим детенышам.Теория эгоистичного гена вызвала споры на протяжении многих лет и до сих пор обсуждается учеными в смежных областях.
Даже люди с меньшим родством, с меньшей генетической идентичностью, чем у родителей и потомков, извлекают выгоду из, казалось бы, альтруистического поведения. Действия социальных насекомых, таких как пчелы, осы, муравьи и термиты, являются хорошими примерами. Бесплодные рабочие в этих обществах заботятся о королеве, потому что они тесно связаны с ней, и поскольку королева имеет потомство, она косвенно передает гены от рабочих.Таким образом, для рабочего выгодно содержать матку, не имея прямого шанса передать ее гены из-за ее бесплодия. Понижение индивидуальной приспособленности для повышения репродуктивной приспособленности родственника и, следовательно, индивидуальной приспособленности происходит через родственный отбор. Этот феномен может объяснить многие на первый взгляд виды альтруистического поведения, наблюдаемые у животных. Однако такое поведение нельзя по-настоящему определить как альтруизм в этих случаях, потому что субъект на самом деле увеличивает свою приспособленность либо напрямую (через собственное потомство), либо косвенно (через инклюзивную приспособленность, которую он приобретает через родственников, которые имеют общие с ним гены).
Неродственные люди также могут действовать альтруистично друг другу, и это, кажется, бросает вызов объяснению «эгоистичного гена». Пример этого наблюдается у многих видов обезьян, когда обезьяна показывает свою спину обезьяне, не являющейся родственницей, чтобы та выбрала паразитов из ее шерсти. По прошествии определенного времени роли меняются, и первая обезьяна ухаживает за второй обезьяной. Таким образом, в поведении присутствует взаимность. Оба получают выгоду от взаимодействия, и их приспособленность повышается больше, чем если бы они не сотрудничали или если бы один сотрудничал, а другой не сотрудничал.Такое поведение все же не обязательно является альтруизмом, поскольку «отдающее» поведение актера основано на ожидании того, что он будет «получателем» поведения в будущем, что называется реципрокным альтруизмом. Взаимный альтруизм требует, чтобы люди неоднократно встречались друг с другом, часто в результате проживания в одной социальной группе, и чтобы мошенники (те, кто никогда не «сдавались») были наказаны.
Эволюционная теория игр, модификация классической теории игр в математике, показала, что многие из этих так называемых «альтруистических моделей поведения» вовсе не альтруистичны.Определение «чистого» альтруизма, основанное на человеческом поведении, — это действие, которое приносит пользу другому без какой-либо прямой выгоды для себя. Похоже, что большинство описанных ранее форм поведения не удовлетворяют этому определению, и теоретики игр умеют находить в них «эгоистичные» компоненты. Другие утверждали, что при обсуждении поведения животных следует полностью отказаться от терминов «эгоистичный» и «альтруистический», поскольку они описывают поведение человека и могут не иметь прямого отношения к инстинктивной активности животных.Ясно, однако, что наследственное поведение, которое увеличивает шансы передачи своих генов или части генов, поддерживается естественным отбором и будет сохраняться в будущих поколениях до тех пор, пока это поведение дает преимущество в приспособленности. Это инстинктивное поведение затем может быть применено, в особых обстоятельствах, к другим видам, если оно не снижает приспособленность животного.
Поиск сексуальных партнеров
Не все животные размножаются половым путем, но многие из них сталкиваются с одной и той же проблемой: им нужно найти подходящую пару и часто приходится соревноваться с другими особями, чтобы получить ее.Значительная энергия тратится на поиск, привлечение и совокупление полового партнера. Во время этого процесса происходят два типа отбора, которые могут привести к чертам, важным для воспроизводства, называемым вторичными половыми признаками: интерсексуальный отбор, выбор партнера, когда люди одного пола выбирают партнеров другого пола, и внутриполовой отбор, конкуренция за спариваются между видами представителей одного пола. Интерсексуальный отбор часто бывает сложным, потому что выбор партнера может основываться на множестве визуальных, слуховых, тактильных и химических сигналов.Пример интерсексуального отбора — это когда самка павлина выбирает спариваться с самцом с самым ярким оперением. Этот тип отбора часто приводит к чертам выбранного пола, которые не повышают выживаемость, но являются теми чертами, которые наиболее привлекательны для противоположного пола (часто за счет выживания). Внутрисексуальный отбор включает в себя брачные демонстрации и агрессивные брачные ритуалы, такие как бода баранов головами — в этих битвах побеждает тот, кто способен спариваться. Многие из этих ритуалов отнимают много энергии, но приводят к выбору самых здоровых, сильных и / или доминирующих особей для спаривания.В популяциях животных наблюдаются три общие системы спаривания, все из которых связаны с врожденным поведением, а не с усвоенным: моногамным, полигамным и полиандрическим.
Ссылка на обучение
Посетите этот веб-сайт, чтобы увидеть информативные видеоролики о сексуальном выборе.
В моногамных системах один самец и одна самка спариваются по крайней мере на один сезон размножения. У некоторых животных, таких как серый волк, эти ассоциации могут длиться намного дольше, даже на всю жизнь. Было предложено несколько объяснений этого типа системы спаривания.«Гипотеза защиты партнера» утверждает, что самцы остаются с самкой, чтобы не дать другим самцам спариваться с ней. Такое поведение выгодно в таких ситуациях, когда помощников мало и их трудно найти. Другое объяснение — это «гипотеза помощи самцам», согласно которой самцы, оставшиеся с самкой, чтобы охранять и воспитывать своих детенышей, будут иметь более здоровое потомство. Моногамия наблюдается во многих популяциях птиц, где, помимо родительской заботы со стороны самки, самец также является основным источником родительской заботы о птенцах.Третье объяснение эволюционных преимуществ моногамии — это «гипотеза женского принуждения». В этом сценарии самка гарантирует, что у самца не будет другого потомства, которое могло бы конкурировать с ее собственным, поэтому она активно вмешивается в сигналы самца, чтобы привлечь других самцов.
Полигинное спаривание означает спаривание одного самца с несколькими самками. В этих ситуациях самка должна нести большую часть родительской заботы, поскольку одинокий самец не способен заботиться о таком большом количестве потомков.В полигинии, основанной на ресурсах, самцы соревнуются за территории с лучшими ресурсами, а затем спариваются с самками, которые проникают на территорию, привлеченные ее богатством ресурсов. Самка получает выгоду от спаривания с доминантным, генетически подходящим самцом; однако это происходит за счет отсутствия помощи мужчин в уходе за потомством. Пример можно увидеть в медогиде с желтым крестцом, птице, самцы которой защищают ульи, потому что самки питаются их воском. По мере приближения самок самец, защищающий гнездо, спаривается с ними.Брачные структуры гаремов — это тип полигинной системы, в которой определенные самцы доминируют при спаривании, контролируя территорию с ресурсами. Примером могут служить морские слоны, у которых альфа-самец доминирует при спаривании в группе. Третий тип многоженства — это система лек. Здесь есть общая площадка для ухаживаний, где несколько самцов демонстрируют замысловатые представления для самок, а самки выбирают себе пару из этой группы. Такое поведение наблюдается у нескольких видов птиц, включая шалфейный тетерев и степной цыпленок.
В полиандрических системах спаривания одна самка спаривается со многими самцами. Эти типы систем гораздо реже, чем моногамные и полигамные брачные системы. У морских игл и морских коньков самцы получают яйца от самок, оплодотворяют их, защищают в сумке и дают потомство ([ссылка]). Таким образом, самка может дать яйца нескольким самцам без бремени вынашивания оплодотворенных яиц.
Спаривание с полиандроусом, при котором одна самка спаривается со многими самцами, происходит у морского конька (а) и рыбы-иглы (b).(кредит А: модификация работы Брайана Грэтвика; кредит В: модификация работы Стивена Чайлдса)
Большинство описанных ранее форм поведения были врожденными или, по крайней мере, имели врожденный компонент (можно изучить варианты врожденного поведения). Они передаются по наследству, и поведение не меняется в ответ на сигналы из окружающей среды. И наоборот, усвоенное поведение, даже если оно может иметь инстинктивные компоненты, позволяет организму адаптироваться к изменениям в окружающей среде и модифицируется предыдущим опытом.Простые усвоенные формы поведения включают в себя привыкание и импринтинг — оба они важны для процесса созревания молодых животных.
Привыкание
Привыкание — это простая форма обучения, при которой животное перестает реагировать на раздражитель после периода многократного воздействия. Это форма неассоциативного обучения, поскольку стимул не связан ни с каким наказанием или вознаграждением. Луговые собачки обычно издают тревожный сигнал, когда им угрожает хищник, но они привыкают к звуку человеческих шагов, когда с этим звуком не связано никакого вреда, поэтому они больше не реагируют на них тревожным звонком.В этом примере привыкание относится к звукам шагов человека, поскольку животные по-прежнему реагируют на звуки потенциальных хищников.
Печать
Импринтинг — это тип обучения, который происходит в определенном возрасте или на определенном этапе жизни, который является быстрым и независимым от участвующих видов. Птенцы утки узнают первого взрослого, которого они увидят, свою мать, и завязывают с ней связь. Привычное зрелище — утята, идущие или плывущие вслед за матерью ([ссылка]). Это еще один тип неассоциативного обучения, но он очень важен в процессе созревания этих животных, поскольку побуждает их оставаться рядом с матерью, чтобы они были защищены, что значительно увеличивает их шансы на выживание.Однако, если новорожденные утки увидят человека до того, как увидят свою мать, они запечатлеваются на человеке и будут следовать за ним точно так же, как они следовали бы за своей настоящей матерью.
Привязанность утят к матери — пример импринтинга. (кредит: модификация работы Марка Харкина)
Ссылка на обучение
Международный фонд журавлей помог увеличить мировую популяцию американских журавлей с 21 особи до примерно 600.Отпечаток вылупившихся птенцов был ключом к успеху: биологи носят полные костюмы журавля, чтобы птицы никогда не «видели» людей. Посмотрите это видео, чтобы узнать больше.
Условное поведение — это типы ассоциативного обучения, при котором стимул ассоциируется со следствием. Во время оперантного обусловливания поведенческая реакция модифицируется ее последствиями, касающимися ее формы, силы или частоты.
Классический кондиционер
В классическом кондиционировании реакция, называемая условной реакцией, связана со стимулом, с которым она ранее не была связана, — условным раздражителем.Реакция на исходный безусловный раздражитель называется безусловной реакцией. Наиболее цитируемым примером классической обусловленности являются эксперименты Ивана Павлова с собаками ([ссылка]). В экспериментах Павлова безусловной реакцией было выделение у собак слюноотделения в ответ на безусловный стимул видеть или чувствовать запах их пищи. Обусловливающим стимулом, который исследователи связали с безусловной реакцией, был звонок в колокольчик. Во время кондиционирования каждый раз, когда животному давали пищу, звонили в звонок.Это повторилось во время нескольких испытаний. Через некоторое время собака научилась ассоциировать звон колокольчика с едой и реагировать выделением слюны. По окончании периода кондиционирования собака реагировала выделением слюны на звонок, даже когда безусловный стимул, еда, отсутствовала. Таким образом, звон колокольчика стал условным раздражителем, а слюноотделение — условной реакцией. Хотя некоторые ученые считают, что безусловная и условная реакции идентичны, даже Павлов обнаружил, что слюна у условных собак имеет характерные отличия по сравнению с безусловной собакой.
В классическом павловском ответе собака начинает ассоциировать звонок в колокольчик с едой.
Некоторые ученые полагали, что для этого типа кондиционирования требуется многократное воздействие парных стимулов и ответов, но теперь известно, что это не обязательно во всех случаях, и что определенному условию можно научиться в одном парном эксперименте. Классическая обусловленность — главный принцип бихевиоризма, ветви психологической философии, которая предполагает, что все действия, мысли и эмоции живых существ являются поведением, которое можно лечить путем модификации поведения и изменений в окружающей среде.
Оперантное кондиционирование
При оперантном обусловливании условное поведение постепенно модифицируется его последствиями по мере того, как животное реагирует на раздражитель. Основным сторонником такой обусловленности был психолог Б.Ф. Скиннер, изобретатель ящика Скиннера. Скиннер поместил крыс в свои ящики, в которых был рычаг, который выдавал крысе пищу при нажатии. Хотя изначально крыса несколько раз нажимала на рычаг случайно, в конечном итоге нажатие на рычаг ассоциировалось с получением еды.Этот тип обучения является примером оперантного обусловливания. Оперантное обучение — основа большинства дрессировок животных. Обусловленное поведение постоянно изменяется с помощью положительного или отрицательного подкрепления, часто в виде награды, такой как еда или какой-либо вид наказания, соответственно. Таким образом, животное приучают ассоциировать определенный тип поведения с наказанием или вознаграждением, и со временем его можно побудить к поведению, которое они не стали бы делать в дикой природе, например, к «трюкам» дельфинов. шоу в морском парке развлечений ([ссылка]).
Дрессировка дельфинов путем вознаграждения их пищей является примером положительного подкрепления оперантной обусловленности. (кредит: Роланд Танглао)
Классическая и оперантная обусловленность — неэффективные способы обучения людей и других разумных животных. Некоторые приматы, в том числе люди, могут учиться, подражая поведению других и следуя инструкциям. Развитие сложного языка людьми сделало когнитивное обучение, манипулирование информацией с помощью разума, наиболее известным методом человеческого обучения.Фактически, именно так студенты учатся прямо сейчас, читая эту книгу. По мере чтения учащиеся могут создавать мысленные образы объектов или организмов, воображать изменения в них или их поведение и предвидеть последствия. Помимо визуальной обработки, когнитивное обучение также улучшается за счет запоминания прошлого опыта, прикосновения к физическим объектам, слушания звуков, дегустации еды и множества других сенсорных входов. Когнитивное обучение настолько мощно, что его можно использовать для детального понимания обусловленности.В обратном сценарии кондиционирование не может помочь кому-то узнать о познании.
Классическая работа по когнитивному обучению была проделана Вольфгангом Кёлером с шимпанзе. Он продемонстрировал способность этих животных к абстрактному мышлению, показав, что они могут научиться решать головоломки. Когда банан был подвешен в их клетке слишком высоко, чтобы они могли достать до них, и несколько ящиков были случайным образом расставлены на полу, некоторые шимпанзе могли складывать ящики друг на друга, взбираться на них и подниматься. банан.Это означает, что они могли визуализировать результат складывания ящиков еще до того, как они выполнили действие. Этот тип обучения гораздо более мощный и универсальный, чем кондиционирование.
Когнитивное обучение не ограничивается приматами, хотя они наиболее эффективно его используют. Эксперименты по бегу в лабиринте, проведенные на крысах Х. Блоджетт в 1920-х годах был первым, кто продемонстрировал когнитивные способности простого млекопитающего. Мотивом для животных проложить себе путь через лабиринт был кусок еды на его конце.В этих исследованиях животных в группе I запускали по одной пробе в день, и по завершении пробежки они получали пищу каждый день ([ссылка]). Крыс группы II не кормили в лабиринте в течение первых шести дней, а затем проводили последующие прогоны с пищей в течение нескольких дней. Крысы группы III получали пищу на третий день и каждый последующий день. В результате контрольные крысы, группа I, быстро научились и поняли, как пройти лабиринт за семь дней. Группа III не многому научилась в течение трех дней без еды, но быстро догнала контрольную группу, когда ей дали пищевое вознаграждение.Группа II училась очень медленно в течение шести дней без вознаграждения, чтобы мотивировать их, и они не начали догонять контрольную группу до тех пор, пока не был получен дневной корм, а затем потребовалось на два дня больше, чтобы выучить лабиринт.
Группа I (сплошная зеленая линия) находила пищу в конце каждого испытания, группа II (синяя пунктирная линия) не находила пищу в течение первых 6 дней, а группа III (красная пунктирная линия) не находила пищу в течение работает в первые три дня. Обратите внимание, что крысы, которым давали корм раньше, учились быстрее и в конечном итоге догнали контрольную группу.Оранжевые точки на линиях групп II и III показывают дни, когда в лабиринты добавлялись пищевые награды.
Может быть не сразу очевидно, что этот тип обучения отличается от кондиционирования. Хотя у кого-то может возникнуть соблазн поверить в то, что крысы просто научились находить путь через условную серию поворотов вправо и влево, Е.К. Толмен через десять лет доказал, что крысы в своем сознании создавали представление о лабиринте, что он назвал «познавательная карта.«Это была ранняя демонстрация силы когнитивного обучения и того, что эти способности не ограничиваются только людьми.
Социобиология — междисциплинарная наука, первоначально популяризированная исследователем социальных насекомых Э. Уилсоном в 1970-е годы. Уилсон определил эту науку как «распространение популяционной биологии и эволюционной теории на социальную организацию». 1 Основная идея социобиологии состоит в том, что поведение животных и человека, включая агрессивность и другие социальные взаимодействия, можно объяснить почти исключительно с точки зрения генетики и естественного отбора.Эта наука противоречива; известный ученый, такой как покойный Стивен Джей Гулд, раскритиковал этот подход за игнорирование воздействия окружающей среды на поведение. Это еще один пример дискуссии «природа против воспитания» о роли генетики в сравнении с ролью окружающей среды в определении характеристик организма.
Социобиология также связывает гены с поведением и связана с «биологическим детерминизмом», верой в то, что все виды поведения жестко запрограммированы в наших генах. Никто не оспаривает, что определенные формы поведения могут передаваться по наследству и что естественный отбор играет роль в их сохранении.Применение этих принципов к человеческому поведению вызывает споры, которые не утихают и сегодня.
Поведение — это реакция на раздражители. Это может быть либо инстинктивное / врожденное поведение, на которое не влияет окружающая среда, либо усвоенное поведение, на которое влияют изменения окружающей среды. Инстинктивное поведение включает системы спаривания и способы общения. Приобретенное поведение включает импринтинг и привыкание, обусловливание и, что наиболее важно, когнитивное обучение.Хотя связь между поведением, генетикой и эволюцией хорошо известна, объяснение человеческого поведения как полностью генетического является спорным.
Способность крыс научиться управлять лабиринтом является примером ________.
- отпечаток
- классическое кондиционирование
- оперантное кондиционирование
- когнитивное обучение
Дрессировка животных обычно включает ________.
- отпечаток
- классическое кондиционирование
- оперантное кондиционирование
- когнитивное обучение
Жертвовать жизнью человека ради передачи генов родственников называется ________.
- оперантное обучение
- родовой отбор
- кинезис
- отпечаток
Опишите эксперименты Павлова с собаками как пример классической обусловленности.
У собак выделяется слюна в ответ на еду. Это был безусловный стимул и ответ. У собак, подвергшихся воздействию пищи, в одно и то же время неоднократно звонили в колокольчик, и в конечном итоге они научились ассоциировать звонок с едой. Со временем у собак выделялась слюна, когда звонили в колокольчик, даже при отсутствии еды.Таким образом, звонок стал условным раздражителем, а слюноотделение в ответ на звонок — условной реакцией.
Сноски
- 1 Эдвард О. Уилсон. О природе человека (1978; репр., Кембридж: издательство Гарвардского университета, 2004), xx.
Глоссарий
- агрессивный дисплей
- Визуальная демонстрация членом вида, чтобы отпугнуть других представителей того же вида или других видов
- поведение
- Изменение деятельности организма в ответ на раздражитель
- поведенческая биология
- Изучение биологии и эволюции поведения
- классическое кондиционирование
- Связь определенного стимула и реакции посредством кондиционирования
- когнитивное обучение
- знания и навыки, приобретенные путем манипулирования информацией в уме
- условное поведение
- Поведение, которое ассоциируется с определенным стимулом в результате кондиционирования
- дисплей ухаживания
- Визуальный дисплей, используемый для привлечения помощника
- отвлекающий дисплей
- Визуальный дисплей, используемый для отвлечения хищников от места гнездования
- этология
- Биологические исследования поведения животных
- шаблон фиксированного действия
- серия инстинктивных форм поведения, которые, будучи инициированы, всегда завершаются независимо от изменений в окружающей среде
- кормодобывание
- видов поведения, которые используют для поиска пищи
- привыкание
- Способность вида игнорировать повторяющиеся стимулы, не имеющие последствий
- печать
- идентификация родителей новорожденными как первый организм, который они видят после рождения
- врожденное поведение
- инстинктивное поведение, не измененное изменениями в окружающей среде
- интерсексуальный отбор
- Выбор желанной спутницы противоположного пола
- внутрисексуальный отбор
- Соревнование между представителями одного пола за партнершу
- родовой отбор
- приносить в жертву свою жизнь ради передачи своих генов будущим поколениям родственниками
- кинез
- ненаправленное движение организма в ответ на раздражитель
- усвоенное поведение
- поведение, которое реагирует на изменения в окружающей среде
- миграция
- дальние сезонные перемещения видов животных
- моногамия
- Система спаривания, при которой один самец и одна самка остаются спарившимися по крайней мере в течение одного брачного сезона
- оперантное кондиционирование
- усвоенных моделей поведения в ответ на положительное и / или отрицательное подкрепление
- многомужество
- Система спаривания, при которой одна самка спаривается со многими самцами
- полигиния
- Система спаривания, при которой один самец спаривается со многими самками
- рефлекторное действие
- действие в ответ на прямое физическое раздражение нерва
- сигнал
- способ общения между животными, в том числе с помощью обоняния, слуха, зрения или осязания
- такси
- направленное движение в ответ на раздражитель
стереотипный ответ | биология | Британника
стереотипный ответ , неизученная поведенческая реакция организма на какой-либо раздражитель окружающей среды.Это адаптивный механизм, который может выражаться по-разному. Все живые организмы демонстрируют один или несколько типов стереотипной реакции.
Общие положения
Способность к необученному поведению генетически определяется во многом в том же смысле, что и положение, размер, форма и функция органов. Подобно структурным особенностям, стереотипные ответы являются результатом непрерывного процесса эволюционной модификации и уточнения. Те действия, которые наиболее успешно помогают животному или растению в его основных побуждениях ( e.g., размножение, поиск пропитания, спасение от хищников) — те, которые, скорее всего, сохранятся в следующих поколениях. По мере изменения условий окружающей среды внутренне определенные реакции также изменяются, чтобы обеспечить продолжение вида.
Проблемы, возникающие при изучении стереотипных ответов, многочисленны и разнообразны. Особые реакции у животных не всегда поддаются идентификации в высокоразвитых формах, потому что усвоенные модели поведения затемняют лежащее в основе необученное поведение; кроме того, стереотипные ответы служат строительными блоками инстинктивного поведения, сложность которого может затенять составные части (см. инстинкт).У низших животных, как и у растений, у которых обученное поведение отсутствует или отсутствует, анализ поведенческих механизмов ограничен тем фактом, что многие из наиболее фундаментальных клеточных процессов недостаточно изучены.
Поведение животных как раздел психологии представляет собой слияние дисциплин этологии и сравнительной психологии. Большую часть пионерской работы над стереотипными реакциями животных проделали этологи. В первой половине 20-го века, когда были заложены многие основы экспериментальной психологии, этологи (по большей части европейцы) интересовались поведением насекомых, рыб и птиц и особенно интересовались эволюцией инстинктов. .Психологами-сравнителями в этот период становления были в основном американцы. Они изучали в первую очередь поведение обычных лабораторных животных, таких как морские свинки, мыши, крысы и обезьяны, и их интерес, как правило, был сосредоточен на влиянии окружающей среды на поведение, а не на генетическом влиянии. С 1950-х годов психологи в целом признали, что факторы окружающей среды и генетические факторы играют важную роль в любом биологическом явлении. Однако вследствие раздельного развития этологии и сравнительной психологии возникли некоторые трудности в использовании терминологии.Немецко-американский биолог Жак Леб применил термин «тропизм» ко всем ориентированным движениям организмов и предположил, что все поведение состоит из тропизмов. Впоследствии, во избежание путаницы, термины «налоги» (в единственном числе: такси) и кинезы были введены другими исследователями для обозначения реакций животных, отличных от реакций оседлых, похожих на растения форм. Эти термины также применялись к определенным движениям растений. Хотя у растений происходит множество дискретных стереотипных ответных движений, особенно у высших форм, таких как цветковые растения, эти автономные движения обычно происходят слишком медленно, чтобы их можно было обнаружить путем случайного наблюдения.То, что движения растений или органов растений действительно имеют место, может быть наглядно продемонстрировано с помощью покадровой фотографии, в которой отдельные фотографии делаются через равные промежутки времени, равные секундам или нескольким дням и более. Затем фотографии сравниваются или просматриваются в быстрой последовательности как движущееся изображение.
Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчасТипы стереотипных ответов
Стереотипный ответ у животных можно разделить на следующие четыре категории: неорганизованный или плохо организованный ответ, рефлекторные движения определенной части организма, рефлекторная активность всего организма и инстинкт.
Неорганизованные или плохо организованные реакции дают ранние эмбрионы или животные (например, губки), у которых отсутствует нервная система.
Собственно рефлексы или рефлекторно-дуговые движения включают такие реакции, как немедленное отдергивание руки при прикосновении к горячей поверхности. Основными компонентами рефлекторной дуги являются рецептор, или сенсорно-нервная клетка, которая воспринимает раздражитель, и аффектор, нервная клетка, которая непосредственно активирует мышцу. Это теоретический минимум, а не наблюдаемое функциональное расположение клеток в теле животного (см. Инстинкт: разновидности инстинктивного поведения).
Рефлекторная деятельность
Рефлекторная деятельность целых организмов может быть неориентированной или ориентированной. Неориентированные реакции включают кинезы — ненаправленное ускорение или замедление скорости передвижения или частоты перехода от покоя к движению (ортокинез), или частоты или количества поворотов всего животного (клинокинез), скорость частоты зависит от интенсивности стимуляции. . Примеры ортокинеза наблюдаются у миног, которые более активны при высокой интенсивности света, и у тараканов, которые более активны при низкой интенсивности; плоские черви и многие виды личинок мух, среди других беспозвоночных, обладают ортокинезом.Клинокинез хорошо демонстрируют движения мокрицы ( Porcellio scaber ). Когда мокрицы помещаются в сухой воздух, они активно ползают, но без направления, пока постепенно не обезвоживаются. Когда мокрицы помещаются во влажный воздух, они сначала двигаются, но всякая активность, которую они проявляют, вскоре прекращается, и они затихают. Мыши, помещенные в контейнер с сухим воздухом на одном конце и влажным — на другом, постепенно собираются на влажном конце. Эта передача достигается скорее случайными, чем направленными движениями.
Ориентированная рефлекторная деятельность целых организмов включает тропизмы, налоги и ориентацию под углом. Тропизмы у животных — это те направленные движения роста-искривления сидячих (, т. Е. сидячих) форм, которые приводят к одинаковой интенсивности стимуляции симметрично расположенных частей тела. Эти движения демонстрируют гидроидные животные, такие как Eudendrium .
Границы | На пути к интегративному пониманию социального поведения: новые модели и новые возможности
Введение
Социальное поведение состоит из взаимодействий между сородичами и приводит к отношениям переменной формы, продолжительности и функции.Социальные взаимодействия создают основу для широкого спектра поведенческих явлений, включая многие сложные формы сотрудничества и конфликтов, которые представляют особый интерес для поведенческих биологов (Nowak, 2006). ). Например, групповая жизнь и совместное воспитание молодняка — это формы социальности, возникающие в результате социальных взаимодействий между людьми. Социальность (в широком смысле определяемая как групповая жизнь) представляет собой очевидный эволюционный парадокс, заключающийся в том, что люди могут нести расходы на приспособленность, участвуя в этих взаимодействиях (Alexander, 1974). ; Гриффин и Уэст, 2002 ).В совокупности эти явления составляют основу некоторых из самых сложных и интригующих обществ, наблюдаемых в природе (Wilson, 1975). ). У людей социальные отношения являются важными компонентами благополучия (House et al., 1988 ), и может привести как к положительным, так и к отрицательным последствиям для здоровья (Berkman, 1984 ; Учино и др., 1996 г. ).
Хотя не все формы социального взаимодействия так сложны или трудны для интерпретации, как кооперативное поведение, социальное поведение является фундаментальным атрибутом биологии многих видов и является предметом обширных исследований ученых из разных дисциплин.Несмотря на широкое применение в фундаментальной и прикладной науке, разработка всеобъемлющей основы для объяснения непосредственных и конечных основ социальных взаимодействий — быстро развивающаяся тема в поведенческих исследованиях (Owens, 2006). ) — оказалось проблемным (Johnstone, 2000). ). Традиционные модели социальной изменчивости сосредоточены в первую очередь на экологических и эволюционных факторах, которые приводят к определенным формам социального взаимодействия (например, групповая жизнь: Emlen and Oring, 1977 ; Эмлен, 1995 г. ; Brashares и Arcese, 2002 г. ; Cahan et al., 2002 г. ). Однако недавние достижения в области нейробиологии, эндокринологии и генетики позволили получить знания и инструментарий, необходимые для более интегративных подходов к пониманию социальных взаимодействий (Young and Wang, 2004). ). Очень важно разработать концептуальную основу, включающую эти новые ресурсы; в дополнение к информационному обеспечению исследований на нескольких уровнях организации (например, от молекулярных до эволюционных процессов), такие достижения в нашем понимании социальных взаимодействий должны иметь приложения к благосостоянию и здоровью человека, включая улучшение диагностики и лечения некоторых психических расстройств (Bartz and Hollander, 2006 г. ; Лим и Янг, 2006 ; Bosch et al., 2009 г. ; МакГроу и Янг, 2010 г. ). Например, расстройство аутистического спектра и шизофрения характеризуются выраженным дефицитом социального познания и социального поведения (Couture et al., 2010 ), а нарушение социальных отношений приводит к увеличению риска депрессии (Zisook and Shuchter, 1991 ).
Здесь мы очерчиваем интегративную основу для изучения социального поведения, которая обращается к четырем важным элементам современной поведенческой биологии.Во-первых, в рамках данной концепции освещаются недавние генетические, эндокринные и нейробиологические достижения, которые создают новые возможности для изучения механизмов социального поведения в экологическом контексте. Таким образом, структура объединяет хорошо разработанные, но традиционно отдельные области исследования и способствует использованию новых технологий для изучения социальных взаимодействий в нетрадиционных исследуемых организмах и в внелабораторных условиях. Во-вторых, структура помогает генерировать прогнозы о социальных отношениях, которые можно проверить в полевых условиях или в лаборатории, и которые помогают различать причины и последствия социальных взаимодействий.В-третьих, структура исследует разнообразие механизмов, лежащих в основе социальной изменчивости, тем самым облегчая идентификацию крупномасштабных паттернов в эволюции генетических, эндокринных и нейронных основ социального поведения. В-четвертых, использование нетрадиционных и естественных систем является ключом к пониманию естественной изменчивости социального поведения и социальности, и это прольет свет на механизмы, поддерживающие социальную изменчивость.
Одним из результатов интегративного подхода, который мы описываем, является переключение внимания на исследования новых систем животных, некоторые из которых были хорошо изучены экологами, но которые ранее не рассматривались в качестве объектов для более механистических исследований социальных отношений.В целом, этот обзор должен стимулировать новое сотрудничество между поведенческими биологами из разных дисциплин, что приведет к захватывающим изменениям в нашем понимании социального поведения, включая социальные взаимодействия, лежащие в основе множества человеческих расстройств.
Уровни анализа: новый взгляд на Тинберген
Традиционные исследования социального поведения, как правило, следовали рамкам, установленным Тинбергеном (1963). , который выдвинул четыре ортогональных объяснения разнообразия поведения, наблюдаемого в природе: (1) непосредственная причинная связь, (2) развитие, (3) функция и (4) эволюционная история.Непосредственные причины поведения включают физиологические, генетические или онтогенетические факторы, а также взаимодействия между этими аспектами биологии организма. Например, определение гормонов, которые модулируют формирование привязанности, или генов, связанных с социальными связями, может стать исследованиями, направленными на непосредственную причинную связь. Объяснения, связанные с развитием, включают любые изменения в поведении в результате онтогенетических процессов или переживаний. Например, определение роли раннего опыта в влиянии на решение о том, оставаться ли в натальной группе или разойтись, было бы исследованием, которое рассматривало бы вопрос развития.Функциональные причины описывают, как поведение в настоящее время влияет на выживание и воспроизводство. Например, демонстрация того, что люди, живущие в социальных группах определенного размера, имеют более высокую выживаемость и репродуктивный успех, отвечает на адаптивные вопросы. Наконец, исторические объяснения используют филогенетические отношения между видами, чтобы получить представление об эволюционной выгоде или потере поведенческой черты. Например, демонстрация того, что социальность развивалась (или не развивалась) в определенной группе организмов, может указывать на филогенетическую предрасположенность или ограничение на социальность.Такое исследование обращается к историческим объяснениям социальности.
Эти вопросы (или подходы) ортогональны (т. Е. Не связаны между собой), потому что для любого из четырех вопросов существует ряд взаимоисключающих гипотез, которые могут их объяснить. Другими словами, логично спросить, усиливает ли привязанность гормон а или гормон b. Логично спросить, могут ли животные, живущие в группе, иметь более высокую выживаемость, живя в большей или меньшей группе. Однако ответ на вопрос о конкретном гормоне, который усиливает привязанность, не влияет на оптимальный размер группы, и наоборот.Таким образом, в значительной степени каждый подход не связан между собой.
Эти четыре подхода к изучению поведения можно разделить на два уровня анализа (Sherman, 1988 ). Непосредственная причинность и развитие включают приближенных объяснений, которые отвечают на вопросы, касающиеся того, как поведения возникают механистически. История функций и эволюции включает окончательных объяснений, которые отражают , почему поведения эволюционировали или сохраняются.Эти четыре объяснения дополняют друг друга, а не исключают друг друга, и любая черта может быть изучена на любом (и, вероятно, на всех) из этих уровней исследования (Bolhuis and Verhulst, 2009). ). Мы предполагаем, что включение каждого из этих уровней анализа приведет к более комплексному пониманию социального поведения, которое принесет пользу всем дисциплинам.
Исторически сложилось так, что поведенческие экологи уделяли особое внимание вопросам конечного уровня, в то время как биомедицинские исследователи сосредотачивались на явлениях непосредственного уровня, создавая концептуальный и практический разделение между этими исследовательскими установками, которое включает появление различных терминологий, методологий и исследуемых организмов.Однако недавно специалисты по поведенческой экологии, традиционно работающие на местах, вновь открыли для себя важность изучения ближайших механизмов (например, генетических, физиологических, нейроэндокринных и нейрофизиологических процессов), лежащих в основе поведения, чтобы лучше понять эволюцию поведения (Stamps, 1991). ; Оуэнс, 2006 г. ). Этому растущему признанию важности исследований непосредственного уровня способствовали технические достижения, которые сделали механистические исследования более податливыми и открыли новые направления для понимания социального поведения.В частности, недавние достижения, связывающие физиологию, геномику и поведение, открывают новые захватывающие возможности для интеграции вопросов конечного и ближайшего уровней для изучения социальных взаимодействий, тем самым создавая новые связи между социальным поведением и потенциальными причинами поведенческой дисфункции человека (Робинсон и др., 2008 г. ). Мы твердо убеждены в том, что биомедицинские исследования также могут получить огромную пользу от включения взглядов поведенческих экологов на открытие новых модельных организмов и уточнение экспериментальных парадигм в этологически значимой манере.
Важность природных популяций
Как этолог, исследования поведения Тинбергена основывались в основном на наблюдениях за свободноживущими животными, которые сталкивались с проблемами выживания и воспроизводства в сложных условиях «реального мира». В то время как исследованиям естественных популяций обычно не хватает строгого контроля экологической и межличностной изменчивости, характерной для лабораторных исследований, данные, полученные на свободноживущих животных, имеют решающее значение для всестороннего понимания изменчивости в социальном поведении по нескольким причинам.Во-первых, наблюдения за свободноживущими особями, ведущими себя типично для видов, могут быть использованы для подтверждения данных лабораторных исследований путем подтверждения того, что поведенческие взаимодействия, наблюдаемые в неволе, характерны для взаимодействий, наблюдаемых в естественной среде (Holmes and Sherman, 1982). ; Шрадин и Пиллэй, 2003 ). Во-вторых, когда наблюдения за содержащимися в неволе и свободноживущими особями различаются, сравнения данных, полученных в этих двух различных контекстах, могут быть использованы для получения нового понимания факторов, регулирующих поведение (Calisi and Bentley, 2009 ).В-третьих, исследования свободноживущих животных обеспечивают экологический контекст, необходимый для оценки функциональных последствий социального поведения. В частности, данные природных популяций позволяют исследователям оценить репродуктивные (функциональные) последствия поведения для репродуктивной системы и выживания в тех отборных условиях, в которых оно обычно происходит; в отсутствие такой информации может быть сложно определить функциональные и, следовательно, потенциальные вариации эволюционной значимости социальных взаимодействий.Таким образом, хотя многие из недавних эндокринных, нейронных и генетических достижений в нашем понимании поведения неизбежно начались в лаборатории, применение этих идей и технологий к естественным популяциям имеет важное значение для создания полного понимания причин различий в социальном поведении.
Генетические и геномные достижения: новые возможности для изучения природных систем
Исследования людей, а также традиционных животных моделей (например, Drosophila melanogaster , Caenorhabditis elegans , Mus musculus , Rattus norvegicus ) были неоценимы для идентификации генов, участвующих в регуляции социального поведения ( Робинсон и др., 2005 г. ). Учитывая, что функция генов часто сохраняется, мы ожидаем, что многочисленные гомологичные локусы ждут изучения в нетрадиционных модельных системах, что позволит исследовать экспрессию генов у свободноживущих животных в различных условиях окружающей среды. Например, в лабораторных популяциях D. melanogaster ген поиска пищи отвечает за различия в двигательной активности, связанной с получением пищи; гомологи кормящихся , как было показано, влияют на возраст начала кормления у свободноживущих медоносных пчел ( Apis mellifera ) (Amdam and Page Jr., 2010 г. ; Робинсон и др., 2008 г. ;). У людей аллельные варианты гена Drd4 связаны с личностными чертами и стремлением к новизне; этот локус, по-видимому, играет аналогичную роль в природных популяциях птиц, таких как синицы ( Parus major ; Fidler et al., 2007 ). Наконец, полиморфизм гена рецептора вазопрессина, Avpr1a , связан с поведением парных связей как у людей, так и у степных полевок ( Microtus ochrogaster , Hammock and Young, 2005 ; Дональдсон и Янг, 2008 г. Это сохранение функции генов в порой разрозненных таксономических группах обеспечивает важную основу для расширения исследований генетических основ социального поведения на естественные популяции животных.
В то же время постоянно расширяющийся набор инструментов для генетических исследований делает все более возможными экспериментальные исследования генетических механизмов, лежащих в основе социального поведения. Например, использование миРНК для подавления экспрессии гена у зябликов ( Taeniopygia guttata ) показало, что гомолог FoxP2 (ген, который, как считается, участвует в человеческом языке) также играет важную роль в обучении вокалу у певчих птиц. (Haesler et al., 2007 ).Опосредованный вирусным вектором перенос генов использовался для сверхэкспрессии генов в определенных областях мозга для определения функции генов у таких видов, как степные полевки, что дало критическое представление о причинах различий в социальной системе и системе спаривания у разных видов (Lim et al., 2004 ; Росс и др., 2009 г. ). Трансгенные технологии, хотя в настоящее время ограничены несколькими видами, разрабатываются для таких разнообразных нетрадиционных модельных организмов, как степные полевки, трехиглая колюшка ( Gasterosteus aculeatus, ) и нечеловеческие приматы (Hosemann et al., 2004 г. ; Дональдсон и др., 2009 г. ; Сасаки и др., 2009 г. ). По мере того, как такие технологии становятся более простыми в использовании, диапазон видов, к которым они применяются, должен расти, что дает все более богатую картину генетических основ вариаций в социальном поведении.
Наконец, для организмов, у которых отсутствует полностью секвенированный геном, исследования транскриптомов (т. Е. Количественная оценка экспрессии генов в масштабе всего генома) предоставляют удобные средства идентификации генов, лежащих в основе социального поведения.ДНК-микрочипы медоносных пчел были использованы для разработки одного из первых полногеномных взглядов на социальную структуру (Robinson et al., 2008 ) и исследования муравьев показали, что сходные гены лежат в основе социальной изменчивости (Ingram et al., 2005 ) в обеих линиях насекомых. Микромассивы также использовались у меченосцев ( Xiphophorus nigrensis ), чтобы определить, как экспрессия генов различается между социальными и сексуальными сигналами (Cummings et al., 2008 ) и африканских цихлид ( Astatotilapia burtoni ) для выявления генов-кандидатов, связанных с социальным поведением, включая локусы, которые активируются у доминантных, но не подчиненных особей (Renn et al., 2008 г. ). Быстрое развитие высокопроизводительных и недорогих технологий геномного секвенирования «следующего поколения» делает все более возможным исследование активности генов в различных нетрадиционных системах животных.
Достижения нейроэндокринной системы: улучшение понимания социальной изменчивости
Более глубокое понимание того, как нейроэндокринные процессы регулируют поведение, представляет собой еще одну область, в которой исследования природных популяций могут дополнить наши быстро расширяющиеся знания о традиционных моделях животных.В частности, прогнозирование того, как нейроэндокринная активность должна влиять на социальное поведение свободноживущих животных, является центральным для разработки интегративной модели социальности (Nunes et al., 1999 ; Райан и Ванденберги, 2002 г. ). Ниже мы выделяем несколько интересных областей исследования нейроэндокринных основ социального поведения. Два из них (уровни циркулирующих гормонов, плотность рецепторов и экспрессия) представляют собой пути, свидетельства которых уже собираются в отношении нейроэндокринных или нейроанатомических влияний на социальное поведение.Хотя для третьего пути (связывание глобулинов) доступно мало эмпирических данных, этот аспект нейроэндокринной функции, по-видимому, имеет большой потенциал для будущих исследований ближайших основ социальных вариаций.
Уровни циркулирующих гормонов
Большая часть исследований, связывающих изменчивость окружающей среды, нейроэндокринную функцию и социальное поведение позвоночных, была сосредоточена на циркулирующих концентрациях стероидов (Wingfield et al., 1990 ; Schoech et al., 2004 ; Пфафф, 2005 г. ), особенно глюкокортикоиды (Creel, 2001 ; Пфафф, 2005 г. ).Например, единичные племенные общества с коллективной заботой о потомстве (т. Е. Одна самка доминирует над воспроизводством, а не родительские особи заботятся о потомстве, не являющемся потомком; Silk, 2007 ) может быть избирательно выгодной социальной стратегией в стрессовой среде, в которой независимое воспроизводство редко бывает успешным (Moehlman, 1979 ). В этих условиях члены группы могут производить более низкие уровни глюкокортикоидов, когда размер группы достигает или ниже порогового значения (Silk, 2007 ), что, в свою очередь, снижает социальный стресс и способствует совместному выращиванию потомства.В качестве альтернативы, социальные животные могут испытывать хронически высокие уровни гормонов стресса, что может иметь негативные последствия для приспособленности и, возможно, объяснять результаты приспособленности групповой жизни, специфичные для среды обитания, и установление социальных иерархий (Creel, 2001). ; Янг и др., 2006 ; Рубинштейн, 2007 ). Требуется экспериментальная демонстрация того, являются ли вариации глюкокортикоидов причиной или следствием социальных различий (Rubenstein and Shen, 2009). ). Имеются некоторые свидетельства снижения количества андрогенов у не размножающихся мужчин-аллопатентов, но, как и в случае с глюкокортикоидами, паттерны андрогенных вариаций не совпадают для разных видов (Schoech et al., 2004 г. ). Например, у полосатых мышей ( Rhabdomys pumilio ) — социально гибких видов, у которых самцы демонстрируют альтернативную тактику спаривания и разный уровень родительской заботы — подчиненные самцы-«бродяги» имеют более высокий уровень тестостерона, чем доминирующие территориальные самцы (Schradin et al. , 2009 г. ). Есть также некоторые свидетельства того, что пролактин может играть причинную роль в подчиненном вспомогательном поведении у некоторых птиц (Buntin, 1996 ; Schoech et al., 2004 ).
Плотность и экспрессия рецептора
У грызунов внутривидовые вариации социальной организации и поведения, такие как социальные связи и родительская забота, были связаны с вариациями плотности нейропептидных рецепторов в головном мозге (Hammock and Young, 2004). ; Оласаль и Янг, 2006a , б ; Бери и др., 2008 г. ). У высоко социальных и моногамных видов рецепторы окситоцина и вазопрессина сосредоточены в областях мозга, связанных с вознаграждением и подкреплением, в то время как у немоногамных видов их нет (Young and Wang, 2004 ).Перенос генов вирусного вектора использовался для управления экспрессией нейропептидных рецепторов в этих областях мозга у полевок ( Microtus, spp.), Тем самым демонстрируя причинную связь между экспрессией рецепторов и этими аспектами поведения (Lim et al., 2004 ). На генетическом уровне полиморфизмы в промоторных областях генов, кодирующих эти рецепторы, по-видимому, вносят вклад в разнообразие паттернов экспрессии рецепторов (Hammock and Young, 2004). ). Окситоцин-подобные рецепторы также могут влиять на социальную жизнь птиц (Goodson et al., 2009 г. ). Замечательная пластичность систем рецепторов окситоцина и вазопрессина обеспечивает потенциальный механизм для создания разнообразия социальных фенотипов как мужчин, так и женщин. Дальнейшие исследования необходимы для определения значимости этих рецепторных систем в формировании социальных и репродуктивных взаимодействий в естественных популяциях других таксонов. Виды с популяциями, которые демонстрируют внутривидовую географическую изменчивость в социальных системах спаривания (Roberts et al., 1998 ) могут быть особенно сильными кандидатами для изучения вариаций нейропептидных систем.
Связывающие глобулины
Стероид-связывающие белки — это гликопротеины, которые транспортируют гормоны осей гипоталамус-гипофиз-надпочечники (HPA) и гипоталамо-гипофиз-гонад (HPG) к тканям-мишеням (Westphal, 1983). ; Бройнер и Орчиник, 2002 г. ). Функция связывания глобулинов широко обсуждается. Неясно, действуют ли связывающие глобулины просто для транспортировки стероидов к тканям-мишеням (гипотеза носителя) или они действуют как губка, связывая избыточные стероиды, чтобы предотвратить их попадание в ткани-мишени, тем самым защищая ткани от высоких уровней циркулирующих стероидов (гипотеза буфера). (Ромеро, 2002 ).Более того, функция стероид-связывающих белков может различаться в зависимости от таксона. Например, в отличие от млекопитающих, у птиц отсутствует специфический для половых стероидов связывающий глобулин; вместо этого половые стероиды (например, тестостерон) связываются с кортикостероидсвязывающими глобулинами (CBG, Deviche et al., 2001 ). У содержащихся в неволе крыс подчиненные особи имели более низкие концентрации CBG, чем доминирующие особи (Spencer et al., 1996 ; Стефанский, 2000 ), в то время как у лошадей социальный стресс вызвал снижение связывающей способности CBG (Alexander and Irvine, 1998 ).Эти результаты предполагают потенциально важную связь между стрессом, CBG и межличностными вариациями в социальном поведении. Следовательно, роль CBG и других связывающих глобулинов заслуживает дальнейшего рассмотрения в исследованиях социального поведения позвоночных.
Продолжение исследований того, как нейроэндокринные механизмы влияют на социальное поведение различных организмов, приведет к лучшему пониманию эволюции механистического разнообразия и, в конечном итоге, позволит нам делать более конкретные прогнозы.Эти способности к прогнозированию появятся в результате изучения разнообразия видов, проявляющих разнообразие социального поведения.
Интегративный подход: направление будущих исследований
Рисунок 1 иллюстрирует, как условия окружающей среды и естественный отбор могут взаимодействовать, чтобы сформировать непосредственные механизмы, лежащие в основе вариации социальных взаимодействий, и, в свою очередь, как эти механизмы могут влиять на последствия различий в социальном поведении для приспособленности. Пути связывают социальные и экологические реакции на физиологические и поведенческие изменения человека.Мы считаем, что обе дисциплины (поведенческая экология и биомедицинские исследования) могут получить огромную пользу от принятия этой интегративной точки зрения.
Рис. 1. Интегративная структура для изучения социального поведения. Эта структура соответствует рекурсивной эволюционной парадигме (Feder et al., 2000 ), при этом генотип человека работает во взаимодействии с окружающей средой, создавая данный фенотип (вставка A). Это имеет адаптивные и неадаптивные последствия, которые определяют генотипы в следующем поколении (вставка B).Социальные фенотипы могут влиять на окружающую среду множеством способов, указывая на динамические двунаправленные отношения, которые могут влиять на процессы в обоих блоках. Поскольку среда, в свою очередь, влияет как на ближайшие механизмы, так и на конечные процессы, важно изучить и то, и другое, чтобы получить интегральное понимание социального поведения. Например, если социальное поведение снижает угрозу хищничества, поведенческие биологи должны изучить, как это снижение влияет на нейроэндокринные механизмы, лежащие в основе этого поведения, в дополнение к тому, как оно влияет на избирательный ландшафт.Более того, поскольку непосредственные механизмы (вставка A) сами по себе подвержены эволюционной динамике (вставка B), существует потребность в исследованиях, измеряющих отбор и наследуемость этого нейроэндокринного механизма, а не просто прямое влияние на социальное поведение. Эта модель, хотя и не исчерпывающая, обеспечивает основу и руководство для синтетических исследований социальности.
Направления интегративных исследований
Ниже мы выделяем несколько важных областей исследований, которые расширяются за счет интегративного подхода к социальному поведению; мы ожидаем, что появятся и другие, новые направления исследований, поскольку дополнительные механизмы и методы будут включены в интегративную структуру, такую как мы предлагаем.Чтобы подчеркнуть полезность этой концепции в объединении традиционных и новых взглядов на социальное поведение, мы составили наш обзор с точки зрения четырех ортогональных исследовательских вопросов, обозначенных Тинбергеном почти 50 лет назад:
(1) Непосредственная причинность . Окружающая среда может вносить вклад во внутривидовые вариации поведенческих фенотипов путем изменения ближайших механизмов, управляющих поведением, включая аллельные вариации, экспрессию генов и нервную активность. Например, степные полевки, обитающие в экологически разных популяциях, демонстрируют заметные различия в социальном поведении (Roberts et al., 1998 г. ). Янг и Ван (2004) показали, что это изменение в социальном поведении полевок связано с изменением плотности рецепторов окситоцина / вазопрессина в головном мозге, причем экспрессия этих рецепторов опосредуется генами oxtr и avpr1a . Наша модель объединяет эту информацию, генерируя следующие прогнозы: (а) непосредственные экологические причины социального поведения, специфичного для среды обитания, включают такие факторы, как распределение пищевых ресурсов или открытость среды обитания, (б) дифференцированный отбор, являющийся результатом этого экологического разнообразия, приводит к популяции -уровневые различия в частоте аллелей avpr1a и (c) аллельные различия в экспрессии гена avrp1a объясняют популяционно-специфические плотности нейропептидных рецепторов.
(2) Онтогенез . Изменения в физической или социальной среде могут влиять на время онтогенетических изменений (развитие нервной системы, эндокринная функция, экспрессия генов), которые опосредуют социальное поведение. Например, натальная филопатрия (оставаясь в социальной группе, в которой родился человек) широко признается как важный процесс, ведущий к формированию социальных групп (Emlen, 1995 ). Экологические причины натальной филопатрии обычно связывают с преимуществами филопатрии (например,g., косвенная пригодность) или экологические ограничения на расселение (например, насыщение среды обитания). Nunes et al. (1999) продемонстрировали, что распространение суслика Белдинга ( Urocitellus beldingi ) опосредовано организационными эффектами тестостерона во время раннего постнатального развития. В рамках такой системы млекопитающих наша структура могла бы интегрировать эти идеи, генерируя следующие прогнозы: (а) ограниченные ресурсы приводят к снижению выработки ГнРГ гипоталамусом, реакции гипофиза на ГнРГ и экспрессии рецепторов лютеинизирующего гормона во время раннего постнатального развития, что приводит к меньше диспергаторов и большая натальная филопатрия, (б) люди, оставшиеся в группе, имеют более высокую инклюзивную приспособленность, чем те, которые расходятся или не образуют группы, и (в) формирование социальной группы снижает уровень тестостерона в раннем постнатальном развитии, что приводит к большему рассредоточению и снижение филопатрии.Другие области исследования включают в себя то, как (i) экология влияет на состояние матери и, следовательно, на развитие потомства и поведение взрослых (например, влияние внутриутробного положения на социальное поведение; Ryan and Vandenbergh, 2002 ) и (ii) экологические и социальные различия влияют на генетические и гормональные пути, ведущие к развитию аллопатической помощи.
(3) Функция . Последствия социального поведения для фитнеса часто зависят от среды обитания (Silk, 2007). ).Индивиды сталкиваются с изменчивостью окружающей среды и соответственно реагируют как в онтогенезе (который определяет фенотип), так и в повседневном, сезонном и факультативном опыте. Возникающее в результате социальное поведение (фенотип) имеет свои издержки и выгоды, величина которых определяет приспособленность (репродуктивный успех на протяжении всей жизни). Эти идеи можно проверить в контексте социального поведения, такого как групповой поиск пищи, который, как считается, снижает риск нападения хищников. Согласно этой гипотезе, поиск пищи в обществе избирательно выгоден в открытых местообитаниях или в местообитаниях с многочисленными хищниками (Ebensperger, 2001). ; Brashares и Arcese, 2002 г. ).Однако социальный поиск пищи требует затрат и, следовательно, может происходить меньше в местообитаниях с низким давлением хищников. Ожидается, что в этих экологических и социальных условиях будут активированы различные нервные и сенсорные пути ( sensu Hermes et al., 2009 ). Например, если функция социального поиска пищи состоит в снижении риска поедания хищников, то ожидается снижение аллостатической нагрузки, когда особи кормятся вместе. Эта гипотеза предсказывает, что (а) индивидуумы имеют более высокие уровни циркулирующих глюкокортикоидов в популяциях с многочисленными хищниками и (б) уровни циркулирующих глюкокортикоидов снижаются с увеличением числа особей на группу кормов в популяциях с высоким уровнем риска хищничества.
(4) История эволюции . Хотя таких исследований в основном нет, каждая из вышеперечисленных тем может быть решена с помощью сравнительных исследований, проводимых в филогенетическом контексте. Изучая закономерности механистического разнообразия в широком спектре таксонов, для которых известны филогенетические отношения, должно быть возможным выявить ранее нераспознанные примеры эволюционного расхождения, конвергенции и сохранения механизмов социального поведения. Социальные фенотипы могут быть ограничены генетической архитектурой ближайших механизмов, которая зависит от истории эволюции.После того, как эти явления будут всесторонне охарактеризованы, можно будет изучить эволюцию механизма новыми и всесторонними способами. В частности, как только эти эволюционные паттерны будут идентифицированы, мы сможем завершить гораздо более содержательный анализ того, каким образом экологические факторы и факторы жизненного цикла влияют на механизмы социального поведения, тем самым связав исследования механизмов с вопросами конечного уровня, наиболее важными для них. часто обращаются к поведенческим экологам. Например, по мере того, как мы собираем больше информации о связи между системами спаривания и экспрессией нейропептидных рецепторов, лежащих в основе социальной изменчивости, можно будет использовать филогенетически контролируемые сравнительные подходы.Основываясь на экологических данных (Emlen and Oring, 1977 ) и нейроанатомической (Young and Wang, 2004 ) теории, мы ожидаем, что для разных таксонов экспрессия нейропептидных рецепторов, лежащих в основе систем спаривания (например, окситоцина, вазопрессина), будет одинаковой у видов, обитающих в аналогичных средах обитания.
Эволюционный контекст для понимания механизмов социальной изменчивости
Как отмечалось выше, эволюционный подход к изучению непосредственных механизмов, лежащих в основе социального поведения, может привести к новому, сравнительному пониманию того, как такие механизмы развиваются и поддерживаются в популяциях животных.Мы считаем, что этот результат комплексного подхода к социальному поведению настолько важен, что мы хотим подробнее остановиться на этой теме на следующих примерах:
(1) Дивергенция . Есть несколько способов решить механистическую проблему, ведущую к потенциальному значительному эволюционному расхождению в механистических основах социального поведения. Это изменение может быть очевидным, например, в способах регуляции гормонов; в то время как некоторые эндокринные механизмы модулируются на секреторном уровне, другие модулируются на уровне числа рецепторов или чувствительности (Bamberger et al., 1996 г. ). Также очевидны временные вариации нейроэндокринных механизмов. Например, у некоторых птиц агрессия (например, защита территории) во время брачного сезона регулируется циркулирующим тестостероном, в то время как территориальность в период отсутствия размножения регулируется локализованным (т. Е. Не циркулирующим) производством тестостерона и эстрадиола, вырабатываемого внутри мозг (Soma et al., 2008 ).
(2) Сходимость . Разнообразие механизмов, связанных с социальным поведением, также увеличивает возможность эволюционного сближения в гормональных и других ближайших решениях поведенческих проблем.В качестве потенциального примера такой конвергенции, звуковые сигналы птиц и рыб — оба участвуют в защите территории — затрагивают очень разные органы, производящие звук (сиринкс, плавательный пузырь, электрические органы), но обе системы модулируются андрогенами и регулируются различными, но одинаково функционирующими нервными цепями (Smith et al., 1997 ; Закон и Данлэп, 1999 ; Басс и др., 2008 ).
(3) Сохранение функции . Особенно эффективные механизмы могут сохраняться с течением времени и в результате событий видообразования из-за адаптивных преимуществ, которые они дают.Например, окситоцин и вазопрессин (или их гомологи) участвуют в поддержании аффилиативных отношений у певчих птиц (Goodson et al., 2009 ) и у полевок (Donaldson, Young, 2008 ). Точно так же глюкокортикоиды играют общую роль в мобилизации энергии и перенаправлении поведения у многих позвоночных и во многих различных социальных контекстах (Denver, 2009). ). Эти наблюдения предполагают, что эти эндокринные системы имеют глубокую историю эволюции и что их функции сохраняются в течение длительных периодов эволюции позвоночных.
В общем, эти закономерности возникнут только при сравнительном эволюционном подходе. Однако для проведения таких сравнений требуется больше информации о механизмах социального поведения по разным таксонам. В настоящее время у нас нет доказательств механизмов, лежащих в основе социального поведения от достаточного количества таксонов — до тех пор, пока эта информация не будет обнаружена, такие сравнительные эволюционные подходы недостижимы. Таким образом, мы утверждаем, что поведенческие экологи и нейробиологи должны участвовать в более тесном сотрудничестве, направленном на определение механизмов социального поведения в новых модельных организмах, расширяя ограниченное количество традиционных модельных организмов, используемых на сегодняшний день.
Новое определение модельных организмов
Естественный отбор породил необычайное разнообразие поведенческих адаптаций к изменчивости окружающей среды. Поведенческие экологи использовали это разнообразие для определения экологических причин и последствий изменения социального поведения у широкого круга видов. Однако наше понимание генетической и нейроэндокринной основы социального поведения все еще ограничено и было получено в основном из лабораторных исследований инбредных линий классических модельных организмов (например,g., мыши, крысы и плодовые мухи; Соколовский, 2010 г. ). В самом деле, могут быть немодельные системы, которые могут быть лучшими моделями, чем эти классические модельные организмы. Чтобы начать использовать нашу интегративную модель для всестороннего понимания социальных взаимодействий, пора переосмыслить традиционное определение модельного организма и начать разработку новых модельных систем, в которых также можно изучать вопросы функций конечного уровня. как нейробиологические и генетические механизмы ближайшего уровня, лежащие в основе социальной изменчивости.
Идеальная модельная система должна демонстрировать заметные пространственные или временные вариации социального поведения, которые можно изучать количественно как в лаборатории, так и в естественной (или полуестественной) среде. Традиционно черты, которые делали организм ценным для лабораторных исследований, включали короткое время генерации, регулярное размножение в лабораторных условиях, простоту использования для экспериментальных манипуляций и доступность инструментов для характеристики генетических, нервных и эндокринных механизмов.Тем не менее, некоторые свободноживущие виды могут иллюстрировать способы распространения человеческих популяций по всему миру, и, следовательно, они могут предоставить идеальную модель для анализа социальных вариаций и поведенческих механизмов, которые уникальным образом сформированы этой историей расселения и последующей адаптацией к различным экологическим ситуациям. . По мере совершенствования генетических и других технологий увеличивается возможность изучения механизмов у менее традиционных модельных видов (Donaldson and Young, 2008 г. ; МакГроу и Янг, 2010 г. ).Другие желательные качества, такие как простота отлова и наблюдения в дикой природе, будут гарантировать, что по мере того, как эти технологии становятся все более доступными и доступными.
Релевантность для социального поведения человека
Предлагаемый нами интегративный подход в сочетании с разработкой новых модельных организмов для изучения различий в социальном поведении может предоставить мощные средства для перевода результатов исследований в более глубокое понимание социальной когнитивной функции человека (Ebstein et al., 2010 ; Инсель, 2010 г. ).Различные психоневрологические расстройства характеризуются нарушениями социального поведения и социального познания, включая депрессию, расстройства аутистического спектра, биполярные расстройства, обсессивно-компульсивные расстройства и шизофрению. Следовательно, интегративные исследования с использованием более точных моделей социального поведения на животных могут привести к разработке стратегий лечения психопатологий. Другими словами, перспективы поведенческой экологии могут привести к открытиям моделей на животных, которые идеально подходят для решения вопросов, относящихся к психиатрии.Например, открытие роли окситоцина в социальной привязанности у социально моногамных степных полевок уже привело к клиническим испытаниям терапии для улучшения социальной когнитивной функции при расстройствах аутистического спектра за счет использования фармакотерапии окситоцином (Opar, 2008). ). Таким образом, принимая более экологический и эволюционный подход к биомедицинским исследованиям, мы должны улучшить наши биомедицинские открытия.
Выводы
Наша интеграционная структура (Рисунок 1 ) имеет широкое значение для изучения социального поведения.Полностью интегративная модель социальности была «святым Граалем» поведенческих экологов, изучающих социальные отношения между людьми. Проводя исследования с явными связями между ближайшим и конечным уровнями анализа, мы получаем всеобъемлющее и комплексное понимание этих отношений, что может привести к сдвигу парадигмы при изучении изучения социального поведения. Кроме того, у этого подхода есть много приложений. Во-первых, интегративный взгляд на социальность, включающий генетические, нейронные и эндокринные механизмы, даст представление об эволюционном потенциале и, таким образом, предоставит мощный инструмент для прогнозирования реакции видов на изменение окружающей среды.Во-вторых, такое фундаментально интегративное понимание может дать понимание психопатологических расстройств человека, что может улучшить лечение этих состояний. В-третьих, понимание связи между экологической изменчивостью, механизмом и социальностью может привести к улучшениям в животноводстве и благополучии как части управления исчезающими видами, так и животноводства. Чтобы добиться этих преимуществ, мы должны обучить наше следующее поколение студентов интегративным подходам к поведению и создать культуру междисциплинарной совместной работы.Это требует финансовой и материально-технической поддержки со стороны университетов, медицинских учреждений и финансовых агентств. Хотя эти усилия и трудны, они принесут исключительную пользу для изучения социального поведения.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Эта рукопись является продуктом международной рабочей группы, которая собралась 4–8 августа 2009 г. в Папском католическом университете Чили в Сантьяго, Чили.Рабочая группа финансировалась международным грантом Национального научного фонда Дэниелу Т. Блюмштейну (# 05) и Лорен Д. Хейс (# 0
6), грантом Departamento de Relaciones Internacionales de CONICYT Луису А. Эбенспергеру и MIDEPLAN ICM. -P05-002 и PFB-23-CONICYT-IEB — средства Родриго А. Васкеса. Кроме того, СИНЕСТАВ поддержал Габриэлу Гонсалес-Марискаль. После четырех организаторов семинара в алфавитном порядке перечислены авторы.Список литературы
Александр, Р.Д. (1974). Эволюция социального поведения. Annu. Rev. Ecol. Syst. 5, 325–383.
CrossRef Полный текст
Александр, С. Л., и Ирвин, К. Х. Г. (1998). Влияние социального стресса на активность оси надпочечников у лошадей: важность мониторинга способности связывать кортикостероиды глобулина. J. Endocrinol. 157, 425–432.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Бамбергер, К.М., Шульте, Х. М., и Хрусос, Г. П. (1996). Молекулярные детерминанты функции рецепторов глюкокортикоидов и чувствительности тканей к глюкокортикоидам. Endocr. Ред. 17, 245–261.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Бартц, Дж. А., и Холландер, Э. (2006). Нейробиология принадлежности: налаживание связей между фундаментальными и клиническими исследованиями нейропептидов и социального поведения. Horm.Behav. 50, 518–528.
Pubmed Аннотация | Pubmed Полный текст
Бири А. К., Лейси Е. А. и Фрэнсис Д. Д. (2008). Распределение рецепторов окситоцина и вазопрессина у одиночных и социальных видов туко-туко ( Ctenomys haigi и Ctenomys sociabilis ). J. Comp. Neurol. 507, 1847–1859.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Болхуис, Дж.Дж., И Ферхюльст, С. (2009). Наследие Тинбергена: функции и механизмы в поведенческой биологии . Нью-Йорк: Издательство Кембриджского университета.
CrossRef Полный текст
Босх, О. Дж., Наир, Х. П., Ахерн, Т. Д., Нойман, И. Д., Янг, Л. Дж. (2009). Система CRF опосредует возникновение депрессивного поведения после потери партнера у моногамного грызуна. Нейропсихофармакология 34, 1406–1415.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Брашарис, Дж.С., и Арсезе, П. (2002). Роль корма, среды обитания и хищничества в пластичности поведения маленькой африканской антилопы. J. Anim. Ecol. 71, 628–638.
CrossRef Полный текст
Бунтин, Дж. Д. (1996). Нервно-гормональный контроль родительского поведения у птиц. Adv. Изучите поведение. 25, 161–213.
CrossRef Полный текст
Кахан, С. Х., Блюмштейн, Д. Т., Сундстрём, Л., Либих, Дж., и Гриффин, А. (2002). Социальные траектории и эволюция социального поведения. Oikos 96, 206–216.
CrossRef Полный текст
От кутюр, С. М., Пенн, Д. Л., Лош, М., Адольф, Р., Херли, Р., и Пивен, Дж. (2010). Сравнение социальных когнитивных функций при шизофрении и высокофункциональном аутизме: больше конвергенции, чем расхождения. Psychol. Med. 40, 569–579.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Крил, С.(2001). Социальное доминирование и гормоны стресса. Trends Ecol. Evol. 16, 491–497.
CrossRef Полный текст
Каммингс, М. Э., Ларкинс-Форд, Дж., Рейли, К. Р. Л., Вонг, Р. Ю., Рэмси, М., и Хофманн, Х. А. (2008). Сексуальные и социальные стимулы вызывают быстрые и контрастные геномные реакции. Proc. R. Soc. Лонд., Б, Биол. Sci. 275, 393–402.
CrossRef Полный текст
Девиче, П., Брейнер, К., и Орчиник, М. (2001). Тестостерон, кортикостерон и фотопериод взаимодействуют, регулируя уровни связывающего глобулина и свободного стероидного гормона в плазме у Темноглазых Juncos, Junco hyemalis . Gen. Comp. Эндокринол. 122, 67–77.
CrossRef Полный текст
Дональдсон, З. Р., Янг, С. Х., Чан, А. У. и Янг, Л. Дж. (2009). Получение зародышевой линии трансгенных степных полевок ( Microtus ochrogaster ) с использованием лентивирусных векторов. Biol. Репрод. 81, 1189–1195.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Эбенспергер, Л. А. (2001). Обзор эволюционных причин группового проживания грызунов. Acta Theriol. 46, 115–144.
CrossRef Полный текст
Эбштейн, Р. П., Израиль, С., Чу, С. Х., Чжун, С., и Кнафо, А. (2010). Генетика социального поведения человека. Нейрон 65, 831–844.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Федер М. Э., Беннетт А. Ф. и Хьюи Р. Б. (2000). Эволюционная психология. Annu. Rev. Ecol. Syst. 31, 315–341.
CrossRef Полный текст
Фидлер, А. Э., ван Оерс, К., Дрент, П. Дж., Кун, С., Мюллер, Дж. К., и Кемпенаерс, Б. (2007). Полиморфизм гена Drd4 связан с изменчивостью личности воробьиных птиц. Proc. R. Soc. Лонд., Б, Биол. Sci. 274, 1685–1691.
CrossRef Полный текст
Гудсон, Дж. Л., Шрок, С. Е., Клатт, Дж. Д., Кабелик, Д., и Кингсбери, М. А. (2009). Рецепторы мезотоцина и нонапептида способствуют формированию скопления эстрилидов. Наука 325, 862–866.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Гриффин, А.С., Уэст, С.А. (2002). Родственный отбор: факт и вымысел. Trends Ecol. Evol. 17, 15–21.
CrossRef Полный текст
Хэслер, С., Рошфор, К., Георги, Б., Личнерски, П., Остен, П., и Шарфф, К. (2007). Неполная и неточная имитация голоса после нокдауна FoxP2 в области ядра базальных ганглиев певчих птиц X. PLoS Biol. 5, е321. DOI: 10.1371 / journal.pbio.0050321.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Гермес, Г.Л., Дельгадо, Б., Третьякова, М., Кавигелли, С. А., Краус, Т., Концен, С. Д., и Мак-Клинток, М. К. (2009). Социальная изоляция нарушает регуляцию эндокринного и поведенческого стресса, увеличивая при этом злокачественное бремя спонтанных опухолей молочной железы. Proc. Natl. Акад. Sci. США 106, 22393–22398.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Холмс У. Г. и Шерман П. У. (1982). Онтогенез родства у двух видов сусликов. Am . Зоолог 22, 491–517.
CrossRef Полный текст
Хосеманн, К. Э., Колозимо, П. Ф., Саммерс, Б. Р. и Кингсли, Д. М. (2004). Простой и эффективный протокол микроинъекций для получения трансгенной колюшки. Поведение 141, 1345–1355.
CrossRef Полный текст
Джонстон, Р. А. (2000). Модели репродуктивного перекоса: обзор и обобщение. Этология 106, 5–26.
CrossRef Полный текст
Лим, М. М., Ван, З., Оласабаль, Д. Э., Рен, X., Тервиллигер, Э. Ф., Янг, Л. Дж. (2004). Повышенное предпочтение партнера у беспорядочных полов путем манипулирования экспрессией одного гена. Природа 429, 754–757.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Мельман, П. (1979). Помощники шакалов и выживание щенков. Природа 277, 382–383.
CrossRef Полный текст
Нуньес, С., Дунец, Т. Р., Швеппе, С. А., и Холекамп, К. Э. (1999). Энергетическое и эндокринное опосредование естественного рассредоточения у сусликов Белдинга. Horm. Behav. 35, 113–124.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Олазабаль Д. Э. и Янг Л. Дж. (2006a). Виды и индивидуальные различия в уходе за молодыми самками аллопатников связаны с плотностью рецепторов окситоцина в полосатом теле и боковой перегородке. Horm. Behav. 49, 681–687.
CrossRef Полный текст
Олазабаль Д. Э. и Янг Л. Дж. (2006b). Рецепторы окситоцина в прилежащем ядре способствуют «спонтанному» материнскому поведению у взрослых самок степных полевок. Неврология 141, 559–568.
CrossRef Полный текст
Пфафф, Д. (2005). Гормоны, мозг и поведение , Vol. 5. Бостон: Academic Press.
CrossRef Полный текст
Ромеро, Л.М. (2002). Сезонные изменения концентрации глюкокортикоидов в плазме у свободноживущих позвоночных. Gen. Comp. Эндокринол. 128, 1–24.
Pubmed Аннотация | Pubmed Полный текст
Росс Х. Э., Фриман С. М., Шпигель Л. Л., Рен Х., Тервиллигер Э. Ф. и Янг Л. Дж. (2009). Вариация плотности рецепторов окситоцина в прилежащем ядре оказывает различное влияние на аффилиативное поведение у моногамных и полигамных полевок. Дж.Neurosci. 29, 1312–1318.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Рубинштейн, Д. Р. (2007). Гормоны стресса и социальность: интеграция социальных и экологических стрессоров. Proc. R. Soc. Лонд., Б, Биол. Sci. 274, 967–975.
CrossRef Полный текст
Сасаки, Э., Суэмизу, Х., Шимада, А., Ханадзава, К., Оива, Р., Камиока, М., Томиока, И., Сотомару, Ю., Хиракава, Р., Это, Т., Сиодзава, С., Маэда, Т., Ито, М., Ито, Р., Кито, К., Ягихаси, К., Кавай, К. , Миёси, Х., Таниока, Ю., Тамаоки, Н., Хабу, С., Окано, Х., и Номура, Т. (2009). Получение трансгенных приматов, отличных от человека, с передачей по зародышевой линии. Природа 459, 523–527.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Шох, С. Дж., Рейнольдс, С. Дж., И Боутон, Р. К. (2004).«Эндокринология», в Экология и эволюция кооперативного разведения птиц, , ред. У. Д. Кениг и Дж. Дикенсон (Кембридж: издательство Кембриджского университета), 128–141.
CrossRef Полный текст
Шрадин, К., Скантлбери, М., Пиллэй, Н., Кониг, Б. (2009). Уровни тестостерона у доминирующих общительных самцов ниже, чем у одиночных бродяг: физиологическая разница между тремя репродуктивными тактиками самцов у социально гибкого млекопитающего. Am.Nat. 173, 376–388.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Шерман, П. В. (1988). Уровни анализа. Anim. Behav. 36, 616–619.
CrossRef Полный текст
Смит, Г. Т., Бреновиц, Э. А., Бичер, М. Д., и Вингфилд, Дж. К. (1997). Сезонные изменения тестостерона, нейронных атрибутов ядер, управляющих песней, и структуры песни у диких певчих птиц. J. Neurosci. 17, 6001–6010.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Сома, К. К., Скотти, М. А. Л., Ньюман, А. Е. М., Шарлье, Т. Д., и Демас, Г. Е. (2008). Новые механизмы нейроэндокринной регуляции агрессии. Фронт. Нейроэндокринол. 29, 476–489.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Спенсер, Р.Л., Миллер, А. Х., Модей, Х., МакИвен, Б. С., Бланшар, Р. Дж., Бланшар, Д. К., и Сакаи, Р. Р. (1996). Хронический социальный стресс приводит к снижению уровня связывания рецепторов кортикостероидов типа II в селезенке и уровней глобулина, связывающего кортикостероиды в плазме крови. Психонейроэндокринология 21, 95–109.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Марки, Дж. А. (1991). Почему проблемы эволюции возрождают интерес к ближайшим поведенческим механизмам. Am. Zool. 31, 338–348.
CrossRef Полный текст
Тинберген, Н. (1963). О целях и методах этологии. Z. Tierpsychol. 20, 410–433.
CrossRef Полный текст
Uchino, B.N., Cacioppo, J.T., и Kiecolt-Glaser, J.K. (1996). Взаимосвязь между социальной поддержкой и физиологическими процессами: обзор с упором на основные механизмы и последствия для здоровья. Psychol. Бык. 119, 488–531.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Уилсон, Э. О. (1975). Социобиология: новый синтез . Кембридж: Belknap Press издательства Гарвардского университета.
CrossRef Полный текст
Вингфилд, Дж. К., Хегнер, Р. Э., Дафти, А. М. младший, и Болл, Г. Ф. (1990). «Гипотеза вызова»: теоретические последствия для моделей секреции тестостерона, систем спаривания и стратегий разведения. Am. Nat. 136, 829–846.
CrossRef Полный текст
Янг А.Дж., Карлсон А.А., Монфорт С.Л., Рассел А.Ф., Беннетт Н.С. и Клаттон-Брок Т. (2006). Стресс и подавление подчиненного воспроизводства у сурикатов, выращивающих совместно. Proc. Natl. Акад. Sci. США 103, 12005–12010.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Зисук, С.и Шухтер С. Р. (1991). Депрессия в течение первого года после смерти супруга. Am. J. Psychiatry 148, 1346–1352.
Pubmed Аннотация | Pubmed Полный текст
Привыкание — обзор | Темы ScienceDirect
II.A. Модели привыкания и исчезновения
Процессы, часто упоминаемые в качестве объяснения эффектов воздействия, — это привыкание и исчезновение.
Привыкание относится к снижению реакций страха, особенно физиологических реакций, на многократное воздействие провоцирующих страх раздражителей.Привыкание рассматривается как невыученная временная реакция. Предполагается, что познания не влияют на привыкание. Классическая теория привыкания предсказывает, что привыкание не произошло бы, если бы (исходное) возбуждение было высоким. Затем возбуждение будет еще больше усиливаться и приводить к сенсибилизации (т. Е. Усилению реакции страха на повторяющиеся воздействия на провоцирующие страх стимулы). Однако в литературе показано, что, например, во время воздействия сильных раздражителей, вызывающих страх (например, терапия наводнения), можно было ожидать снижения, а не дальнейшего увеличения психофизиологической и субъективной тревожности.Недавние теории привыкания были расширены с учетом этих результатов. Эти теории двойного процесса описывают сложные взаимодействия между привыканием и сенсибилизацией, при которых привыкание может в конечном итоге произойти после воздействия сильных раздражителей, вызывающих страх.
Несколько исследований предоставили подтверждающие доказательства роли привыкания в экспозиционной терапии. Во-первых, в нескольких исследованиях страх и физиологическое возбуждение, о которых сообщают сами люди, демонстрируют тенденцию к снижению в зависимости от воздействия, согласующегося с привыканием.Во-вторых, исследования конкретных фобий показали, что физиологическое привыкание связано с улучшением, в то время как отказ от привычки — нет. В-третьих, переменные, не зависящие от привыкания (например, уровень возбуждения, скорость стимуляции и регулярность предъявления), похоже, одинаковым образом влияют на привыкание и снижение страха.
Основная трудность теорий привыкания — это предполагаемый краткосрочный эффект привыкания. Теория привыкания предсказывает возврат страха через определенный промежуток времени без воздействия стимулов.Исследования, однако, показывают, что значительная часть пациентов сообщают, что не испытывают беспокойства в течение длительных периодов времени. Чтобы объяснить долгосрочные эффекты воздействия, некоторые теоретики проводят различие между краткосрочным и долгосрочным привыканием. Например, было высказано предположение, что долгосрочное привыкание зависит от высших когнитивных процессов, тогда как краткосрочное привыкание представляет собой в основном вегетативное демпфирование. У теории привыкания больше ограничений. Например, модель привыкания не может адекватно объяснить стойкость некоторых пугающих реакций, несмотря на неоднократные воздействия.
Угасание относится к уменьшению реакции посредством повторения неусиленной реакции. Таким образом, теории, основанные на исчезновении, предполагают, что снижение тревожности происходит в результате повторных встреч с провоцирующими тревогу ситуациями без отталкивающих последствий. Предполагается, что в отличие от привыкания, на вымирание влияют познания. Другими словами, предполагается, что это активный, а не пассивный процесс. Теория вымирания предполагает, что фобическому поведению научились и поэтому от него можно отказаться.Было предложено несколько когнитивных объяснений тому, чему научились во время угасания фобического поведения. Например, постулируется, что изучается неподтверждение ожиданий результата, или повышенная самоэффективность, или тот факт, что возбуждение, связанное с воздействием, не опасно. Эти когнитивно-ориентированные теории будут обсуждаться более подробно в следующих параграфах.
Еще одно объяснение состоит в том, что контекст определяет, какое значение приобретает ситуация.Было высказано предположение, что совместное присутствие стимула (например, сердцебиение) и контекста (например, вдали от дома) определяют значение (например, сердечный приступ). Последняя теория согласуется с доказательствами того, что возвращение страха можно свести к минимуму с помощью длительных сеансов воздействия в различных контекстах.
Доказательства, противоречащие теории вымирания при объяснении уменьшения страха и тревоги во время воздействия, получены из исследований «побега». Согласно теории вымирания, пациенты должны оставаться в провоцирующей страх ситуации до тех пор, пока тревога существенно не уменьшится, чтобы понять, что ничего плохого не происходит.Однако несколько исследований продемонстрировали, что снижение тревожности может происходить, даже если пациентам позволяли вырваться из пугающей ситуации до достижения максимальной тревожности. В этих исследованиях неясно, что изучается. Похоже, что у пациентов не было времени и шанса узнать о возможных последствиях, которых они боялись.
Как поведение меняет мозг? Множественные методы ответа на старые вопросы1 | Интегративная и сравнительная биология
Аннотация
Очевидно, что мозг контролирует поведение, но может ли поведение также «управлять» мозгом? В эволюционном масштабе времени выборочное экологическое давление формирует сенсорные и двигательные способности, а также тело и поведение.Соответственно, в процессе развития поведение действует согласованно с окружающей средой, вызывая структурные изменения в мозге на всю жизнь. Удивительно, но в «реальном времени» социальное поведение также может вызывать изменения, обычно обратимые, в мозге взрослых животных. Изменения, вызванные поведенческими взаимодействиями, могут быть драматичными, и во многих случаях эти взаимодействия напрямую связаны с репродуктивным поведением. Понимание того, как поведение формирует мозг в ходе поведенческих взаимодействий, является серьезной проблемой.Для анализа таких изменений необходима модельная система, позволяющая управлять биологической и поведенческой средой многих животных одновременно, но позволяющая регулировать физиологические, клеточные и молекулярные процессы. Цихлида Haplochromis (Astatotilapia) burtoni (Günther) из озера Танганьика, вынашивающая ротовую полость, пригодна для изучения социальных влияний на мозг. У него сложное, но легко наблюдаемое индивидуальное и социальное поведение, регулируемое двумя различными классами мужчин: с территориями и без.Многие особенности животных сформированы социальными контактами, включая созревание молодых особей, гипоталамо-гипофизарно-гонадную ось, скорость роста, уровень базального стресса и другие. Как социальная информация влияет на изменение мозга и тела? Животные должны проявлять внимание к общественной жизни, чтобы определить свои шансы. Изучение того, как социальная информация преобразуется в клеточные изменения у этого вида, должно помочь понять, как это происходит у других социальных животных.
Фон Икскюлл (1921) понял, что каждый вид переживает жизнь по-своему, потому что он живет в своем собственном особом «умвельте», или уникальном мире восприятия.Например, летучая мышь использует звуковое эхо, чтобы исследовать темноту, формируя изображения из звука, отраженного от окружающей среды, но также может видеть своими глазами в ультрафиолете, возможно, для добычи нектара (Winter et al. , 2003), давая ей два уникальных окна в мире. Такие сенсорные способности определяют, что можно почувствовать, позволяя одни и ограничивая другие поведенческие реакции животного. Писавший в начале двадцатого века, фон Икскюлл не мог предвидеть открытия магнитных, электрических или чувствительных ощущений, а также не мог представить себе «визуальное» ощущение, простирающееся от инфракрасного до ультрафиолетового.Лоренц (1932) расширил идею «умвельта», включив в нее не только обнаружение стимулов от физического окружения, но и от других животных. Его влиятельная статья «Компаньоны как факторы среды обитания птиц» показала, что бихевиористам необходимо расширить свои взгляды на мир восприятия животных, включив в него других людей и их совокупный социальный контекст. Поведение — главный судья выживания животных и того, как животные реагируют во время взаимодействия с другими, а окружающая среда формирует его фенотип.Однако поведение, в свою очередь, зависит от сложных физиологических, клеточных и, в конечном итоге, молекулярных адаптаций, выработанных в процессе эволюции.
Основная задача биологии состоит в том, чтобы обнаружить, как поведение контролируется с помощью физиологических процессов и, соответственно, как поведение влияет на физиологические, клеточные или молекулярные события. Несмотря на амбициозность, эта цель становится все более реалистичной благодаря множеству новых методов, появившихся в результате достижений клеточной и молекулярной биологии. Окончательные вопросы об эволюции и контроле поведения требуют понимания причинных механизмов у животных, когда они взаимодействуют друг с другом, предпочтительно в достаточно естественной обстановке.Тем не менее, подавляющее большинство экспериментов проводится на изолированных особях, многие из которых являются одомашненными видами. Похоже, что мало что можно узнать о том, как эволюция повлияла на социальное поведение, анализируя отдельных животных, поскольку социальные взаимодействия невозможны. В этом обзоре я опишу результаты нашей исследовательской программы с использованием нескольких методов для изучения людей в полуестественном социальном контексте. Эксперименты направлены на обнаружение того, как социальный контекст репродуктивного поведения формирует мозг и, в свою очередь, изменяет поведение животных во время их взаимодействия.
Ранние этологи вышли за рамки описательного анализа, предоставив основу для понимания порядка, лежащего в основе поведения животных. Изучая важные жизненные события, такие как кормление и размножение у видов с менее сложными поведенческими взаимодействиями, ранние этологи Конрад Лоренц (1981) и Нико Тинберген (1951) определили центральные принципы классической этологии и привели критическое рациональное обсуждение в понимание поведения. Открытия того, как животные реагируют на стимулы от сородичей, предоставили важную информацию о ближайших факторах, ответственных за то, как животные ведут себя так, как они себя ведут.Однако эти элементарные компоненты поведения можно понять только в естественном контексте животного. А в реальной жизни животные ведут себя и постоянно взаимодействуют, обеспечивая неразрывную взаимосвязь между тем, что они видят, и тем, что они делают. Учитывая важность сложных социальных взаимодействий, ученые искали модельные системы для исследования социальных взаимодействий.
В 1950 году Бэрэндс и Баэрэндс ван Роон опубликовали знаменательную монографию, описывающую поведение многочисленных видов рыб-цихлид.В этой публикации описываются атрибуты этих костистых рыб и утверждается, что эти животные хорошо подходят для новой на тот момент области этологии. Во-первых, в разумных пределах, цихлид можно изучать в лаборатории без ущерба для их естественного поведенческого контекста или экологической достоверности результатов. Во-вторых, цихлиды активны, что делает сбор количественных данных реальной целью. В-третьих, существует множество видов цихлид, которые эволюционировали, сталкиваясь с различными ограничениями окружающей среды, что делает сравнение близкородственных видов возможной стратегией для выявления потенциальных селективных сил.В самом деле, цихлиды сыграли важную роль в понимании скорости эволюции (Verheyen et al. , 2003) и в обнаружении того, как условия окружающей среды могут поставить под угрозу половой отбор (Seehausen et al. , 1997). Таким образом, цихлиды предоставили уникальные возможности для изучения как ближайших механизмов, так и конечных функций поведения животных в одной и той же модельной системе. Безусловно, для анализа аспектов социального поведения с успехом использовались и другие системы моделей рыб, и действительно, есть заявления о ряде довольно замечательных наборов поведенческих адаптаций ( e.g., Bshary et al. , 2002). Несколько видов рыб были полезны для анализа звуковой коммуникации (Bass et al. , 2000), для определения роли поведения в смене пола (, например, Godwin et al. , 2003) и социальной модуляции андрогенов. уровни были изучены в костистых костях (, например, , Oliveria, и др., , 1996).
Социальная система африканских цихлид, Haplochromis (Astatotilapia) burtoni (Günther), включает два типа взрослых самцов: с территориями и без территорий (Fernald, 1977).Территориальные (Т) самцы ярко окрашены, с синей или желтой основной окраской тела, с черной полосой через глаз (слезный), вертикальными черными полосами на теле от жаберной крышки до хвоста, черным пятном на кончике жабр. крышка и большая красная плечевая заплатка сразу за ней. Напротив, нетерриториальные (NT) самцы имеют загадочную окраску, из-за чего их трудно отличить от фона и от самок, которые так же замаскированы (рис. 1). Социальное общение между этими рыбами, по-видимому, зависит в первую очередь от визуальных сигналов (Fernald, 1984; см. Ниже).Хотя было показано, что другие виды цихлид продуцируют определенные стероидные сигналы (, например, Oliveira et al. , 1996), мы не обнаружили такие химические сигналы у H. burtoni (Robison et al. , 1998). . Анализ их естественной среды обитания, мелководных прибрежных бассейнов и устьев рек озера Танганьика (Coulter, 1991), H. burtoni живут в социальной системе, в которой T-самцы энергично защищают прилегающие территории (Fernald and Hirata, 1977, a , б ).
Территориальные самцы H. burtoni очень активны, демонстрируя 19 различных поведенческих паттернов в социальных взаимодействиях (Fernald, 1977). Территориальные самцы роют ямы на своей территории, обмениваются демонстрациями угрозы с соседними территориальными самцами, преследуют NT животных со своей территории, а также ухаживают за самками и ухаживают за ними. Ухаживая за самками и ухаживая за ними, самцы Т демонстрируют яркую окраску по отношению к самке, за которой ухаживают. Т-самец ведет самку к своей территории, обычно используя большие движения хвоста, и ухаживает, тряся своим открытым ярко окрашенным анальным плавником перед самкой.Когда T-самцу удается заманить самку на свою территорию, она обычно ест, просеивая субстрат на территории. Самцы NT будут достаточно хорошо имитировать поведение самок, так что самец T позволяет NT проникать на территорию и питаться до того, как обман будет обнаружен. Это поведение NT-самцов происходит потому, что пища есть только в тех участках, которые охраняются как территории, поэтому NT-самцы должны заходить туда, чтобы поесть. Однако обычно новозеландскую женщину-имитатора быстро прогоняют. Если самка отвечает на ухаживания самца, Т-самец приведет ее в свою яму и продолжит ухаживания.Самцы T энергично плавают перед самкой, дрожа всем телом расправленными анальными плавниками. При соответствующей стимуляции самка откладывает яйца в яму и сразу же собирает их в рот. После того, как она откладывает несколько яиц, самец будет плавать перед ней, показывая на своем анальном плавнике (глазках) яичные пятна. Самцы T имеют этот плавник, потому что самке пятна могут показаться еще не собранными яйцами (Wickler, 1962). Таким образом, пытаясь «собрать» яйцеклетки, самка поедает молоки, выброшенные рядом с ними самцом, и обеспечивает оплодотворение.Нерестящийся самец может неоднократно прерывать свои ухаживания и спаривание, чтобы отогнать злоумышленников на свою территорию. После нескольких периодов откладки яиц и оплодотворения самка улетает с оплодотворенными яйцами, которые она вынашивает во рту (Fernald, 1984).
Даже это сокращенное описание естественного поведения H. burtoni показывает важную роль, которую визуальные сигналы играют в опосредовании социального поведения. Как типично для социальных взаимодействий, каждый поведенческий акт влияет на следующий как у человека, так и у других животных, участвующих в столкновении.На что обращают внимание животные во время агрессивных социальных взаимодействий? Используя этологические методы, первые исследователи идентифицировали несколько фиксированных паттернов действий и ключевых стимулов, которые опосредуют передачу социальных сигналов у H. burtoni . В частности, Леонг (1969) проанализировал роль черной полосы под глазом, проверив, как Т-самцы отреагировали на манекенов H. burtoni , окрашенных в различные конфигурации характерных узоров тела. Когда глазная полоса была представлена отдельно, T-самцы увеличивали свою готовность к атаке, в то время как наличие только оранжево-красного пятна на плечевой чешуе уменьшало готовность к атаке.Последующие эксперименты проверяли важность ориентации перемычки для глаз относительно тела и других зрительных стимулов (Heiligenberg and Kramer, 1972; Heiligenberg et al. , 1972). Все исследования подтверждают мнение о том, что действие черной перемычки под глазами и красного плечевого пятна влияет на агрессивность Т-самцов в противоположных направлениях. Самцы, выращенные после вылупления в полной изоляции, показали такую же реакцию на предъявление манекенов, что и нормальные животные, предполагая, что реакция на эти ключевые стимулы является врожденной (Fernald, 1980). H. burtoni Агрессивное поведение также оценивалось на предмет других сигналов, основанных на теории игр ( ср., Enquist and Leimar, 1983; Enquist and Jakobsson, 1986).
У H. burtoni зрительная система имеет весьма заметную адаптацию к поведенческим сигналам вида. В основной среде обитания, в прибрежных бассейнах и устьях рек вдоль озера Танганьика, цветовые узоры на теле соответствуют фильтрующим свойствам воды, что обеспечивает максимальную видимость важнейших визуальных сигналов (Fernald and Hirata, 1977, a ).Сетчатка глаза H. burtoni имеет три типа колбочковых фоторецепторов и тип стержня, характерно чувствительный к разным длинам волн света, что означает, что у них может быть трехцветное зрение (Fernald and Liebman, 1980). Фоторецепторы колбочек расположены в квадратном массиве, который оптимален для цветового зрения (Fernald, 1981), а спектральная чувствительность, измеренная поведенческими методами (Allen and Fernald, 1985), показывает, что H. burtoni может различать цвета, как это было предсказано на основе морфологических измерений.Поскольку глаз продолжает расти за счет добавления новых нейронов, зрительная система H. burtoni оказалась полезной для понимания того, как контролируется развитие сетчатки (, например, Fernald, 2000, a , b ).
Одной из самых замечательных особенностей позвоночных с неопределенным ростом является то, как постоянные сенсорные и моторные функции поддерживаются при изменении размера тела. Например, рост глаза у H. burtoni настолько быстр, что тело только что выпущенного малька могло поместиться в глазу годовалого самца T. (Zygar et al., 1999). Рост достигается за счет добавления новых клеток в хрусталик и сетчатку без ущерба для зрения (Johns and Fernald, 1981; Fernald and Wright, 1983; Fernald, 1983, 1989). Наблюдая за животными, становится очевидным, что скорость роста неоднородна и критически зависит от социальной ситуации (Fraley and Fernald, 1982). Сообщалось о скорости роста в зависимости от социальной ситуации для других видов рыб (, например, Borowsky, 1973; Francis, 1988; Berglund, 1991; Schultz et al., 1991), но механизмы, с помощью которых осуществляется такой контроль, не поняты. Используя социальную систему H. burtoni , мы начинаем открывать механизмы, с помощью которых социальное поведение может регулировать физиологию H. burtoni .
После выпуска молодняка матерью H. burtoni рост, поведение и развитие гонад критически зависят от социальной среды (Fraley and Fernald, 1982). Выращивание животных, физически изолированных при визуальном контакте или в группах сородичей, не показало различий в росте на основе стандартной длины и веса в течение первых 10 недель (рис.2). Выращенные в группе самцы, которые стали NT, набирают меньше веса, чем те, которые становятся T, хотя эта разница больше не очевидна через 20 недель (рис. 2A). Гонады также развиваются быстрее у T-самцов, чем у NT-самцов, хотя и медленнее, чем у изолированных самцов в 14 недель (Fig. 2A). Физически изолированные самцы фактически становятся Т-самцами и демонстрируют все виды поведения, связанные с этим статусом. Возможно, у них развиваются большие размеры и гонады, потому что они не сталкиваются с реальной физической конкуренцией. При сравнении проявления поведенческих признаков, выращенные в группе самцы T демонстрируют характерные агонистические манеры поведения (погоня, взмах хвоста, раскрытие плавников) и окраску (глазная полоса, глазное пятно) более чем за две недели до животных, выращенных в условиях физической изоляции (рис.3). Обратите внимание, что это агрессивное поведение полностью подавляется у NT-самцов, выращенных в группе. В социальной системе H. burtoni , где территориальное пространство является ограничивающим фактором, такое надежное регулирование созревания на раннем этапе развития кажется адаптивным решением для ограниченных ресурсов.
Очевидно, что выращивание с одноклассниками может подавить раннее социальное и физическое развитие, но у H. burtoni может происходить даже более эффективное социальное регулирование, когда более старые животные содержатся с более молодыми животными.Дэвис и Ферналд (1990) выращивали животных после вылупления в присутствии взрослых самцов и показали, что эти рыбы подавляли созревание гонад по сравнению с рыбами, выращенными без взрослых особей (рис. 4). Этот эксперимент показал, что у подавленных животных были не только гипогонадные яички, но и меньшие нейроны, содержащие гонадотропин-рилизинг-гормон (GnRH), в преоптической области. Нейроны GnRH являются ключевой точкой в оси мозг-гипофиз-гонады, которая контролирует воспроизводство у всех позвоночных. В г.burtoni , как и у всех позвоночных, нейроны GnRH проецируются в гипофиз (Bushnik and Fernald, 1995), где они высвобождают GnRH, сигнальный пептид, посылаемый из мозга в гипофиз, чтобы вызвать высвобождение гонадотропинов и, в конечном итоге, рост яичек. Davis и Fernald (1990) показали, что GnRH-содержащие клетки в головном мозге в 8 раз больше по T, чем у NT мужчин. Таким образом, социальный контроль созревания H. burtoni осуществляется путем изменения GnRH-содержащих клеток в головном мозге.
Социальный статус может регулировать физиологию репродуктивного состояния даже у взрослых H. burtoni , как показано при переключении самцов с T → NT или NT → T, перемещая их в новые сообщества. В частности, когда T-самцы были перемещены в сообщества с более крупными T-самцами, они стали NT (, например, T → NT), и аналогично NT-самцы были перемещены в сообщества с более мелкими сородичами, они стали T (, например, NT → T) . Через четыре недели в изменившейся социальной обстановке был измерен размер клеток ГнРГ (рис.5), показывающий, что одного лишь изменения социального статуса было достаточно, чтобы изменить размер нейрона ГнРГ в головном мозге. Как и ожидалось, соответственно изменился гонадосоматический индекс (GSI) (Francis et al. , 1993). Таким образом, как взрослые, так и подростки подвергаются социальному контролю над воспроизводством через изменения в нейронах гонадолиберин в головном мозге.
Хотя изменение структуры мозга путем изменения социального статуса весьма примечательно, временная шкала этого первоначального эксперимента не отражала, насколько быстро могут произойти поведенческие и нейронные изменения.Действительно, испытанный четырехнедельный интервал был значительно больше, чем любые наблюдаемые изменения в поведении после переключения статуса, которые могут происходить за считанные минуты. Анализ социально-индуцированных изменений нейронных структур в значительно более коротком временном масштабе выявил еще один сюрприз.
Используя парадигму изменения социального статуса путем перемещения животных, аналогичную описанной выше, White et al. (2002) обнаружил несколько важных новых аспектов социального контроля репродуктивной оси (рис.6). Во-первых, при социальном восхождении от NT к T-статусу, изменение размера клеток было довольно быстрым со значительным ростом за один день, а размер T-клеток мужского GnRH был достигнут за одну неделю. Нейроны GnRH фактически продолжали расти еще больше, так что через две недели они были значительно больше, чем нормальный размер Т-самца, прежде чем вернуться к размеру, подходящему для Т-самца (рис. 7). Это массивное усиление выработки ГнРГ, весьма вероятно, позволяет социально восходящему животному быстро достичь репродуктивной компетентности и, очевидно, не наблюдалось в 4-недельном эксперименте, описанном выше.Поведенческое переключение с NT на T, хотя сразу видно как изменение от неагрессивного к агрессивному поведению, не полностью соответствует таковому у стабильного T-самца для ок. 1,5 недели. Вторым открытием в этом эксперименте было то, что изменение между T и NT заметно асимметрично. Рыбы с нисходящим социальным статусом (T → NT) немедленно перестают проявлять агрессивное поведение, но нейроны, содержащие GnRH в POA, не уменьшаются до размера NT до ок. через три недели после поражения, тогда как восхождение NT → Τ занимает меньше недели.Значение этого гистерезиса в нервных и поведенческих изменениях между T- и NT-мужчинами можно объяснить как следствие жизни в нестабильном мире, где репродуктивные возможности могут возникнуть быстро для NT (см. Ниже). После поражения быстрое переключение на подчиненное поведение, вероятно, снижает шансы получить травму для проигравшего. Однако, учитывая, что шанс обосноваться на территории может появиться в ближайшее время, поддержание активной репродуктивной системы на более длительное время может быть адаптивным. Социальный статус определяет как размер сомы ГнРГ-ir нейронов POA, так и GSI, и эти эффекты обратимы.Относительно большие семенники и нейроны GnRH-ir, характерные для T-самцов, являются следствием их социального доминирования, и когда это преимущество доминирования утрачивается, и нейроны, и яички сокращаются, хотя, как показано здесь (White et al. , 2002), там Поразительна асимметрия физиологических реакций. Социальная информация о статусе вызывает изменения в мозгу, но как это достигается, неизвестно.
Белый et al. (2002) также показали, что социально-индуцированные изменения статуса приводят к значительным изменениям экспрессии генов.Измеряя изменения мРНК всех трех форм GnRH, они обнаружили, что только мРНК POA GnRH регулируется в соответствии с изменением социального статуса. Изменение мРНК в форме POA GnRH было очевидным через три дня после изменения социального статуса. Такое социальное регулирование демонстрирует, что ключевая социальная информация используется для управления клеточными и молекулярными процессами в мозге.
Важно отметить, что влияние социального статуса на размер клеток GnRH и экспрессию мРНК GnRH ограничивается GnRH-содержащими нейронами преоптической области.Как мы показали, H. burtoni имеет три различных гена, которые кодируют три различных GnRH-подобных молекулы (White et al. , 1994; White and Fernald, 1999), экспрессируемых в трех разных участках мозга (White и др., , 1995). Формы GnRH, не обнаруженные в POA, экспрессируются в двух других отдельных группах клеток, одна расположена в области терминального нерва, другая — в среднем мозге (подробности см. White et al. , 1995). Ни одна из этих других клеток, содержащих GnRH, не показала никаких изменений в размере в зависимости от социального статуса (Davis and Fernald, 1990), ни их мРНК не изменилась с изменением статуса.Таким образом, связанные со статусом вариации в размере сомы не являются общим свойством нейронов, содержащих GnRH, а, скорее, ограничены популяцией POA (Davis and Fernald, 1990). Тот же результат был показан для мРНК GnRH с использованием гибридизации in situ и (White et al. , 1995). Самцы и самки разделяют ось мозг-гипофиз-гонад, которая используется для контроля воспроизводства, но самка H. burtoni имеет совершенно другую систему, регулирующую размножение. ГнРГ-содержащие клетки в POA женщин также изменяют размер, но это происходит только в зависимости от их репродуктивного статуса (White and Fernald, 1993), и это не влияет на социальный статус.
Как и ожидалось, социальный контроль репродуктивной оси через гонадолиберин также влияет на важные эндокринные факторы. Андрогены, выделяемые гонадными железами, зависят от социального статуса. Кастрированные мужчины H. burtoni T имеют гипертрофированные нейроны ГнРГ (Фрэнсис и др. , 1992, a ; Сома и др. , 1996), показывая, что андроген оказывает обратное влияние на размер клеток ГнРГ (рис. 7) . Важным моментом является то, что уставкой для этой обратной связи является социальный статус, поскольку у Т-мужчин нейроны ГнРГ больше, несмотря на более высокий уровень андрогенов (Soma et al., 1996). Кастрированные самцы T способны сохранять свой статус, несмотря на пониженный уровень андрогенов (Francis et al. , 1992 b ). Возможно, предыдущий опыт доминирования со стороны Т-самца и разница в размерах животных способствуют этому результату. Возможно, но менее вероятно, что индивидуальное признание также могло сыграть роль.
Очевидно, что социальный статус регулирует выработку и выброс ГнРГ в гипофиз. Другой потенциальный сайт регуляции — рецептор GnRH в гипофизе.Недавняя работа в нашей лаборатории показала, что H. burtoni имеет гены, которые кодируют два разных рецептора GnRH (Robison et al. , 2001). Используя ПЦР в реальном времени, мы смогли показать, что мРНК одного из этих типов рецепторов быстро и резко повышается в гипофизе Т-самцов по сравнению с NT-самцами (Au et al. , 2003). Остается выяснить, является ли эта регуляция рецепторов результатом исключительно изменения социального статуса или же задействованы другие факторы.
Интересной особенностью H. burtoni и других видов цихлид является их яркая окраска и критическая роль зрения в социальных взаимодействиях. Недавняя демонстрация того, что видимость влияет на внутривидовую коммуникацию у цихлид озера Виктория, так что помутнение может вызвать исчезновение видов, подчеркивает этот момент (Seehausen et al. , 1997). Производство и обнаружение визуальных сигналов подверглось естественному и половому отбору, о чем свидетельствует нейронный контроль черной полосы под глазом в H.Бертон . Глазная перемычка контролируется небольшой ветвью VI черепного нерва, который контролирует миграцию гранул меланина, чтобы изменить цвет с прозрачного на черный (Muske and Fernald, 1987, , a ). Глазные перегородки Т-самцов намного более чувствительны к нейромедиатор, норэпинефрин, чем у NT-самцов. Кроме того, в долгосрочной перспективе глазная полоса Т-самцов вставляет иридифоры позади черного пигмента, увеличивая контраст и эффективность сигнала глазной полоски (Muske and Fernald, 1978, b ).Это означает, что на клеточном и молекулярном уровне как контроль, так и эффективность этого визуального сигнала также зависят от социального состояния. По сути, на всех изученных уровнях существуют социально-индуцированные изменения физиологии, лежащие в основе различий T-NT.
Когда фон Икскюлл (1921) впервые описал умвельт животного, он осознал, что среда обитания важна для животных и, вероятно, будет рассматриваться иначе, чем человек-наблюдатель. Разработка умвельта для включения в него социального мира подразумевает, что среда обитания может оказывать прямое влияние на социальную структуру ( e.г., Lott, 1982). В случае H. burtoni исследования показали, что сложность среды обитания влияет на долю самцов T, способных поддерживать территорию, а стабильность этой среды обитания влияет на продолжительность владения территорией (Hofmann et al. , 1999). Поскольку среда обитания возле озера Танганьика подвержена сильным ежедневным ветрам и, следовательно, нарушению правил, социальное регулирование воспроизводства, роста и развития кажется адаптивным. Не все самцы могут быть Т-самцами и, следовательно, размножаться в любой момент времени ( ок. 10–30%). Хотя эти животные кажутся более уязвимыми для птичьих хищников (Fernald and Hirata, 1977, b ), у нас нет количественных данных, подтверждающих это. Яркоокрашенные животные по-разному привлекательны для хищников, как было показано на примере нескольких видов рыб, следствием чего является дифференцированный отбор по этим особям (, например, Haas, 1976; Endler, 1988, 1991; Brick, 1998; O’Steen et al. , 2002; Godin and McDonough, 2003), хотя мы не знаем, верно ли это для H.Burtoni. Легко представить, что репродуктивные возможности могут быстро приходить и уходить, что, возможно, объясняет асимметричный ответ нейронов ГнРГ на изменения социального статуса (, например, Рис. 7).
Наш анализ роли среды обитания в социальных изменениях привел к ряду интересных выводов. Во-первых, это внутренняя нестабильность в обслуживании территорий (Hofmann and Fernald, 2000). Мы показали, что нестабильность вызвана дифференцированными темпами роста. Скорость роста, измеренная у взрослых особей, сильно отличается от той, которая наблюдается у молодых животных, описанных выше.В раннем возрасте (, например, от 0 до 14–21 недель) животные, подверженные социальному влиянию со стороны сородичей, могут иметь замедленный рост и задержку репродуктивного развития (см. Выше). Эта ранняя форма социального влияния несколько отличается от той, которую испытывают взрослые животные того же размера, живущие в социальных колониях. Мы объясняем это различие рядом факторов, которые отличают раннее подавление от социального взаимодействия среди старых животных. У животных почти одинакового размера, по-видимому, существуют поведенческие стратегии, которые позволяют животным успешно функционировать среди более крупных особей и избегать регуляции размера тела, но не регуляции гонад.В результате НТ и NT → Ts растут быстрее, чем Ts и T → NT (рис. 8). Наблюдения показывают, что самцы Т, хотя и могут иметь скачок роста после получения Т-статуса, начинают расходовать энергию с гораздо большей скоростью, чем самцы NT. Эта повышенная стоимость энергии приводит к замедлению роста самцов (Hofmann and Fernald, 2000). Второе открытие заключается в том, что социальная регуляция роста у взрослых может зависеть от высвобождения соматостатина в гипофизе, где этот нейрогормон подавляет высвобождение гормона роста (GH; Brazeau et al., 1973; Гиллис, 1997; Lin et al. , 2000; Очень et al. , 2001). Эту идею поддерживает тот факт, что соматостатин-содержащие нейроны в POA изменяют размер при изменении социального статуса и, следовательно, скорости роста (Hofmann and Fernald, 2000 и рис. 8).
Животные теряют территории, потому что скорость их роста снизилась, а в некоторых случаях самцы T даже уменьшились в размерах. Как отмечалось выше, похоже, что поведенческий стресс может сыграть свою роль. Как показано Fox et al. (1997) в H. burtoni переключение статуса в обоих направлениях может сопровождаться повышенными уровнями основного гормона стресса кортизола, причем изменение T → NT демонстрирует наиболее выраженное повышение. NT → T рыбы с повышенным уровнем кортизола обычно не сохраняли территориальность. T → ΝΤ мужчины постоянно имели высокий уровень кортизола. Как было показано на примере другой цихлиды, тилапии Oreochromis mossambicus , хроническое введение кортизола приводит к снижению массы тела и репродуктивных параметров, таких как размер гамет и уровни половых стероидов (Foo and Lam, 1993).Хотя регуляторные взаимодействия между гормоном роста и кортизолом очень сложны (Thakore and Dinan, 1994; van Weerd and Komen, 1998, для критических обзоров), in vivo эксперименты продемонстрировали ингибирующий эффект глюкокортикоидов на соматический рост у многих позвоночных, включая рыб. (, например, Пикеринг, 1990).
Fox et al. (1997) показали, что уровни кортизола в Ts и NT не различаются, пока сообщество рыб нестабильно, но когда стабильность достигается, у самцов T низкий уровень кортизола, а у самцов NT высокий уровень кортизола.Поскольку NT-самцы могут расти быстрее, чем T-самцы, их рост не может быть эффективно подавлен кортизолом, но другие факторы могут стать важными. Недавно мы идентифицировали, клонировали и охарактеризовали рецепторы кортизола у H. burtoni (Greenwood et al. , 2003). Интересно, что в H. burtoni существует четыре формы рецепторов кортизола, и количественная ПЦР выявила дифференциальное распределение их экспрессии. Селективное связывание кортизола с этими рецепторами показало совершенно разные уровни ответа, предполагая, что животное могло регулировать свою реакцию на кортизол, изменяя экспрессируемый подтип рецептора.Учитывая социальную модуляцию рецептора GnRH, это не может быть неожиданностью.
Остается важный и сложный вопрос: как социальная информация вызывает клеточные и молекулярные изменения в головном мозге и нервной системе. H. burtoni имеют стабильные социальные взаимодействия, требующие, чтобы они следовали правилам в своем поведении по отношению к другим. Для этого они используют информацию о других животных, основанную на социальном и репродуктивном состоянии и недавних поведенческих встречах. Ясно, что все это поведение поддерживается физиологическими, клеточными и, в конечном итоге, молекулярными механизмами.Понимание того, как происходит такой контроль, зависит от одновременной оценки многих животных в экологически реалистичном контексте. Наша недавняя работа предполагает, что H. burtoni внимательно следят за своим окружением и реагируют должным образом, чего мы не ожидали. Чтобы проверить их реакцию в надежных социальных контекстах, мы разрабатываем виртуальную рыбу, которая позволит нам предъявлять повторяющиеся стимулы в социальном контексте (Rosenthal, 1999), чтобы наблюдать за животными в социальных ситуациях, которые можно точно воспроизвести.Уменьшение дисперсии, которая является центральной частью взаимодействия многих животных, поможет нам отличить важные взаимодействия от остальных. Кроме того, мы разрабатываем нейроанатомические методы маркировки, позволяющие отслеживать активные цепи, когда животные переживают социальные изменения. Таким образом мы сможем понять, где и когда определенные области мозга играют роль в социальной реакции. Наконец, новый проект по глобальному анализу экспрессии генов у животных, которые испытали различные социальные ситуации, должен дать нам представление о том, какие наборы генов могут быть важны для успешного социального взаимодействия (Hofmann et al., 2001; Хофманн, 2003).
В целом модуляция мозга поведением имеет смысл в эволюционной структуре, где поведенческий фенотип является локусом давления отбора. Фенотипическая пластичность позволяет H. burtoni обратимо адаптировать свое поведение и физиологию к изменяющимся социальным возможностям, распределяя таким образом ресурсы между воспроизводством и ростом (Williams, 1966). Учитывая ограниченное территориальное пространство в их естественной среде обитания, селективное преимущество животных, которые могут быстро изменять поведение и физиологию, кажется очевидным.Эволюция этой стратегии жизненного цикла сформировала мозг и нервную систему H. burtoni , что дает шанс понять механизмы, поддерживающие эту гибкость. Замечательное разнообразие цихлид в Африке и Южной Америке дает шанс открыть общие принципы избирательного давления среды обитания, поведения и мозга.
Рис. 1. Фотография телосложения типичных территориальных и внетерриториальных самцов. Вверху: Самцы, не принадлежащие к территории, имеют камуфляжную окраску без четких отметин, характерных для их территориальных собратьев. Внизу: Территориальные самцы ярко окрашены, включая оранжевую плечевую чешую, с характерными пятнами на анальных плавниках, темным лбом и полосами слезы. Общий цвет тела может быть либо желтым, либо синим.
Рис. 1. Фотография строения тела типичных территориальных и внетерриториальных самцов. Вверху: Самцы, не принадлежащие к территории, имеют камуфляжную окраску без четких отметин, характерных для их территориальных собратьев. Внизу: Территориальные самцы ярко окрашены, включая оранжевую плечевую чешую, с характерными пятнами на анальных плавниках, темным лбом и полосами слезы.Общий цвет корпуса может быть желтым или синим
Рис. 2. Развитие и созревание мальков H. burtoni , выращенных группами (белые и темные кружки) или физически изолированными (ромбы). ( A ) Вес тела показан как функция времени. Звездочки означают, что территориальные рыбы, выращенные в группах (Ts, закрашенные кружки), весят значительно больше через 10 и 14 недель, чем их товарищи по танку NT. Обратите внимание, что по прошествии 20 недель различия в размерах больше не очевидны.( B ) Показано относительное количество зрелых сперматозоидов в поперечных срезах центральной дольки яичка. (По Fraley and Fernald, 1982)
Рис. 2. Развитие и созревание мальков H. burtoni , выращенных группами (белые и темные кружки) или физически изолированными (ромбы). ( A ) Вес тела показан как функция времени. Звездочки означают, что территориальные рыбы, выращенные в группах (Ts, закрашенные кружки), весят значительно больше через 10 и 14 недель, чем их товарищи по танку NT.Обратите внимание, что по прошествии 20 недель различия в размерах больше не очевидны. ( B ) Показано относительное количество зрелых сперматозоидов в поперечных срезах центральной дольки яичка. (По Фрейли и Фернальду, 1982 г.)
Рис. 3. Онтогенез цветовых паттернов и агонистических поведенческих паттернов у мальков H. burtoni , выращенных группами или изолированно (см. Рис. 2 выше). Символы, представляющие социальные условия, показывают средние значения ± стандартное отклонение в днях, когда каждый образец был впервые обнаружен.Происхождение — это когда мальков выпустили изо рта. (Модифицировано из Fraley and Fernald, 1982)
Рис. 3. Онтогенез цветовых паттернов и агонистических поведенческих паттернов у мальков H. burtoni , выращиваемых группами или изолированно (см. Рис. 2 выше). Символы, представляющие социальные условия, показывают средние значения ± стандартное отклонение в днях, когда каждый образец был впервые обнаружен. Происхождение — это когда мальков выпустили изо рта. (Изменено из Fraley and Fernald, 1982)
Рис.4. Вес гонад нанесен на график как функция среднего размера GnRH-иммунореактивной сомы (± стандартное отклонение) для самых больших 30% клеток. Данные для животных, выращенных без взрослых особей (Ts; закрашенные кружки) или с присутствующими взрослыми особями (NT; пустые кружки) через 20 недель. Обратите внимание, что размеры нейронов GnRH не зависят от размера тела в этом эксперименте и больших различий в размере клеток и весе семенников между T- и NT-самцами. (По Дэвису и Фернальду, 1990)
Рис. 4. Вес гонад, построенный как функция среднего размера GnRH-иммунореактивной сомы (± стандартное отклонение) для самых больших 30% клеток.Данные для животных, выращенных без взрослых особей (Ts; закрашенные кружки) или с присутствующими взрослыми особями (NT; пустые кружки) через 20 недель. Обратите внимание, что размеры нейронов GnRH не зависят от размера тела в этом эксперименте и больших различий в размере клеток и весе семенников между T- и NT-самцами. (По Дэвису и Фернальду, 1990)
Рис. 5. Трехмерное представление средних размеров сомы нейронов, содержащих GnRH, в POA в зависимости от экспериментальной группы. Обратите внимание на существенные различия между самцами T и T → NT и между самцами NT и NT → T.Вертикальная ось показывает процент людей со средним размером сомы в данной ячейке. (Изменено из Francis et al. , 1993)
Рис. 5. Трехмерное представление средних размеров сомы нейронов, содержащих GnRH, в POA в зависимости от экспериментальной группы. Обратите внимание на существенные различия между самцами T и T → NT и между самцами NT и NT → T. Вертикальная ось показывает процент людей со средним размером сомы в данной ячейке. (Изменено из Francis et al., 1993)
Рис. 6. Средний двумерный размер сомы нейронов GnRH-ir в зависимости от частоты агрессивного и подчиненного поведения, выраженный как индекс доминирования (сумма агрессивных действий минус сумма событий побега / 3-минутный интервал наблюдения). Обратите внимание на гистерезисную функцию, поскольку изменения социального статуса асимметричны по отношению к поведению и размеру сомы: хотя изменение поведения у мужчин T → NT происходит значительно быстрее ( около 1 день), чем у мужчин NT → T ( ок. 2 недели), последние достигают размеров сомы, эквивалентных размерам Ts за одну неделю, тогда как T → NT требуется три недели для их нейронов, чтобы сузиться до размеров NT. Пустые кружки указывают на случаи, когда размер сомы предсказан. Различия не показаны для ясности. (Данные White et al. , 2002)
Рис. 6. Средний двумерный размер сомы нейронов ГнРГ-ir в зависимости от частоты агрессивного и покорного поведения, выраженный в виде индекса доминирования (сумма агрессивных действий минус сумма побеги / интервал наблюдения 3 мин).Обратите внимание на гистерезисоподобную функцию, поскольку изменения социального статуса асимметричны в отношении поведения и размера сомы: хотя изменение поведения у мужчин T → NT происходит значительно быстрее ( примерно 1 день), чем у мужчин NT → T ( примерно ). 2 недели), последние достигают размеров сомы, эквивалентных таковым у Ts за одну неделю, тогда как T → NT требуется три недели, чтобы их нейроны сократились до размеров NT. Пустые кружки указывают на случаи, когда размер сомы предсказан. Различия не показаны для ясности. (Данные White et al., 2002)
Рис. 7. Модель обратной связи регуляции ГнРГ у самца H. burtoni . Нейроны в преоптической области (POA) интегрируют как социальные, так и гормональные сигналы, чтобы регулировать высвобождение гонадолиберина. В этой модели заданное значение уровня ГнРГ определяется социальными сигналами, а поддержание уровня ГнРГ в этом заданном значении достигается за счет отрицательной обратной связи от гонадных андрогенов. (Изменено из Soma et al. , 1996)
Рис. 7. Модель контроля с обратной связью регуляции GnRH у мужчин H.Бертон . Нейроны в преоптической области (POA) интегрируют как социальные, так и гормональные сигналы, чтобы регулировать высвобождение гонадолиберина. В этой модели заданное значение уровня ГнРГ определяется социальными сигналами, а поддержание уровня ГнРГ в этом заданном значении достигается за счет отрицательной обратной связи от гонадных андрогенов. (Изменено из Soma et al. , 1996)
Рис. 8. График скорости роста как функция среднего размера иммунореактивного соматостатина сомы у H. burtoni .NT и NT → T самцы (закрашенный кружок; среднее ± стандартное отклонение) имеют меньшую площадь поперечного сечения сомы и растут быстрее, чем Ts и T → NT (закрашенный ромб; среднее ± стандартное отклонение). Анализ линейной регрессии дает y = 0,19 * × + 8,95, при этом r 2 = 0,4163 ( P <0,001; n = 18). (Изменено из Hofmann and Fernald, 2000)
Рис. 8. Графики скорости роста как функция среднего размера иммунореактивной соматостатина сомы у H. burtoni .NT и NT → T самцы (закрашенный кружок; среднее ± стандартное отклонение) имеют меньшую площадь поперечного сечения сомы и растут быстрее, чем Ts и T → NT (закрашенный ромб; среднее ± стандартное отклонение). Анализ линейной регрессии дает y = 0,19 * × + 8,95, при этом r 2 = 0,4163 ( P <0,001; n = 18). (Изменено из Hofmann and Fernald, 2000)
Мне посчастливилось работать со многими талантливыми коллегами на протяжении многих лет, которых я благодарю за бесценное обсуждение и помощь в работе, описанной здесь.Поддерживается премией Джейкоба Джавитса (NIH-NS 34950) и NIH-EY-05051 для RDF.
Список литературы
Allen
,E. E.
и R. D. Fernald.1985
. Спектральная чувствительность африканской цихлиды, Haplochromis burtoni .J. Comp. Physiol. А
,157
247
-253.Au
,T. M.
, N. Illing и R. D. Fernald.2003
. Социальная регуляция рецептора гонадотропин-рилизинг-гормона у рыб цихлид, Haplochromis burtoni .Soc. Neurosci. 611,6. (Аннотация).Baerends
,G. P.
и J. M. Baerends van Roon.1950
. Введение в изучение этологии рыб-цихлид.Behavior, Supplementum
,1
1
-235.Басс
,А. Х.
, Д. А. Боднар и М. А. Маршатер.2000
. Акустическая схема среднего мозга у кричащей рыбы.J. Comp. Neurol
,419
505
-531.Берглунд
,А.
1991
. Конкуренция яиц у иглобрюхих с перевернутыми половыми ролями: субдоминирующие самки обменивают воспроизводство на рост.Evolution
,45
770
-774.Боровски
,р.
1973
. Социальный контроль размера взрослой особи у самцов Xiphophorus varatus .Природа
,245
332
-335.Brazeau
,P.
, W.Вейл, Р. Бургус, Н. Линг, М. Бутчер, Дж. Ривье и Р. Гийемен.1973
. Гипоталамический полипептид, подавляющий секрецию иммунореактивного гормона роста гипофиза.Наука
,179
77
-79.Кирпич
,О.
1998
. Боевое поведение, бдительность и риск нападения хищников у цихлид Nannacara anomala .Anim. Бех
,56
309
-317.Бшары
,р.
, W. Wickler и H. Fricke.2002
. Познание рыбы: взгляд приматов.Anim. Cogn
,5
1
-13.Бушник
,Т. Л.
и Р. Д. Фернальд.1995
. Популяция GnRH-содержащих нейронов, показывающих социально-опосредованные изменения размера, проецируется на гипофиз в костистости, Haplochromis burtoni .Brain Behav. Evol
,46
371
-377.Coulter
,г.W.
1991
. Озеро Танганьика и его жизнь . Издательство Оксфордского университета, Оксфорд.Дэвис
,М. Р.
и Р. Д. Фернальд.1990
. Социальный контроль размера нейрональной сомы.J. Neurobiol
,21
1180
-1188.Эндлер
,Дж. А.
1988
. Частотно-зависимое хищничество, крипсис и апосематическая окраска.Фил. Пер. R. Soc. London B Biol.Sci
,319
505
-523.Эндлер
,Дж. А.
1991
. Различия во внешнем виде цветовых узоров гуппи для гуппи и их хищников в различных визуальных условиях.Vision Res
,31
587
-608.Enquist
,M.
и С. Якобссон.1986
. Принятие решений и оценка боевого поведения Nannacara anomala (Cichlidae, Pisces).Этология
,72
143
-153.Enquist
,M.
и О. Леймар.1983
. Эволюция боевого поведения: правила принятия решений и оценка относительной силы.J. Теорет. Биол
,102
387
-410.Фернальд
,Р. Д.
1977
. Количественные наблюдения Haplochromis burtoni в полуестественных условиях.Anim. Поведение
,25
643
-653.Фернальд
,Р. Д.
1980
. Ответ самцов цихлид, Haplochromis burtoni , выращенных изолированно на моделях сородичей.Zeitschrift für Tierpsychologie
,54
85
-93.Фернальд
,Р. Д.
1981
. Хромная организация сетчатки цихлид.Vision Res
,21
1749
-1753.Фернальд
,р.Д.
1983
. Нейронные основы распознавания зрительных образов у рыб. В Дж. П. Эверт, Р. Р. Капраника и Д. Дж. Ингл (ред.), Успехи в нейроэтологии позвоночных, стр. 569–580. Пленум Пресс, Нью-Йорк.Фернальд
,Р. Д.
1984
. Зрение и поведение африканских цихлид и рыб.г. Sci
,72
58
-65.Фернальд
,Р. Д.
1989
.Нейрогенез палочек сетчатки. В Б. Л. Финли и Д. Р. Сенгелауб (ред.), Развитие сетчатки позвоночных, стр. 31–42. Пленум Пресс, Нью-Йорк.Фернальд
,Р. Д.
2000 9000 5. Сенсорные системы 15.1 Зрение — общая функциональная анатомия. In G. K. Ostrander (ed.), Laboratory fish , pp. 225–235. Academic Press, London.
Фернальд
,Р. Д.
2000 9000 5. Сенсорные системы 27.1 Визуально-микроскопическая функциональная анатомия. В Г. К. Острандер (ред.), Лабораторные рыбы , стр. 451–462. Академик Пресс, Лондон.
Фернальд
,Р. Д.
и Н. Р. Хирата.1977
. Полевое исследование Haplochromis burtoni : количественные наблюдения за поведением.Anim. Behav
,25
964
-975.Фернальд
,Р. Д.
и Н. Р. Хирата.1977
. Полевое исследование Haplochromis burtoni : Среда обитания и сожители.Environ. Биол. Рыбы
,2
299
-308.Fernald
,R. D.
и P. A. Liebman.1980
. Пигменты зрительных рецепторов у африканских рыб-цихлид, Haplochromis burtoni .Vision Res
,20
857
-864.Фернальд
,Р. Д.
и С. Э. Райт.1983
. Поддержание оптического качества при росте хрусталика.Природа
,301
618
-620.Fernald
,Р. Д.
и Р. Б. Уайт.1999
. Гены гонадотропин-рилизинг-гормона: филогения, структура и функции.Frontiers Neuroendocrinol
,20
224
-240.Foo
,J. T. W.
и T. J. Lam.1993
. Замедление роста яичников и снижение уровня сывороточных стероидов у тилапии Oreochromis mossambicus путем имплантации кортизола.Аквакультура
,115
133
-143.Fox
,H. E.
, S. A. White, M. H. F. Kao и R. D. Fernald.1997
. Стресс и доминирование у социальной рыбы.J. Neuro
,17
6463
-6469.Fraley
,N. B.
и R. D. Fernald.1982
. Социальный контроль скорости развития африканской цихлиды, Haplochromis burtoni .Zeitschrift für Tierpsychologie
,60
66
-82.Фрэнсис
,Р. К.
1988
. Социально опосредованные вариации в скорости роста цихлиды midas: главенство ранних различий в размерах.Anim. Поведение
,36
1844
-1845.Фрэнсис
,Р. К.
, Б. Якобсен, Дж. К. Вингфилд и Р. Д. Фернальд.1992
. Гипертрофия нейронов, содержащих гонадотропин-рилизинг-гормон, после кастрации костистых зубов, Haplochromis burtoni .J. Neurobiol
,23
1084
-1093.Фрэнсис
,Р. К.
, Б. Якобсен, Дж. К. Вингфилд и Р. Д. Фернальд.1992
. Кастрация снижает агрессивность, но не снижает социальное доминирование у самцов Haplochromis burtoni (Cichlidae).Этология
,90
247
-255.Фрэнсис
,Р. К.
, К. Сома и Р. Д. Фернальд.1993
. Социальная регуляция оси мозг-гипофиз-гонад.Proc. Nat. Акад. Sci. США
,90
7794
-7798.Гиллис
,г.
1997
. Соматостатин: нейроэндокринная история.Trends Pharmacol. Sci
,18
87
-95.Godin
,J. G. J.
и H. E. McDonough.2003
. Предпочтение хищниками ярких самцов у гуппи: цена жизнеспособности по выбранному половому признаку.Behav. Экол
,14
194
-200.Годвин
,J.
, J. A. Luckenbach и R. J. Borski.2003
. Экология встречается с эндокринологией: определение пола у рыб в окружающей среде.Evol. Дев
,5
40
-49.Гринвуд
,А. К.
, П. Батлер, У. Де Марко, Р. К. Уайт, Д. Пирс и Р. Д. Фернальд.2003
. Множественные рецепторы кортикостероидов у костистых рыб: отчетливые последовательности, паттерны экспрессии и транскрипционная активность.Эндокринология
,144
4226
-4236.Haas
,Р.
1976
. Половой отбор у Nothobranchius-guentheri рыбы Cyprinodontidae.Evolution
,30
614
-622.Heiligenberg
,W.
и U. Kramer.1972
. Агрессивность как функция внешнего раздражения.J. Comp. Physiol. Психол
,77
332
-340.Heiligenberg
,W.
, U. Kramer и V. Schulz.1972
. Угловая ориентация черной полосы под глазом у Haplochromis burtoni (Cichlidae, Pisces) и ее отношение к агрессивности.Zeitschrift für vergleichende Physiologie
,76
168
-176.Hofmann
,H.A.
2003
. Функциональная геномика нейронной и поведенческой пластичности.J. Neurobiol
,54
272
-282.Hofmann
,H.A.
, T. Barrett, A.K. Greenwood, K.G.Becker и R.D. Fernald.2001
. Социальный статус контролирует профили экспрессии генов у африканских рыб-цихлид. Soc. Neurosci. 611,6. (Тезисы).Hofmann
,H.A.
, M.E.Benson и R.D. Fernald.1999
. Социальный статус регулирует скорость роста: последствия для стратегий жизненного цикла.Proc. Nat. Акад. Sci. США
,96
14171
-14176.Hofmann
,H.A.
и R. D. Fernald.2000
. Социальный статус контролирует размер и рост соматостатин-нейронов.Журнал неврологии
,20
4740
-4.Johns
,P. R.
и R. D. Fernald.1981
. Генезис стержней сетчатки костистых рыб.Природа
,293
141
-142.Лин
,X. W.
, К. Дж. Отто, Р. Карденас и Р.Э. Петр.2000
. Семейство пептидов соматостатина и его рецепторы у рыб.банка. J. Physiol. Pharmacol
,78
1053
-1066.Леонг
,C.-Y.
1969
. Количественное влияние пусковых агентов на готовность рыбы к атаке Haplochromis burtoni .Zeitschrift für vergleichende Physiologie
,65
29
-50.Лоренц
,К.
1932
.Der Kumpan in der Umwelt des Vogels. J. f. Ornithologie 80, Heft 1.Lorenz
,K.
1981
. Основы этологии (Vergleichende Verhaltensforschung) . Спрингер-Верлаг, Нью-Йорк.Лотт
,Д. Ф.
1982
. Внутривидовая изменчивость социальных систем диких позвоночных. Кембриджские исследования по поведенческой биологии . Издательство Кембриджского университета, Кембридж.Муске
,л.E.
и Р. Д. Фернальд.1987
. Контроль социального сигнала костистости. I. Нейронная основа дифференциального выражения цветового узора.J. Comp. Physiol. А
,160
89
-97.Muske
,L. E.
и R. D. Fernald.1987
. Контроль социального сигнала костистости. II. Анатомо-физиологические особенности хроматофоров.J. Comp. Physiol. А
,160
99
-107.Oliveira
,R. F.
, V. C. Almada и V. Caniaro.1996
. Социальная модуляция концентраций половых стероидов в моче самцов цихлид Orechromis mossambicus .Horm. Поведение
,30
2
-12.O’Steen
,S.
, A. J. Cullum и A. F. Bennett.2002
. Быстрая эволюция способности к бегству у тринидадских гуппи ( Poecilia reticulata ).Внутр.J. Org. Evol
,56
776
-84.Пикеринг
,А. Д.
1990
. Стресс и подавление соматического роста костистых рыб. In A. Epple, C.G. Scanes и M. H. Stetson (ред.), Progress in Clinical and Biological Research, Vol. 342, Успехи сравнительной эндокринологии, с. 473–479. Вили-Лисс, Нью-Йорк.Розенталь
,Г. Г.
1999
.Использование воспроизведения видео для изучения сексуального общения.Environ. Биол. Рыбы
,56
307
-316.Робисон
,Р. Р.
, Р. Д. Фернальд и Н. Э. Стейси.1998
. Обонятельный ответ на предполагаемые химические сигналы рыб-цихлид ( Haplochromis burtoni ).J. Fish Biol
,53
226
-229.Робисон
,Р. Р.
, Р. Б. Уайт, Н. Иллинг, Б. Э. Троски, М.Морли, Р. П. Миллар и Р. Д. Фернальд.2001
. Рецептор гонадолиберина в костистых костях Haplochromis burtoni : структура, расположение и функция.Эндокринология
,142
1737
-1743.Schultz
,E. T.
, L. M. Clifton и R. R. Warner.1991
. Энергетические ограничения и тактика, основанная на размерах: адаптивное значение изменения графика размножения у морской рыбы, Micrometrus minimus (Embiotocidae).г. Natur
,138
1408
-1430.Seehausen
,O.
, J. J. M. Van Alphen и F. Witte.1997
. Разнообразию рыб цихлид угрожает эвтрофикация, ограничивающая половой отбор.Science
,277
1808
-1811.Soma
,K. K.
, R. C. Francis, J. C. Wingfield и R. D. Fernald.1996
. Андрогенная регуляция гипоталамических нейронов, содержащих гонадотропин-рилизинг гормон, у цихлид: интеграция с социальными сигналами.Horm. Поведение
,30
216
-226.Thakore
,J. H.
и T. G. Dinan.1994
. Секреция гормона роста: роль глюкокортикоидов.Life Sci
,55
1083
-1099.Тинберген
,N.
1948
. Социальные релизеры и экспериментальный метод, необходимый для их изучения.Уилсон Булл
,60
6
-52.Тинберген
,Н.
1951
. Изучение инстинктов . Кларендон Пресс, Оксфорд.van Weerd
,J. H.
и J. Komen.1998
. Влияние хронического стресса на рост рыб: критическая оценка.Сост. Биохим. Physiol
,120
107
-112.Verheyen
,E.
, W. Salzburger, J. Snoeks и A. Meyer.2003
. Происхождение суперстайки цихлид из озера Виктория, Восточная Африка.Science
,300
325
-329.Very
,N. M.
, D. Knutson, J. D. Kittilson и M. A. Sheridan.2001
. Соматостатин подавляет рост радужной форели.Дж. Фиш Биол
,59
157
-165.Фон Икскюлл
,J.
1921
. Umwelt und Innenweld der Tiere. Берлин.Белый
,S.A.
и R. D. Fernald.1993
.Нейроны, содержащие гонадотропин-рилизинг-гормон, изменяют размер в зависимости от репродуктивного состояния у самок Haplochromis burtoni .J. Neurosci
,13
434
-441.White
,S.A.
, К. Т. Бонд, Р. К. Фрэнсис, Т. Л. Кастен, Р. Д. Фернальд и Дж. П. Адельман.1994
. Второй ген GnRH: кДНК и характер экспрессии.Proc. Nat. Акад. Sci. США
,91
1423
-1427.Белый
,S.A.
, T. L. Kasten, C. T. Bond, J. P. Adelman и R. D. Fernald.1995
. Три гена гонадотропин-рилизинг-гормона в одном организме предполагают новую роль древнего пептида.Proc. Nat. Акад. Sci. США
,92
8363
-8367.Белый
,S. A.
, T. Nguyen и R. D. Fernald.2002
. Социальная регуляция экспрессии генов при изменении социального статуса мужчин и женщин.J. Exp. Биол
,205
2567
-2581.Виклер
,W.
1962
. Zur Stammesgeschichte funktionell korrelierter Organ- und Verhaltensmerkmale: Eiattrappen und Maulbrüten bei afrikanischen Cichliden.Zeitschrift für Tierpsychologie
,19
129
-164.Винтер
,Ю.
, Х. Лопес и О. фон Хелверсон.2003
. Ультрафиолетовое зрение у летучей мыши.Природа
,425
612
-624.Уильямс
,г.С.
1966
. Адаптация и естественный отбор . Издательство Принстонского университета, Принстон, Нью-Джерси.Zygar
,C.A.
, M. L. Lee и R. D. Fernald.1999
. Назотемпоральная асимметрия роста сетчатки костистых желез: поддержание области специализации.J.