Физиологические основы памяти
1.Процессы запоминания, сохранения воспроизведения.
2.Некоторые закономерности процесса запоминания.
3.Рациональные способы заучивания учебного материала.
4.Виды и типы памяти.
Литература
1.Асмолов А.Г.
«Принципы организации памяти человека» Москва 1983
2.Вейн А.М. Каменецкая Б.И.
«Память человека» Москва 1973
3.Зинченко П.И.
«Непроизвольные типы запоминания»
4.Ляудис
«Память в процессе развития»
5.Роговин
«Проблемы психологии в развитии»
Общие понятия о памяти.
Процесс запечатления,сохранения,воспроизведение со знанием прошлого опыта называется памятью. Реминисценция — отсроченное на два три дня воспроизведение заученного материала. Память как и восприятие и ощущение является процессом отражения,но отражается не только действует в данный момент,но и то что было в прошлом. Амнезия — забывание. Гипоамнезия — общее забывание.
Физиологические основы памяти.

Физиологческой основой памяти является образование и закрепление нервных связей в коре головного мозга с их последующим оживлением при воздействии различных раздражителей. Существуют 4 теории памяти.
-ассоциативная теория. Эта теория утверждает что в коре головного мозга образуются связи или ассоциации. Чем больше связей тем лучше логическая память и мышление. Выделяют простые и сложноые ассоциации. Аристотель указывал на ассоциации по смежности сходству по контрасту.
Современная научная Классификация ассоциативная
Классификация по характеру связей
а) вид- род(стол — мебель)
б) часть целое(корма — корабль)
в) по контрасту (белый- черный)
г) по определению св-ва предмета
д)по адекватности (пещера — убежище)
е) по отрицанию ( щедрый — не щедрый)
—
нейро-физиологическая теория. Мозговые
нейроны образуют замкнутые цепи по
которым совершаются движение нервных
импульсов там сохраняется информация
поступившая и переработанная в этих
цепях.
химическая теория. В определенных объединениях нервных клеток есть так называемые тельца Ниссля, которые содержат ДНК,которая является носителем генетической памяти.
Теория условных рефлексов. В нервной системе человека хранится два вида информации. Информация накопленная в ходе эволюции. Она проявляется в виде безусловных рефлексов,инстинктов. Информация приобретает влияние в индивидуальной жизни.
3)Процессы
запоминания,сохранения воспроизведения.
Память включает в себя следующие
психические процессы — запоминание,
сохранение усвоенного, узнавание
воспроизведение. То есть установление
связей между предметами и явлениями.
Процесс запоминания характеризуется
различной степенью сознательной волевой
активности. В зависимости от этого
выявляют два виды запоминания.
Непроизвольное и произвольное.
Непроизвольное запоминание происходит
без постановки специальной цели и без
использование определенных приемов
запоминания.
Произвольное запоминание характеризуется целенаправленностью и предполагает использование специальных приемов. Подготовка к будничной профессии требует специального запоминания. В зависимости от степени мыслительной активности запоминание бывает механическим и логическим. Механическое запоминание состоит ттолько в повторении. Иногда повторение происходит без понимание исходного материала. Такое запоминание дается с трудом.
При осмысленном запоминании большое значение имеют процессы мышления. Человек старается понять что надо запоснить. Установить связь нового со старым. Важным условием ясного и твердого запоминания явяются способы заучивания.
Рациональные способы заучивания учебного материала. Для успешного запоминания учебного материала пользется приемами осмысленного запоминания.
группировка материала
деление его на части
установление связи между частями
выделение смысловых опорных пунктов
озаглавливание частей текста
составление плана будущего ответа
повторение заученного материала
Важен
не сам план а осмысленная работа
проводившаяся при его составлении.
Узнавание и воспроизведение. Простой формай воспроизведение является узнавание которое имеет место при повторном восприятии уже знакомого обьекта. Различают два вида воспроизведения. Нерпеднамеренное и преднамеренное. Непреднамеренное воспроизведение осуществляется без заранее поставленной цели, когда образ прошлого независимо от нашего желания возникает в сознании. Но при выполнении любой работы чаще приходится опиратся на преднамеренное воспроизведение которое связанно с мышлением. Активный мыслительный процесс требующий волевых усилий при воспоизведении называется припоминанием. Яркое изображение этого процесса дает антон палыч чехов в рассказе лошадиная фамилия.
Иногда проявляется еще одна форма непреднамеренного воспроизведения,которая называется персеверация(навязчивость). В какой-то мере она есть у всех людей. Систематическое возникновение персеверации является признаком заболевания.
Сохранение
и забывание. Необходимым условием
воспроизведения является сохранение
материала. Сохранение с физиологической
точки зрения обьясняется пластичностью
невной системы, которая проявляется в
способности хранить ранее образовавшиеся
неврые связи.Павлов подчеркивает что
новые связи хорошо сохранаяются если
они хорошо закреплены.
Сохранение с физиологической
точки зрения обьясняется пластичностью
невной системы, которая проявляется в
способности хранить ранее образовавшиеся
неврые связи.Павлов подчеркивает что
новые связи хорошо сохранаяются если
они хорошо закреплены.
Виды и типы памяти. В зависимости от удержанногов памяти материала различают наглядно-образную словесно-логическую и эмоциональную память.
Мышление.
1)Общие понятия о мышлении.
2)Физиологические основы мышления.
3)Операции мышления.
4)Представление,понятие и понимание.
Литература:
1.Энгельс
«Роль труда в процессе превращения обезьяны в человека»
2.К.Маркс и Ф.Энгельс Том 20.
3.Рубинштейн С.Л.
«О мышлении и путях его исследования» Москва 1958
4.Матюшкин А.М.
«Психология мышления» Москва 1965
5.Соколов А.Н.
« Внутренняя речь и мышление» Москва 1968
6.Выходский Л.С.
«Мышление и речь» Москва 1956
Полное
собрание сочинений Том 2.
7.Тихомиров О.К.
«Структура мыслительной деятельности человека» Москва МГУ 1969
1) Общие понятия о мышлении.
Познание начинается с ощущений и восприятий , которые сами опираются на память, то есть на те следы которые образовались в процессе предыдущей деятельности.
Но познание опирающееся на на ощущение и восприятие поверхностное, так как при ощущении и восприятии человек отражает только внешний признаки и свойства предметов. В мышлении же человек отражает такие свойства предмета , явления, которые не представляются в визуальном отражении. Мышление это психический процесс отражения закономерных связей и отношений между предметами.
2)Физиологические основы мышления.
Мышление
будучи сложным процессом опирается на
ощущение, восприятие, память, эмоции и
в свою физиологическую основу включает
физиологические механизмы данных
процессов. Сущность физиологических
процессов заключается в образовании
новых связей в коре головного мозга, в
результате сложной аналитико-синтетической
деятельности коры в целом.
3) Операции мышления.
Всякое мышление осуществляется при помощи отдельных операций,которые выступают парами.
а) Сравнение и противопоставление.
В результате сравнение и противопоставления,человек находит различия, так же осуществляется связь старого с новым.
б) Анализ и синтез. Анализ — мысленное разложение целого на части на единой основе. Синтез — соединение разрозненных элементов в целое на единой основе. Анализ и синтез осуществляются одновременно.
в)Абстрагирование и конкретизация. Абстрагирование — отрыв, отличие свойства предмета от самого предмета.
г) Обобщение и систематизация.
4) Представление понятие понимание , в процессе познание человек всегда оперирует имеющисися знаниями.
Физиологические основы памяти
Память
начинается с запоминания той информации,
которую принимают наши органы чувств
из окружающего мира. Все образы, слова,
вообще впечатления должны удержаться,
остаться в нашей памяти. В зависимости
от критериев, перечисленных далее, можно
говорить о разных видах памяти.
Все образы, слова,
вообще впечатления должны удержаться,
остаться в нашей памяти. В зависимости
от критериев, перечисленных далее, можно
говорить о разных видах памяти.
В нейрологической памяти выделяют:
— генотипическую (врожденную) память, которая обуславливает сохранение инстинктов, импринтинга;
— фенотипическую память, мозговые механизмы которой обеспечивают обработку и хранение информации, приобретаемой организмом в процессе индивидуального развития (Смирнов В.М., Свешников Д.С., Яковлев В.Н., 2006).
1. По длительности хранения информации выделяют:
Мгновенная (сенсорная, иконическая) (0,1 – 0,5 с)
– удержание точной и полной картины только что воспринятого органами чувств информации.
Кратковременная (до 20 с)
– представляет
собой способ хранения информации в
течение короткого промежутка времени. В ней сохраняется наиболее существенные
элементы образа. Из мгновенной памяти
в нее попадает только та информация,
которая привлекает к себе повышенное
внимание.
В ней сохраняется наиболее существенные
элементы образа. Из мгновенной памяти
в нее попадает только та информация,
которая привлекает к себе повышенное
внимание.
Оперативная (промежуточная) (до нескольких дней)
– хранение информации в течение определенного, заранее заданного срока. Срок хранения информации в этой памяти определяется задачей вставшей перед человеком.
Долговременная
— неограниченное хранение информации в неограниченном промежутке времени. Данная информация может воспроизводиться сколько угодно раз без утраты.
2. По характеру запоминаемого материала:
Моторная (двигательная)
— запоминание и сохранение, а при необходимости и воспроизведение с достаточной точностью многообразных сложных движений;
Эмоциональная
—
память на переживания. То, что у человека
вызывает эмоциональные переживания,
запоминается им без особого труда и на
более длительный срок.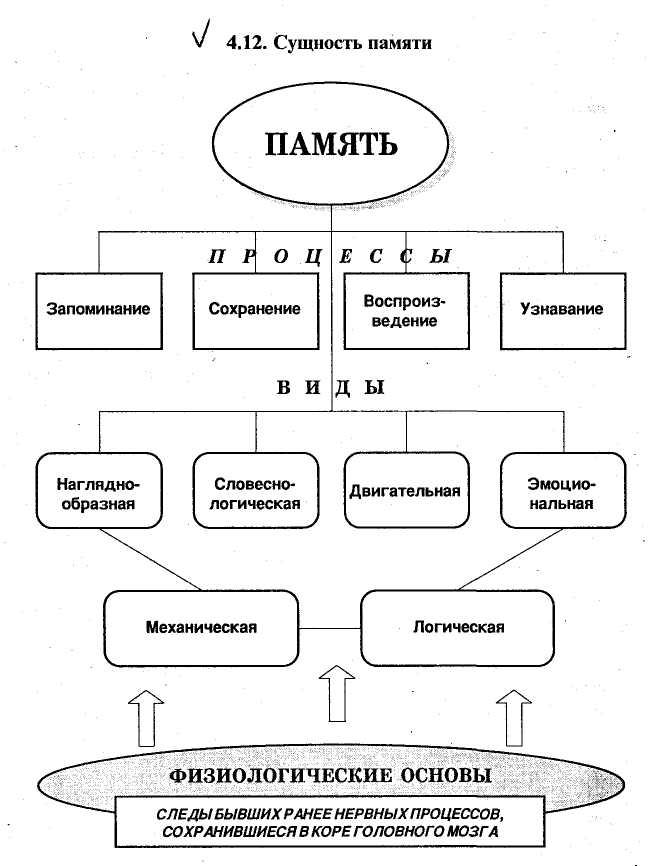
Образная
— обеспечивает запоминание наглядного материала (предметов, изображений). Это память на представления, лица, звуки, запахи, формы предметов и т.д.
Вербальная (словесно-логическая)
— позволяет запоминать информацию с помощью слов, опосредованно, с помощью смысловой структуры. Содержание этого вида памяти являются наши мысли. Словесно-логическая память существует только у человека, в отличие от двигательной, эмоциональной, образной, которая есть и у животных (Рубинштейн С. Л., 1999).
3. По модальности запоминания в зависимости от того, какой анализатор принимает наибольшее участие в процессе запоминания, выделяют следующие виды памяти:
Зрительная
— сохранение и воспроизведение зрительных образов;
Слуховая
— запоминание и точное воспроизведение разнообразных звуков;
Осязательная, обонятельная, вкусовая …
—
удовлетворение биологических потребностей
или потребностей связанных с безопасностью
и самосохранением организма.
Двигательная;
Пространственная;
Чаще всего все эти виды памяти развиты неодинаково.
4. По характеру мнестической деятельности в зависимости от наличия или отсутствия цели запомнить информацию выделяют:
Непроизвольное запоминание
— происходит автоматически и без особых усилий со стороны человека, без постановки перед ним специальной мнемической задачи.
Произвольное запоминание
— процесс запоминания и воспроизведения требует волевых усилий. При условии наличия мнемической задачи.
5. По способу запоминания:
Непосредственное (механическое запоминание)
—
процесс сохранения информации происходит
без участия мышления, т.е. без осмысления
материала, без установления логической
структуры и использования ассоциативных
приемов запоминания. В обычной жизни
это называется «зазубрил».
В обычной жизни
это называется «зазубрил».
Опосредованная память
— процесс запоминания опосредован логикой, мышлением и, запоминаемый материал здесь необходимо предварительно осмыслить (Сунцова А.В., Курдюкова С.В., 2009).
Во всех вышеперечисленных видах памяти имеет место фиксация информации, включающая в себя, по крайней мере, три этапа:
1.формирование энграммы, т.е. следа, оставляемого в мозгу тем или иным событием;
2.сортировка и выделение новой информации;
3.долговременное хранение значимой информации (Смирнов В.М., Будылина С.М, 2004).
Накопление
и хранение информации в памяти
обеспечивается за счет электрических
и химических процессов, происходящих
в мозге и обуславливающих происходящие
в нем структурные изменения. Электрическая
активность нервных цепей, несомненно,
лежит в основе получения «непосредственного
отпечатка» сенсорной информации и
кратковременной памяти. Нервная цепь,
обеспечивающая циркуляцию возбуждения,
вызванного стимулом, должна сохранять
специфическую активность, связанную с
действием раздражителя и после его
выключения (Ткаченко Б.И., 1994).
Нервная цепь,
обеспечивающая циркуляцию возбуждения,
вызванного стимулом, должна сохранять
специфическую активность, связанную с
действием раздражителя и после его
выключения (Ткаченко Б.И., 1994).
Физиологической основой памяти являются следы в нервной системе от предыдущих раздражений. Следовые процессы являются общим свойством нервной системы, и поэтому трудно предполагать наличие каких-либо специализированных центров памяти (Шмидт Р., Тевс Г., 1996).
Пионер
в области исследования памяти Карл
Лешли пытался с помощью хирургического
вмешательства в мозг дать ответ о
пространственном расположении памяти,
по аналогии с речевыми, моторными или
сенсорными зонами. Лешли обучал разных
животных решать определенную задачу.
Потом он удалял у этого животного один
за другим различные участки коры — в
поисках места расположения следов
памяти — энграмм. Однако, независимо
от того, какое количество корковой ткани
было удалено, найти то специфическое
место, где хранятся следы памяти
(энграммы), Лешли не удалось. Свою
классическую статью он закончил выводом
о том, что память одновременно находится
в мозгу везде и нигде.
Свою
классическую статью он закончил выводом
о том, что память одновременно находится
в мозгу везде и нигде.
В последствии, результаты морфологических и физиологических исследований способствовали формированию представлений о существовании кольцевой системы: гиппокамп – гипоталамус — ретикулярная формация. Эта филогенетически древняя система объединена прямыми и обратными связями, через которые новая кора осуществляет саморегуляцию и регуляцию других мозговых систем (Смирнов В.М., Свешников Д.С., Яковлев В.Н., 2006).
По современным представлениям, фиксация следа в памяти осуществляется в три этапа.
Вначале, в иконической памяти на основе деятельности анализаторов возникают сенсорный след (зрительный, слуховой, тактильный и т. п.). Эти следы составляют содержание сенсорной памяти.
На
втором этапе сенсорная информация
направляется в высшие отделы головного
мозга. В корковых зонах, а также в
гиппокампе и лимбической системе
происходит анализ, сортировка и
переработка сигналов, с целью выделения
из них новой для организма информации. Есть данные, что гиппокамп в совокупности
с медиальной частью височной доли играет
особую роль в процессе закрепления
(консолидации) следов памяти. Речь идет
о тех изменениях, которые происходят в
нервной ткани при образовании энграмм.
Гиппокамп, по-видимому, выполняет роль
селективного входного фильтра. Он
классифицирует все сигналы и отбрасывает
случайные, способствуя оптимальной
организации сенсорных следов в
долговременной памяти. Он также участвует
в извлечении следов из долговременной
памяти под влиянием мотивационного
возбуждения. Роль височной области
предположительно состоит в том, что она
устанавливает связь с местами хранения
следов памяти в других отделах мозга,
в первую очередь, в коре больших полушарий.
Другими словами, она отвечает за
реорганизацию нервных сетей в процессе
усвоения новых знаний; когда реорганизация
закончена, височная область в дальнейшем
процессе хранения участия не принимает.
Есть данные, что гиппокамп в совокупности
с медиальной частью височной доли играет
особую роль в процессе закрепления
(консолидации) следов памяти. Речь идет
о тех изменениях, которые происходят в
нервной ткани при образовании энграмм.
Гиппокамп, по-видимому, выполняет роль
селективного входного фильтра. Он
классифицирует все сигналы и отбрасывает
случайные, способствуя оптимальной
организации сенсорных следов в
долговременной памяти. Он также участвует
в извлечении следов из долговременной
памяти под влиянием мотивационного
возбуждения. Роль височной области
предположительно состоит в том, что она
устанавливает связь с местами хранения
следов памяти в других отделах мозга,
в первую очередь, в коре больших полушарий.
Другими словами, она отвечает за
реорганизацию нервных сетей в процессе
усвоения новых знаний; когда реорганизация
закончена, височная область в дальнейшем
процессе хранения участия не принимает.
На
третьем этапе следовые процессы переходят
в устойчивые структуры долговременной
памяти. Перевод информации из
кратковременной памяти в долговременную,
по некоторым предположениям может
происходить как во время бодрствования,
так и во сне (Ткаченко Б.И., 1994).
Перевод информации из
кратковременной памяти в долговременную,
по некоторым предположениям может
происходить как во время бодрствования,
так и во сне (Ткаченко Б.И., 1994).
В психологии этот процесс называется – сохранение. Когда нужно, мы воспроизводим ранее виденное, слышанное, пережитое. Именно по воспроизведению судят о качестве работы всего аппарата памяти.
Хорошая память – это способности быстро и много запоминать, точно и вовремя воспроизводить.
Кратковременная и долговременная память взаимосвязаны и работают как единая система. Одна из концепций, описывающих их совместную, взаимосвязанную деятельность, разработана американскими учеными Р.Аткинсоном и Р.Шифрином.
Кратковременная
память характеризуется ограниченностью
ее объема (в среднем 7±2). При переполнении
объема кратковременной памяти человека
вновь поступающая информация частично
вытесняет хранящуюся там, и последняя
безвозвратно исчезает. Кратковременная
память выступает в роли обязательного
промежуточного хранилища и фильтра,
перерабатывающего самый большой объем
информации, сразу отсеивающего ненужную
и оставляющего потенциально полезную.
Кратковременная
память выступает в роли обязательного
промежуточного хранилища и фильтра,
перерабатывающего самый большой объем
информации, сразу отсеивающего ненужную
и оставляющего потенциально полезную.
Процесс запоминания может протекать более эффективно, если сосредоточиться на усваиваемом материале. Установлено, что лучше усваивается информация, которая является объектом внимания и сознания, выступает в качестве цели. Таким образом, сокращается объем исходной информации, облегчается работа по ее переработке.
Другим мнемоническим приемом является запоминание путем повторения. Этот механизм основан на том, что запоминаемый материал посредством сознательного повторения удерживается в кратковременной памяти на более длительный срок, чем несколько секунд; повышается шанс перевода информации в долговременное хранилище. Обычно же без повторения в долговременной памяти оказывается лишь то, что находится в сфере внимания.
Одним
из возможных механизмов кратковременного
запоминания является временное
кодирование, то есть отражение
запоминаемого материала в виде
определенных, последовательно
расположенных знаков в слуховой и
зрительной системе человека. Как правило,
информация перекодируется в акустическую
форму, а затем сохраняется в долговременной
памяти в смысловой форме. Именно смысл
вспоминаемого приходит на память первым,
мы, в конечном счете, можем вспомнить
желаемое или по крайней мире заменить
его тем, что достаточно близко к нему
по смыслу. На этом, в частности, основан
процесс узнавания когда-то виденного
или слышанного.
Как правило,
информация перекодируется в акустическую
форму, а затем сохраняется в долговременной
памяти в смысловой форме. Именно смысл
вспоминаемого приходит на память первым,
мы, в конечном счете, можем вспомнить
желаемое или по крайней мире заменить
его тем, что достаточно близко к нему
по смыслу. На этом, в частности, основан
процесс узнавания когда-то виденного
или слышанного.
Особенностью долговременной памяти является то, что она, по мнению Р.Аткинсона и Р.Шифрина, практически неограниченна по объему и длительности хранения в ней информации.
Схема
памяти по Р.Аткинсону и Р.Шифрину
достаточно хорошо описывает работу
кратковременной памяти, но абсолютно
не учитывает обратную связь долговременной
памяти с кратковременной. Дело в том,
что оба типа памяти работают во взаимосвязи
и параллельно. В памяти идет постоянная
работа по обращению к прошлому опыту,
его дополнение новой информацией, а
также корректировка усвоенных сведений.
Иначе говоря, человеку нет необходимости
заучивать то, что он и так хорошо знает. На этом основана ассоциативная память
(Рубинштейн
С. Л., 1999).
На этом основана ассоциативная память
(Рубинштейн
С. Л., 1999).
Теория памяти, которая легла в основу первых классических экспериментальных исследований Г. Эббингауза и его продолжателей (Г. Э. Мюллера, А. Пильцекера, Ф. Шумана и др.), была целиком построена на учении об ассоциациях.
Существенным
в этой теории является то, что факт
внешней смежности впечатлений сам по
себе признается достаточным для
установления связи между представлениями
и для их воспроизведения. Исходя из
этого, Эббингауз и построил все свое
исследование. Он пользовался рядами
бессмысленных слогов, состоящих из трех
букв (одной гласной, расположенной между
двумя согласными, например туг — фал —
дор — сэт), с выключением всех тех
комбинаций, которые давали какое — либо
осмысленное слово. В подборе такого
материала Эббингауз руководился
стремлением получить однородный материал
и создать единообразные условия для
различных испытуемых. Отсутствие
осмысленного содержания в заучиваемом
материале и смысловых связей в нем было
для Эббингауза несущественно, потому
что для него процесс воспроизведения
определялся фактом создающей ассоциативные
связи внешней смежности заучиваемого
материала.
По мнению С.П.Рубинштейн, об этой классической теории, которая попыталась свести память к одним лишь ассоциативным связям, видно что, ассоциативные связи играют, несомненно, значительную роль, особенно в элементарных формах памяти. Однако, работа памяти в целом, особенно высшие формы памяти у человека, несводимы к одним лишь ассоциациям и не могут быть безостаточно объяснены ассоциативной теорией. Помимо ассоциативных связей по смежности, в работе человеческой памяти, в процессах запоминания, припоминания, воспроизведения существенную роль играют смысловые связи. Память человека носит осмысленный характер.
Исследования
(А. Бине, К. Бюлера и ряда других,
(Рубинштейн
С. Л., 1999),
отчетливо вскрыли роль осмысливания в
запоминании и показали, что, осмысленное
запоминание подчинено иным закономерностям,
чем механическое воспроизведение на
основе ассоциаций по смежности. При
воспроизведении осмысленного текста
его основные, наиболее существенные по
смыслу части воспроизводятся значительно
лучше, забывается по большей части
второстепенное, несущественное. Несущественное таким образом, отсеивается;
существенные же по смыслу части как бы
вычленяются из смежных с ними, но
существенно по смыслу с ними не связанных
частей, объединяясь в памяти с теми, с
которыми их связывает смысловой контекст.
Таким образом, вместо механического
воспроизведения смежных частей, которое
должно было бы иметь место согласно
законам ассоциации, фактически при
запоминании и воспроизведении осмысленного
текста происходит значительно более
сложный процесс смыслового отбора, в
результате которого преимущественно
закрепляется наиболее существенное
для данного субъекта, основной смысловой
остов текста. Сам текст, при этом
подвергается более или менее значительной
реконструкции. Воспроизведение в этих
случаях определяется не смежностью, а
может совершаться вопреки связям по
смежности, в соответствии со смысловыми
связями (Рубинштейн
С. Л., 1999).
Несущественное таким образом, отсеивается;
существенные же по смыслу части как бы
вычленяются из смежных с ними, но
существенно по смыслу с ними не связанных
частей, объединяясь в памяти с теми, с
которыми их связывает смысловой контекст.
Таким образом, вместо механического
воспроизведения смежных частей, которое
должно было бы иметь место согласно
законам ассоциации, фактически при
запоминании и воспроизведении осмысленного
текста происходит значительно более
сложный процесс смыслового отбора, в
результате которого преимущественно
закрепляется наиболее существенное
для данного субъекта, основной смысловой
остов текста. Сам текст, при этом
подвергается более или менее значительной
реконструкции. Воспроизведение в этих
случаях определяется не смежностью, а
может совершаться вопреки связям по
смежности, в соответствии со смысловыми
связями (Рубинштейн
С. Л., 1999).
Физиология, долговременная память — StatPearls
Книжная полка NCBI. Служба Национальной медицинской библиотеки, Национальных институтов здоровья.
StatPearls [Интернет]. Остров сокровищ (Флорида): StatPearls Publishing; 2023 янв.
StatPearls [Интернет].
Показать подробностиКритерий поиска
Алехандро Альмараз-Эспиноза; Майкл Х. Грайдер.
Информация об авторе и организацияхПоследнее обновление: 18 июля 2022 г.
Введение
Память — это сложная функция мозга, предназначенная для хранения и извлечения информации. У людей воспоминания о жизненном опыте в совокупности способствуют представлению о том, кто мы есть. Это функция, которая помогает нам взаимодействовать с окружающей средой; похожие ситуации вызывают определенные реакции, знакомые лица вызывают реакции, а повторяющиеся стимулы помогают нам учиться и реагировать.[1] Долговременные воспоминания делятся на две разные группы: процедурные и декларативные. Процедурная память включает в себя определенные действия, которым мы учимся, практикуясь и повторяя ряд двигательных действий (например, езда на велосипеде или вождение автомобиля). Декларативные воспоминания, которые могут быть сознательно вызваны, состоят из двух других групп: семантических воспоминаний, представляющих отдельные факты, такие как даты, определения слов и выученные понятия, и эпизодических воспоминаний, которые представляют явный опыт, который человек пережил, например, особый день рождения или день рождения. день свадьбы. В литературе иногда упоминаются эти две группы долговременных воспоминаний, неявные или явные, соответственно, хотя эти термины используются не так часто, потому что они более двусмысленны. Долговременная память объединяется от краткосрочных к долговременным воспоминаниям, в основном в гиппокампе и хранится по всей коре. Эти взаимосвязанные структуры помогают человеку интерпретировать стимулы, чтобы действовать соответствующим образом, либо извлекая старые воспоминания, чтобы помочь нам ориентироваться, либо сохраняя их, чтобы мы могли начать узнавать больше.[2][3][4][5][6]
Декларативные воспоминания, которые могут быть сознательно вызваны, состоят из двух других групп: семантических воспоминаний, представляющих отдельные факты, такие как даты, определения слов и выученные понятия, и эпизодических воспоминаний, которые представляют явный опыт, который человек пережил, например, особый день рождения или день рождения. день свадьбы. В литературе иногда упоминаются эти две группы долговременных воспоминаний, неявные или явные, соответственно, хотя эти термины используются не так часто, потому что они более двусмысленны. Долговременная память объединяется от краткосрочных к долговременным воспоминаниям, в основном в гиппокампе и хранится по всей коре. Эти взаимосвязанные структуры помогают человеку интерпретировать стимулы, чтобы действовать соответствующим образом, либо извлекая старые воспоминания, чтобы помочь нам ориентироваться, либо сохраняя их, чтобы мы могли начать узнавать больше.[2][3][4][5][6]
Сотовый уровень
Нейроны запускают электрические импульсы типа «все или ничего», называемые потенциалами действия, когда они стимулируются. Передачу сигналов нейронов, связанных с различными сенсорными модальностями, не так просто изучить; однако организация коры такова, что обработка сигналов происходит на нескольких уровнях в разных местах. В моделях приматов кодирование стимулов работает, когда группы нейронов срабатывают на основе определенных стимулов; чем конкретнее деталь, тем меньше нейронов задействовано. Например, меньшее количество нейронов отреагирует на распознавание лица вашей бабушки, чем на любое лицо человека или животного.[1][7][8][9]]
Передачу сигналов нейронов, связанных с различными сенсорными модальностями, не так просто изучить; однако организация коры такова, что обработка сигналов происходит на нескольких уровнях в разных местах. В моделях приматов кодирование стимулов работает, когда группы нейронов срабатывают на основе определенных стимулов; чем конкретнее деталь, тем меньше нейронов задействовано. Например, меньшее количество нейронов отреагирует на распознавание лица вашей бабушки, чем на любое лицо человека или животного.[1][7][8][9]]
Долговременная потенциация (ДП) была новаторским открытием 1960-х годов, которое определило молекулярную основу для формирования и хранения воспоминаний. В LTP, когда два нейрона повторно и одновременно активируются, прочность связи между этими клетками усиливается. Это усиление синапсов обычно называют «клетками, которые активируются вместе, соединяются вместе». Укрепление связей между нейронами помогает нам во многих отношениях, таких как распознавание мест, где мы были раньше, распознавание лиц, с которыми мы встречались, и информация, которую мы можем получить только при многократном воздействии.
Развитие
Результатом LTP является усиление синапсов нейронов за счет увеличения плотности рецепторов на постсинаптической клетке и экспрессии генов роста. Одним из наиболее изученных сигнальных путей, связанных с долговременной потенциацией и консолидацией памяти, является путь протеинкиназы А (ПКА). Непрерывная или повторяющаяся стимуляция определенных нервных связей (опосредованная реализацией повторяющихся стимулов, использованием определенных действий или наград) активирует рецепторы NMDA, которые обеспечивают приток ионов кальция. Этот приток инициирует пути вторичных мессенджеров, которые опосредованы PKA и циклическим аденозинмонофосфатом (цАМФ). Эти пути приводят к изменениям в синтезе белка и росте клеток, опосредованным белком, связывающим элемент ответа цАМФ (CREB), фактором транскрипции, который связывается с элементами ответа цАМФ (CRE). Связывание способствует транскрипции генов; это обычно демонстрировалось с использованием Aplysia , Drosophila и модели животных-грызунов. Рост клеток и синтез белка повышают пластичность нейронов и укрепляют связь между клетками.[14][8][9][15]
Рост клеток и синтез белка повышают пластичность нейронов и укрепляют связь между клетками.[14][8][9][15]
Вовлеченные системы органов
Центральная нервная система является основной вовлеченной системой. Определенные структуры мозга выполняют определенные задачи в рамках развития памяти. Например, гиппокамп необходим для объединения воспоминаний от краткосрочных к долгосрочным, тогда как миндалевидное тело придает воспоминаниям эмоциональную значимость. Именно структура и сила связей между корковыми структурами позволяют хранить и извлекать закодированную информацию.[15][1][16] Кодирование и хранение процедурных воспоминаний также затрагивают несколько областей мозга. Мозжечок, базальные ганглии и ассоциативная кора играют определенную роль в наших заученных действиях, поскольку они связаны с моторным контролем и приспособлением.[5]
Функциональная визуализация, такая как ПЭТ или фМРТ, позволила исследовать структуры, участвующие в кодировании стимула в декларативную память. Определенные области мозга могут иметь определенную функцию в консолидации воспоминаний. Тем не менее, большая часть коры головного мозга работает комбинированно, чтобы улучшить извлечение воспоминаний и обучение. «Рабочая память», временная память, которая не обязательно кодируется в долговременную память, определяется в литературе по-разному, причем некоторые авторы обсуждают рабочую память в тех же механизмах, что и долговременная память. Напротив, другие обсуждают это как другой и отдельный процесс хранения и обработки информации.
Определенные области мозга могут иметь определенную функцию в консолидации воспоминаний. Тем не менее, большая часть коры головного мозга работает комбинированно, чтобы улучшить извлечение воспоминаний и обучение. «Рабочая память», временная память, которая не обязательно кодируется в долговременную память, определяется в литературе по-разному, причем некоторые авторы обсуждают рабочую память в тех же механизмах, что и долговременная память. Напротив, другие обсуждают это как другой и отдельный процесс хранения и обработки информации.
Гиппокамп представляет собой структуру, связанную с зрительно-пространственной обработкой и частично с «рабочей» памятью. Возможны механизмы насыщения рабочей памяти, при этом избыточные детали или количество информации приводят к ошибкам в этой кратковременной памяти.[18] Разделение гиппокампа по продольной оси также обозначает определенные функции, например, задний гиппокамп связан с зрительно-пространственными деталями в определенных воспоминаниях, в то время как передний гиппокамп связан с запоминанием общего представления о месте. ] Гиппокамп также связан с эпизодической памятью, поскольку он помогает восстановить прожитые воспоминания и места, где они находились.[20]
] Гиппокамп также связан с эпизодической памятью, поскольку он помогает восстановить прожитые воспоминания и места, где они находились.[20]
Миндалевидное тело обрабатывает эмоциональные компоненты памяти, такие как боль, страх и удовольствие. Животные, стимулированные вредным или полезным стимулом, проявляют повышенную активность в областях миндалевидного тела, что позволяет животным различать положительные и отрицательные воспоминания. Отрицательные стимулы легче запоминаются и запоминаются, поскольку они, как правило, вызывают реакцию «бей или беги». У людей отрицательные стимулы, представленные во время формирования памяти, были связаны с тем, что субъекты сохраняли больше деталей о стимулах (изображения, слова и т. Д.). Поощрительные стимулы или «хорошие» стимулы также могут повышать удержание, но в меньшей степени.[21][22]
Вентральная префронтальная кора коррелирует с запоминанием заранее изученной схемы. Он имеет прочные связи с медиальными височными структурами и помогает распознавать то, что уже произошло, при этом запоминая некоторые связанные с этим детали, помогая заполнить пробелы в том, что, как нам кажется, мы уже знаем. [23]
[23]
Нервная система имеет глубокие связи с эндокринной системой. Например, высвобождение кортизола, нашего основного гормона стресса, инициируется передачей сигналов в гипоталамусе. Кроме того, существует корреляция между плотностью рецепторов кортизола, глюкокортикоидных рецепторов (ГР) в гиппокампе и способностью нейронных путей хранить информацию. Повышенная регуляция ГР и кортикоидов действует в синергии с пластичностью памяти и хранением памяти, что приводит к стрессовым ситуациям с более сильными воспоминаниями.
Клиническое значение
Корсаковский синдром — заболевание, опосредованное хроническим дефицитом витамина B1 (тиамина), обычно являющееся осложнением алкоголизма. Необратимые повреждения таких структур, как префронтальная кора и гиппокамп, могут изменить хранение информации. Конфабуляция является одним из наиболее распространенных признаков этого заболевания, когда пациент заполняет пробелы в воспоминаниях ложной информацией, которую он не может полностью вспомнить. [23]
[23]
У пациентов с повреждением медиальной височной доли, где находится гиппокамп, возникают проблемы с созданием новых воспоминаний. Эти пациенты, по-видимому, легко забывают или совершают ошибки даже после многократного получения подобных стимулов. Кроме того, у них нарушена кратковременная рабочая память. Например, пациенты с повреждением височной доли демонстрируют неспособность исправить предыдущие ошибки при многократном воздействии правильной информации. Авторы предполагают, что это связано с превышением объема рабочей памяти этих пациентов из-за предоставления слишком большого количества информации за короткое время удержания.
Болезнь Альцгеймера — наиболее распространенная форма деменции, обычно известная потерей кратковременной памяти и дегенерацией лобных и височных долей. Хотя пациенты склонны забывать, где они находятся, с кем они или что они делают в данный момент, они все же могут помнить определенные детали из своего прошлого. На ранних стадиях заболевания эпизодические и семантические воспоминания, а также процедурные воспоминания, как правило, сохранны. Семантическая память страдает больше, чем эпизодическая.[25][26]
Семантическая память страдает больше, чем эпизодическая.[25][26]
Дефицит памяти может проявляться при других типах распространенной деменции, таких как сосудистая деменция, болезнь Паркинсона и деменция с тельцами Леви. Сосудистая деменция может сохранить память или повлиять на нее. Это зависит от территории, пораженной инсультом или кровоизлиянием, так что процедурная память может быть более сохранной, чем декларативная память. Деменция, связанная с болезнью Паркинсона, и деменция с тельцами Леви являются двумя связанными типами деменции. Симптоматика и продолжительность симптомов различают эти заболевания. Деменция с тельцами Леви начинается со снижения когнитивных функций и галлюцинаций, тогда как болезнь Паркинсона начинается с моторных симптомов, таких как ригидность, брадикинезия и тремор. Нарушается кратковременная память, в то время как долговременная память имеет тенденцию демонстрировать относительную сохранность на ранних стадиях прогрессирования заболевания (включая эпизодическую и семантическую) [26].
Контрольные вопросы
Доступ к бесплатным вопросам с несколькими вариантами ответов по этой теме.
Комментарий к этой статье.
Ссылки
- 1.
Камина Э., Гуэль Ф. Нейроанатомические, нейрофизиологические и психологические основы памяти: современные модели и их происхождение. Фронт Фармакол. 2017;8:438. [Бесплатная статья PMC: PMC5491610] [PubMed: 28713278]
- 2.
Коуэн Н. Каковы различия между долговременной, кратковременной и рабочей памятью? Прог Мозг Res. 2008;169:323-38. [Бесплатная статья PMC: PMC2657600] [PubMed: 18394484]
- 3.
Норрис Д. Кратковременная память и долговременная память все же разные. Психологический бык. 2017 сен; 143 (9): 992-1009. [Бесплатная статья PMC: PMC5578362] [PubMed: 28530428]
- 4.
Дженесон А., Сквайр Л.Р. Рабочая память, долговременная память и функция медиальной височной доли. Выучить Мем.
2012 янв;19(1):15-25. [Бесплатная статья PMC: PMC3246590] [PubMed: 22180053]- 5.
Люм Дж. А., Конти-Рамсден Г., Пейдж Д., Ульман М.Т. Рабочая, декларативная и процессуальная память при специфических языковых нарушениях. кора. 2012 Октябрь;48(9):1138-54. [Бесплатная статья PMC: PMC3664921] [PubMed: 21774923]
- 6.
Müller NC, Genzel L, Konrad BN, Pawlowski M, Neville D, Fernández G, Steiger A, Dresler M. Motor Skills Enhance Pro Формирование процедурной памяти и защитить от возрастного ухудшения. ПЛОС Один. 2016;11(6):e0157770. [Бесплатная статья PMC: PMC4917083] [PubMed: 27333186]
- 7.
Suthana NA, Parikshak NN, Ekstrom AD, Ison MJ, Knowlton BJ, Bookheimer SY, Fried I. Специфические реакции нейронов гиппокампа человека связаны с лучшей памятью. Proc Natl Acad Sci U S A. 18 августа 2015 г .; 112 (33): 10503-8. [Бесплатная статья PMC: PMC4547223] [PubMed: 26240357]
- 8.
Kandel ER, Dudai Y, Mayford MR.
Молекулярная и системная биология памяти. Клетка. 2014 27 марта; 157 (1): 163-86. [PubMed: 24679534]- 9.
Бисаз Р., Травалья А., Альберини К.М. Нейробиологические основы формирования памяти: от физиологических состояний к психопатологии. Психопатология. 2014;47(6):347-56. [Бесплатная статья PMC: PMC4246028] [PubMed: 25301080]
- 10.
Лёмо Т. Открытие долговременного потенцирования. Philos Trans R Soc Lond B Biol Sci. 2003 29 апреля; 358 (1432): 617-20. [Бесплатная статья PMC: PMC1693150] [PubMed: 12740104]
- 11.
Такеучи Т., Душкевич А.Дж., Моррис Р.Г. Гипотеза синаптической пластичности и памяти: кодирование, хранение и постоянство. Philos Trans R Soc Lond B Biol Sci. 2014 05 января; 369(1633): 20130288. [Бесплатная статья PMC: PMC3843897] [PubMed: 24298167]
- 12.
Lynch MA. Долговременная потенциация и память. Physiol Rev. 2004 г., январь; 84 (1): 87–136. [PubMed: 14715912]
- 13.
Моррис Р.Г. Долговременная потенциация и память. Philos Trans R Soc Lond B Biol Sci. 2003 29 апреля; 358 (1432): 643-7. [Бесплатная статья PMC: PMC1693171] [PubMed: 12740109]
- 14.
Kandel ER. Молекулярная биология памяти: цАМФ, PKA, CRE, CREB-1, CREB-2 и CPEB. Мол Мозг. 2012 14 мая; 5:14. [Бесплатная статья PMC: PMC3514210] [PubMed: 22583753]
- 15.
Rolls ET, Treves A. Нейронное кодирование информации в головном мозге. Прог Нейробиол. 2011 ноябрь; 95 (3): 448-90. [PubMed: 21907758]
- 16.
Хабер С.Н. Кортико-стриарная схема. Диалоги Clin Neurosci. 2016 март; 18(1):7-21. [Бесплатная статья PMC: PMC4826773] [PubMed: 27069376]
- 17.
Baddeley A. Рабочая память. Наука. 1992 г., 31 января; 255 (5044): 556-9. [PubMed: 1736359]
- 18.
Pastötter B, Tempel T, Bäuml KT. Обновление долговременной памяти: гипотеза сброса кодирования в забывании, направленном по списку.
Фронт Псих. 2017;8:2076. [Бесплатная статья PMC: PMC5711817] [PubMed: 29230187]- 19.
Small SA, Nava AS, Perera GM, DeLaPaz R, Mayeux R, Stern Y. Цепные механизмы, лежащие в основе кодирования и извлечения памяти по длинной оси образование гиппокампа. Нат Нейроски. 2001 г., апрель; 4(4):442-9. [PubMed: 11276237]
- 20.
Москович М., Кабеса Р., Винокур Г., Надель Л. Эпизодическая память и не только: гиппокамп и неокортекс в трансформации. Анну Рев Психол. 2016;67:105-34. [Бесплатная статья PMC: PMC5060006] [PubMed: 26726963]
- 21.
Kensinger EA, Addis DR, Atapattu RK. Активность миндалевидного тела при кодировании соответствует яркости воспоминаний и памяти на отдельные эпизодические детали. Нейропсихология. 2011 март; 49(4):663-73. [Бесплатная статья PMC: PMC3060942] [PubMed: 21262244]
- 22.
Фернандо А.Б., Мюррей Дж.Э., Милтон А.Л. Миндалевидное тело: обеспечение удовольствия и избегание боли.
Фронт Behav Neurosci. 2013 06 декабря; 7:190. [Бесплатная статья PMC: PMC3854486] [PubMed: 24367307]- 23.
Оскар-Берман М. Функция и дисфункция префронтальной мозговой схемы при алкогольном Корсаковском синдроме. Neuropsychol Rev. 2012 Jun; 22 (2): 154–69. [Бесплатная статья PMC: PMC3681949] [PubMed: 22538385]
- 24.
Conrad CD. ВЗАИМОСВЯЗЬ МЕЖДУ ОСТРЫМИ УРОВНЯМИ ГЛЮКОКОРТИКОИДОВ И ФУНКЦИЕЙ ГИППОКАМПА ЗАВИСИТ ОТ АВЕРСИВНОСТИ ЗАДАЧИ И СТАДИИ ОБРАБОТКИ ПАМЯТИ. Нелинейность Biol Toxicol Med. 2005;3(1):57-78. [Бесплатная статья PMC: PMC1431575] [PubMed: 16601824]
- 25.
Ян Х. Потеря памяти при болезни Альцгеймера. Диалоги Clin Neurosci. 2013 Декабрь; 15 (4): 445-54. [Бесплатная статья PMC: PMC3898682] [PubMed: 24459411]
- 26.
Duong S, Patel T, Chang F. Деменция: что нужно знать фармацевтам. Кан Фарм Дж. (Отт). 2017 март-апрель;150(2):118-129. [Бесплатная статья PMC: PMC5384525] [PubMed: 28405256]
 2012 янв;19(1):15-25. [Бесплатная статья PMC: PMC3246590] [PubMed: 22180053]
2012 янв;19(1):15-25. [Бесплатная статья PMC: PMC3246590] [PubMed: 22180053] Молекулярная и системная биология памяти. Клетка. 2014 27 марта; 157 (1): 163-86. [PubMed: 24679534]
Молекулярная и системная биология памяти. Клетка. 2014 27 марта; 157 (1): 163-86. [PubMed: 24679534]
 Фронт Псих. 2017;8:2076. [Бесплатная статья PMC: PMC5711817] [PubMed: 29230187]
Фронт Псих. 2017;8:2076. [Бесплатная статья PMC: PMC5711817] [PubMed: 29230187] Фронт Behav Neurosci. 2013 06 декабря; 7:190. [Бесплатная статья PMC: PMC3854486] [PubMed: 24367307]
Фронт Behav Neurosci. 2013 06 декабря; 7:190. [Бесплатная статья PMC: PMC3854486] [PubMed: 24367307]Раскрытие информации: Алехандро Альмараз-Эспиноза заявляет об отсутствии соответствующих финансовых отношений с неправомочными компаниями.
Раскрытие информации: Майкл Грайдер заявляет об отсутствии соответствующих финансовых отношений с неправомочными компаниями.

Эта книга распространяется на условиях Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0) ( http://creativecommons.org/licenses/by-nc-nd/4.0/ ), что позволяет другим распространять произведение при условии, что статья не изменена и не используется в коммерческих целях. Вам не требуется получать разрешение на распространение этой статьи при условии, что вы указываете автора и журнал.
ID на книжной полке: NBK549791PMID: 31747198
Нейробиологические основы формирования памяти: от физиологических состояний к психопатологии
1. Кандель Э.Р. Молекулярная биология хранения памяти: диалог между генами и синапсами. Наука. 2001; 294:1030–1038. [PubMed] [Google Scholar]
2. Дудай Ю. Нейробиология консолидаций, или насколько стабильна инграмма? Ежегодный обзор психологии. 2004; 55: 51–86. [PubMed] [Google Scholar]
2004; 55: 51–86. [PubMed] [Google Scholar]
3. Lamprecht R, Farb CR, Rodrigues SM, LeDoux JE. Обусловливание страха направляет профилин в дендритные шипы миндалевидного тела. Нат Нейроски. 2006;9: 481–483. [PubMed] [Google Scholar]
4. Альберини CM. Роль синтеза белка во время лабильных фаз памяти: пересмотр скептицизма. Нейробиол Узнать Мем. 2008; 89: 234–246. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Alberini CM. Факторы транскрипции в долговременной памяти и синаптической пластичности. Physiol Rev. 2009; 89: 121–145. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Davis HP, Squire LR. Синтез белка и память: обзор. Психологический бюллетень. 1984;96:518–559. [PubMed] [Google Scholar]
7. Граф П., Шактер Д.Л. Имплицитная и эксплицитная память на новые ассоциации у нормальных и страдающих амнезией субъектов. Журнал экспериментальной психологии Обучение, память и познание. 1985; 11: 501–518. [PubMed] [Google Scholar]
8. Squire LR, Zola SM.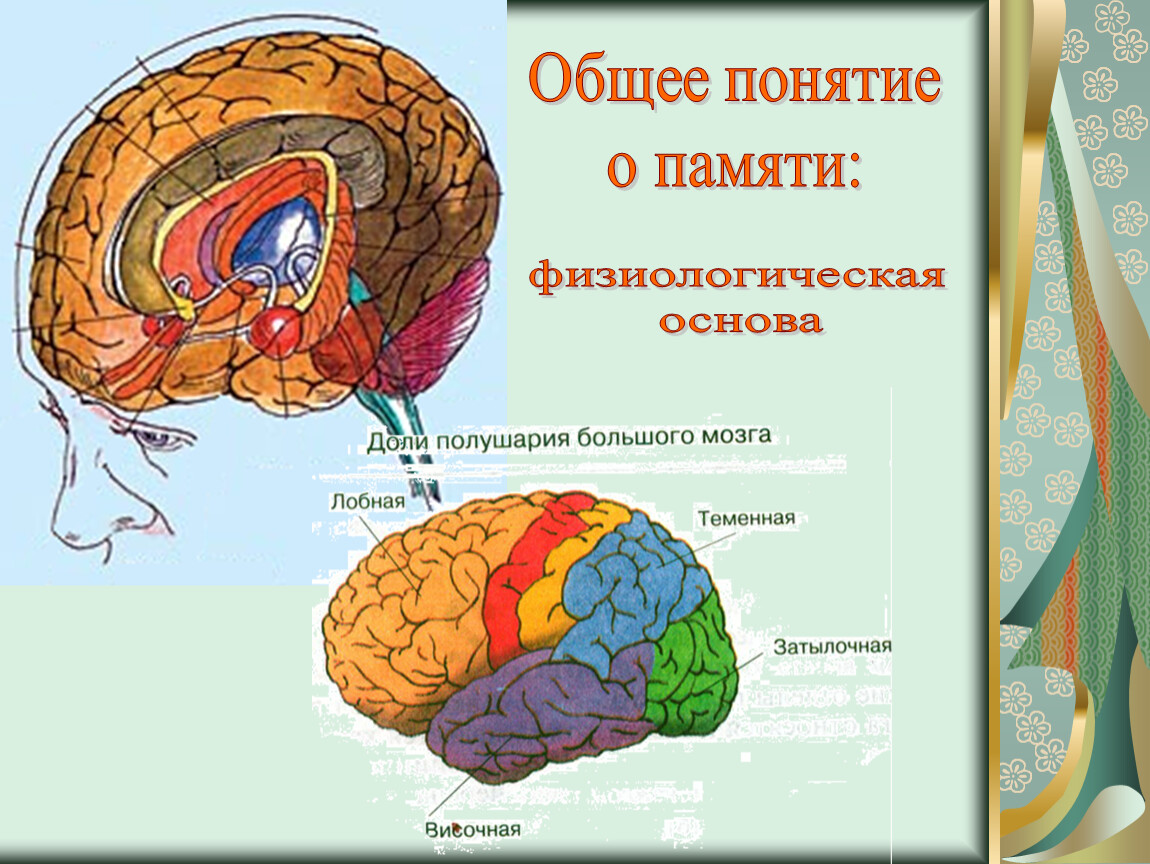 Структура и функции декларативной и недекларативной систем памяти. Proc Natl Acad Sci U S A. 1996; 93:13515–13522. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Структура и функции декларативной и недекларативной систем памяти. Proc Natl Acad Sci U S A. 1996; 93:13515–13522. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Карони П., Донато Ф., Мюллер Д. Структурная пластичность при обучении: регуляция и функции. Нат Рев Нейроски. 2012; 13: 478–49.0. [PubMed] [Google Scholar]
10. Frankland PW, Bontempi B. Организация недавних и отдаленных воспоминаний. Нат Рев Нейроски. 2005; 6: 119–130. [PubMed] [Google Scholar]
11. Дудай Ю. Беспокойная инграмма: Консолидации никогда не заканчиваются. Ежегодный обзор неврологии. 2012; 35: 227–247. [PubMed] [Google Scholar]
12. Рамамурти Б. Четвертое состояние сознания: турия авастха. Психиатрия и клиническая неврология. 1995; 49: 107–110. [PubMed] [Академия Google]
13. Дудай Ю., Айзенберг М. Ритуалы прохождения инграммы: реконсолидация и гипотеза затяжной консолидации. Нейрон. 2004; 44: 93–100. [PubMed] [Google Scholar]
14. McGaugh JL. Память — столетие консолидации. Наука. 2000; 287: 248–251. [PubMed] [Google Scholar]
Наука. 2000; 287: 248–251. [PubMed] [Google Scholar]
15. Alberini CM, Ledoux JE. Реконсолидация памяти. Карр Биол. 2013; 23: Р746–750. [PubMed] [Google Scholar]
16. Надер К., Эйнарссон Э.О. Реконсолидация памяти: обновление. Энн Н.Ю. Академия наук. 2010;1191:27–41. [PubMed] [Google Scholar]
17. Tronson NC, Taylor JR. Молекулярные механизмы реконсолидации памяти. Нат Рев Нейроски. 2007; 8: 262–275. [PubMed] [Google Scholar]
18. Сара SJ. Поиск и реконсолидация: к нейробиологии запоминания. Выучить Мем. 2000;7:73–84. [PubMed] [Google Scholar]
19. Альберини CM. Роль реконсолидации и динамический процесс формирования и хранения долговременной памяти. Фронт Behav Neurosci. 2011;5:12. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Питман РК. Предложит ли блокада реконсолидации новое лечение посттравматического стрессового расстройства? Фронт Behav Neurosci. 2011;5:11. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Hartley CA, Phelps EA. Изменение страха: нейросхема регуляции эмоций. Нейропсихофармакол. 2010; 35: 136–146. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Изменение страха: нейросхема регуляции эмоций. Нейропсихофармакол. 2010; 35: 136–146. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Schafe GE, Nader K, Blair HT, LeDoux JE. Консолидация памяти павловского кондиционирования страха: клеточная и молекулярная перспектива. Тренды Нейроси. 2001; 24: 540–546. [PubMed] [Академия Google]
23. Сквайр Л.Р., Викстед Дж.Т. Когнитивная нейронаука человеческой памяти с тех пор, как Х.М. Ежегодный обзор неврологии. 2011; 34: 259–288. [Бесплатная статья PMC] [PubMed] [Google Scholar]
24. Аргуэлло А.А., Йе Х., Боздаги О., Поллонини Г., Тронель С., Бамбах-Мукку Д., Хантли Г.В., Платано Д., Альберини К.М. Дельта белка, связывающего энхансер Ccaat, играет существенную роль в консолидации и реконсолидации памяти. Дж. Нейроски. 2013; 33:3646–3658. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Milekic MH, Pollonini G, Alberini CM. Временная потребность c/ebpbeta в миндалевидном теле после реактивации, но не приобретения ингибиторного избегания. Выучить Мем. 2007; 14: 504–511. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Выучить Мем. 2007; 14: 504–511. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Таубенфельд С.М., Милекич М.Х., Монти Б., Альберини К.М. Для консолидации новой, но не реактивированной памяти требуется c/ebpbeta гиппокампа. Нат Нейроски. 2001; 4: 813–818. [PubMed] [Google Scholar]
27. Кандель Э.Р. Молекулярная биология памяти: Camp, pka, cre, creb-1, creb-2 и cpeb. Молекулярный мозг. 2012;5:14. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Alberini CM, Chen DY. Улучшение памяти: консолидация, реконсолидация и инсулиноподобный фактор роста 2. Trends Neurosci. 2012 [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Стерн С.А., Альберини С.М. Механизмы улучшения памяти. Междисциплинарные обзоры Wiley по системной биологии и медицине. 2013; 5:37–53. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. de Kloet ER, Joels M, Holsboer F. Стресс и мозг: от адаптации к болезни. Нат Рев Нейроски. 2005; 6: 463–475. [PubMed] [Google Scholar]
31. de Quervain DJ, Aerni A, Schelling G, Roozendaal B. Глюкокортикоиды и регуляция памяти в норме и при болезни. Передний нейроэндокринол. 2009 г.;30:358–370. [PubMed] [Google Scholar]
de Quervain DJ, Aerni A, Schelling G, Roozendaal B. Глюкокортикоиды и регуляция памяти в норме и при болезни. Передний нейроэндокринол. 2009 г.;30:358–370. [PubMed] [Google Scholar]
32. Сэнди С., Рихтер-Левин Г. От черты высокой тревожности к депрессии: нейрокогнитивная гипотеза. Тренды Нейроси. 2009; 32: 312–320. [PubMed] [Google Scholar]
33. Питман Р.К., Расмуссон А.М., Коенен К.С., Шин Л.М., Орр С.П., Гилбертсон М.В., Милад М.Р., Либерзон И. Биологические исследования посттравматического стрессового расстройства. Нат Рев Нейроски. 2012;13:769–787. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. McEwen BS, Gianaros PJ. Пластичность мозга, вызванная стрессом и аллостазом. Ежегодный обзор медицины. 2011;62:431–445. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Кооб Г., Крик М.Дж. Стресс, нарушение регуляции путей вознаграждения за наркотики и переход к наркотической зависимости. Am J Психиатрия. 2007; 164:1149–1159. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Roozendaal B, McGaugh JL. Модуляция памяти. Поведение Нейроски. 2011; 125:797–824. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Roozendaal B, McGaugh JL. Модуляция памяти. Поведение Нейроски. 2011; 125:797–824. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Okuda S, Roozendaal B, McGaugh JL. Влияние глюкокортикоидов на память распознавания объектов требует связанного с обучением эмоционального возбуждения. Proc Natl Acad Sci U S A. 2004; 101:853–858. [Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Санди С, Пинело-Нава, МТ. Стресс и память: поведенческие эффекты и нейробиологические механизмы. Нейр Пласт. 2007; 2007:78970. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Revest JM, Di Blasi F, Kitchener P, Rouge-Pont F, Desmedt A, Turiault M, Tronche F, Piazza PV. Путь mapk и egr-1 опосредуют связанные со стрессом поведенческие эффекты глюкокортикоидов. Нат Нейроски. 2005; 8: 664–672. [PubMed] [Google Scholar]
40. Revest JM, Kaouane N, Mondin M, Le Roux A, Rouge-Pont F, Vallee M, Barik J, Tronche F, Desmedt A, Piazza PV. Улучшение связанной со стрессом памяти глюкокортикоидами зависит от синапсин-ia/ib. Мол Психиатрия. 2010;15:1125, 1140–1151. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мол Психиатрия. 2010;15:1125, 1140–1151. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Чен Д.Ю., Бамбах-Мукку Д., Поллонини Г., Альберини К.М. Глюкокортикоидные рецепторы задействуют пути camkiialpha-bdnf-creb для обеспечения консолидации памяти. Нат Нейроски. 2012;15:1707–1714. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Bilang-Bleuel A, Ulbricht S, Chandramohan Y, De Carli S, Droste SK, Reul JM. Психологический стресс увеличивает фосфорилирование гистона h4 в нейронах гранул зубчатой извилины взрослых: участие в поведенческом ответе, зависящем от глюкокортикоидных рецепторов. Евр Джей Нейроски. 2005;22:1691–1700. [PubMed] [Google Scholar]
43. Чандрамохан Ю., Дросте С.К., Артур Дж.С., Реул Дж.М. Реакция поведенческой неподвижности, вызванная вынужденным плаванием, включает фосфоацетилирование гистона h4 и индукцию c-fos в нейронах гранул зубчатой извилины посредством активации n-метил-d-аспартатной/внеклеточной сигнал-регулируемой киназы/митоген- и стресс-активируемой киназы. путь. Евр Джей Нейроски. 2008; 27: 2701–2713. [PubMed] [Google Scholar]
путь. Евр Джей Нейроски. 2008; 27: 2701–2713. [PubMed] [Google Scholar]
44. Розендаль Б., Эрнандес А., Кабрера С.М., Хагевуд Р., Мальваез М., Стефанко Д.П., Хеттиг Дж., Вуд М.А. Связанная с мембраной глюкокортикоидная активность необходима для модуляции долговременной памяти посредством модификации хроматина. Дж. Нейроски. 2010;30:5037–5046. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Финстервальд С., Альберини С.М. Стресс и механизмы, зависящие от глюкокортикоидных рецепторов, в долговременной памяти: от адаптивных реакций до психопатологий. Нейробиол Узнать Мем. 2013 [бесплатная статья PMC] [PubMed] [Google Scholar]
46. Allen SJ, Dawbarn D. Клиническая значимость нейротрофинов и их рецепторов. Clin Sci (Лондон) 2006; 110: 175–191. [PubMed] [Google Scholar]
47. Думан Р.С., Хенингер Г.Р., Нестлер Э.Дж. Молекулярно-клеточная теория депрессии. Арх генерал психиатрия. 1997;54:597–606. [PubMed] [Google Scholar]
48. Думан Р.С., Монтеджиа Л. М. Нейротрофическая модель расстройств настроения, связанных со стрессом. Биол психиатрия. 2006;59:1116–1127. [PubMed] [Google Scholar]
М. Нейротрофическая модель расстройств настроения, связанных со стрессом. Биол психиатрия. 2006;59:1116–1127. [PubMed] [Google Scholar]
49. Pariante CM, Lightman SL. Ось hpa при большой депрессии: классические теории и новые разработки. Тренды Нейроси. 2008; 31: 464–468. [PubMed] [Google Scholar]
50. Nemeroff CB, Vale WW. Нейробиология депрессии: пути к лечению и открытие новых лекарств. Дж. Клин Психиатрия. 2005; 66 (Приложение 7): 5–13. [PubMed] [Академия Google]
51. Чен Б., Довлатшахи Д., МакКуин Г.М., Ван Д.Ф., Янг Л.Т. Повышенная иммунореактивность bdnf гиппокампа у субъектов, получавших антидепрессанты. Биол психиатрия. 2001; 50: 260–265. [PubMed] [Google Scholar]
52. Dwivedi Y, Rizavi HS, Conley RR, Roberts RC, Tamminga CA, Pandey GN. Измененная экспрессия гена нейротрофического фактора головного мозга и рецепторной тирозинкиназы b в посмертном мозге суицидентов. Архив общей психиатрии. 2003; 60: 804–815. [PubMed] [Академия Google]
53. Karege F, Vaudan G, Schwald M, Perroud N, La Harpe R. Уровни нейротрофинов в посмертном мозге жертв самоубийств и эффекты прижизненной диагностики и психотропных препаратов. Исследования мозга Молекулярные исследования мозга. 2005; 136: 29–37. [PubMed] [Google Scholar]
Karege F, Vaudan G, Schwald M, Perroud N, La Harpe R. Уровни нейротрофинов в посмертном мозге жертв самоубийств и эффекты прижизненной диагностики и психотропных препаратов. Исследования мозга Молекулярные исследования мозга. 2005; 136: 29–37. [PubMed] [Google Scholar]
54. Karege F, Perret G, Bondolfi G, Schwald M, Bertschy G, Aubry JM. Снижение уровня нейротрофического фактора головного мозга в сыворотке крови у пациентов с тяжелой депрессией. Психиатрия рез. 2002; 109: 143–148. [PubMed] [Академия Google]
55. Парианте СМ. Глюкокортикоидный рецептор: часть решения или часть проблемы? Журнал психофармакологии. 2006; 20:79–84. [PubMed] [Google Scholar]
56. Nader K, Schafe GE, Le Doux JE. Воспоминания о страхе требуют синтеза белка в миндалевидном теле для реконсолидации после извлечения. Природа. 2000;406:722–726. [PubMed] [Google Scholar]
57. Альберини CM. Механизмы стабилизации памяти: консолидация и реконсолидация похожи или разные процессы? Тренды Нейроси. 2005; 28:51–56. [PubMed] [Академия Google]
2005; 28:51–56. [PubMed] [Академия Google]
58. Милекич М.Х., Альберини СМ. Временная потребность в синтезе белка после реактивации памяти. Нейрон. 2002; 36: 521–525. [PubMed] [Google Scholar]
59. Suzuki A, Josselyn SA, Frankland PW, Masushige S, Silva AJ, Kida S. Реконсолидация и угасание памяти имеют различные временные и биохимические признаки. Дж. Нейроски. 2004; 24:4787–4795. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Frankland PW, Ding HK, Takahashi E, Suzuki A, Kida S, Silva AJ. Стабильность недавних и отдаленных контекстуальных воспоминаний о страхе. Выучить Мем. 2006; 13: 451–457. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Графф Дж., Джозеф Н.Ф., Хорн М.Е., Самиэй А., Мэн Дж., Сео Дж., Рей Д., Беро А.В., Фан Т.Х., Вагнер Ф., Холсон Э., Сюй Дж., Сунь Дж., Нив Р.Л., Мах Р.Х., Хаггарти SJ, Цай ЛХ. Эпигенетическое праймирование обновления памяти во время реконсолидации для ослабления отдаленных воспоминаний о страхе. Клетка. 2014; 156: 261–276. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2014; 156: 261–276. [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Debec J, LeDoux JE, Nader K. Клеточная и системная реконсолидация в гиппокампе. Нейрон. 2002; 36: 527–538. [PubMed] [Google Scholar]
63. Alberini CM, Johnson SA, Ye X. Механизмы и функции реконсолидации памяти. В: Альберини CM, редактор. Реконсолидация памяти. Том. 1. Академическая пресса; Сан-Диего: 2013. стр. 81–117. [Академия Google]
64. Tronel S, Milekic MH, Alberini CM. Связывание новой информации с реактивированной памятью требует механизмов консолидации, а не реконсолидации. PLoS биол. 2005;3:e293. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Хупбах А., Гомес Р., Надель Л. Реконсолидация эпизодической памяти: обновление или путаница с источником? Объем памяти. 2009; 17: 502–510. [PubMed] [Google Scholar]
66. Хупбах А., Хардт О., Гомес Р., Надель Л. Динамика памяти: контекстно-зависимое обновление. Выучить Мем. 2008; 15: 574–579.. [PubMed] [Google Scholar]
67. Morris RG, Inglis J, Ainge JA, Olverman HJ, Tulloch J, Dudai Y, Kelly PA. Реконсолидация памяти: чувствительность пространственной памяти к ингибированию синтеза белка в дорсальном гиппокампе во время кодирования и поиска. Нейрон. 2006; 50: 479–489. [PubMed] [Google Scholar]
Morris RG, Inglis J, Ainge JA, Olverman HJ, Tulloch J, Dudai Y, Kelly PA. Реконсолидация памяти: чувствительность пространственной памяти к ингибированию синтеза белка в дорсальном гиппокампе во время кодирования и поиска. Нейрон. 2006; 50: 479–489. [PubMed] [Google Scholar]
68. Forcato C, Argibay PF, Pedreira ME, Maldonado H. Реконсолидация человека не всегда происходит при восстановлении памяти: актуальность структуры напоминания. Нейробиол Узнать Мем. 2009 г.;91:50–57. [PubMed] [Google Scholar]
69. Forcato C, Rodriguez ML, Pedreira ME, Maldonado H. Реконсолидация у людей открывает декларативную память для поступления новой информации. Нейробиол Узнать Мем. 2010;93:77–84. [PubMed] [Google Scholar]
70. Taubenfeld SM, Riceberg JS, New AS, Alberini CM. Доклиническая оценка избирательного нарушения травматической памяти посредством постретривного ингибирования глюкокортикоидных рецепторов. Биол психиатрия. 2009; 65: 249–257. [Бесплатная статья PMC] [PubMed] [Google Scholar]
71. Голье Дж.А., Караманика К., Демария Р., Йехуда Р. Пилотное исследование мифепристона при посттравматическом синдроме, связанном с боевыми действиями. Исследования и лечение депрессии. 2012;2012:393251. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Голье Дж.А., Караманика К., Демария Р., Йехуда Р. Пилотное исследование мифепристона при посттравматическом синдроме, связанном с боевыми действиями. Исследования и лечение депрессии. 2012;2012:393251. [Бесплатная статья PMC] [PubMed] [Google Scholar]
72. de Quervain DJ, Henke K, Aerni A, Treyer V, McGaugh JL, Berthold T, Nitsch RM, Buck A, Roozendaal B, Hock C. Глюкокортикоид-индуцированный нарушение декларативной памяти связано со снижением кровотока в медиальных отделах височной доли. Евр Джей Нейроски. 2003; 17: 1296–1302. [PubMed] [Академия Google]
73. Sandi C. Глюкокортикоиды действуют на глутаматергические пути, влияя на процессы памяти. Тренды Нейроси. 2011; 34: 165–176. [PubMed] [Google Scholar]
74. Аэрни А., Трабер Р., Хок С., Рузендал Б., Шеллинг Г., Папасситиропулос А., Нич Р.М., Шнайдер У., де Кервен Д.Дж. Низкие дозы кортизола при симптомах посттравматического стрессового расстройства. Am J Психиатрия. 2004; 161:1488–1490. [PubMed] [Google Scholar]
75. Соравиа Л.М., Генрихс М., Эрни А., Марони С., Шеллинг Г., Элерт У., Рузендал Б., де Кервен Д.Дж. Глюкокортикоиды уменьшают фобический страх у людей. Proc Natl Acad Sci USA. 2006; 103:5585–559.0. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Соравиа Л.М., Генрихс М., Эрни А., Марони С., Шеллинг Г., Элерт У., Рузендал Б., де Кервен Д.Дж. Глюкокортикоиды уменьшают фобический страх у людей. Proc Natl Acad Sci USA. 2006; 103:5585–559.0. [Бесплатная статья PMC] [PubMed] [Google Scholar]
76. de Quervain DJ, Bentz D, Michael T, Bolt OC, Wiederhold BK, Margraf J, Wilhelm FH. Глюкокортикоиды усиливают психотерапию, основанную на угашении. Proc Natl Acad Sci U S A. 2011;108:6621–6625. [Бесплатная статья PMC] [PubMed] [Google Scholar]
77. Hill MN, McEwen BS. Участие эндоканнабиноидной системы в нейроповеденческих эффектах стресса и глюкокортикоидов. Прог Нейропсихофармакол Биол Психиатрия. 2010; 34: 791–797. [Бесплатная статья PMC] [PubMed] [Google Scholar]
78. Hill MN, Patel S, Campolongo P, Tasker JG, Wotjak CT, Bains JS. Функциональные взаимодействия между стрессом и эндоканнабиноидной системой: от синаптической сигнализации до поведенческого результата. Дж. Нейроски. 2010;30:14980–14986. [Бесплатная статья PMC] [PubMed] [Google Scholar]
79. Steiner MA, Wotjak CT. Роль эндоканнабиноидной системы в регуляции гипоталамо-гипофизарно-адренокортикальной оси. Прог Мозг Res. 2008; 170:397–432. [PubMed] [Google Scholar]
Steiner MA, Wotjak CT. Роль эндоканнабиноидной системы в регуляции гипоталамо-гипофизарно-адренокортикальной оси. Прог Мозг Res. 2008; 170:397–432. [PubMed] [Google Scholar]
80. Морена М., Камполонго П. Эндоканнабиноидная система: эмоциональный буфер в модуляции функции памяти. Нейробиология обучения и памяти. 2014 [PubMed] [Академия Google]
81. Линь Х.К., Мао С.К., Гин П.В. Влияние инфузии агонистов рецептора cb1 внутрь миндалевидного тела на реконсолидацию испуга, вызванного вздрагиванием. Выучить Мем. 2006; 13: 316–321. [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Campolongo P, Roozendaal B, Trezza V, Hauer D, Schelling G, McGaugh JL, Cuomo V. Эндоканнабиноиды в базолатеральной миндалине крысы улучшают консолидацию памяти и обеспечивают модуляцию глюкокортикоидов. памяти. Proc Natl Acad Sci U S A. 2009;106:4888–4893. [Бесплатная статья PMC] [PubMed] [Google Scholar]
83. Лавиолетт С.Р., Грейс А.А. Каннабиноиды усиливают пластичность эмоционального обучения в нейронах медиальной префронтальной коры через входы базолатеральной миндалевидного тела. Дж. Нейроски. 2006; 26: 6458–6468. [Бесплатная статья PMC] [PubMed] [Google Scholar]
84. Tan H, Lauzon NM, Bishop SF, Bechard MA, Laviolette SR. Интегрированная передача каннабиноидного рецептора cb1 в миндалевидно-префронтальном кортикальном пути модулирует пластичность нейронов и кодирование эмоциональной памяти. Кора головного мозга. 2010; 20:1486–149.6. [PubMed] [Google Scholar]
85. Tan H, Lauzon NM, Bishop SF, Chi N, Bechard M, Laviolette SR. Передача каннабиноидов в базолатеральной миндалевидном теле модулирует формирование памяти о страхе через функциональные входы в прелимбическую кору. Дж. Нейроски. 2011;31:5300–5312. [Бесплатная статья PMC] [PubMed] [Google Scholar]
86. Pamplona FA, Bitencourt RM, Takahashi RN. Краткосрочные и долгосрочные эффекты каннабиноидов на угасание контекстуальной памяти о страхе у крыс. Нейробиол Узнать Мем. 2008;90:290–293. [PubMed] [Google Scholar]
87. Памплона Ф.А., Предигер Р.Д., Пандольфо П., Такахаши Р.Н. Агонист каннабиноидных рецепторов win 55,212-2 способствует угасанию контекстуальной памяти о страхе и пространственной памяти у крыс. Психофармакология (Берл) 2006; 188: 641–649. [PubMed] [Google Scholar]
Психофармакология (Берл) 2006; 188: 641–649. [PubMed] [Google Scholar]
88. Чхатвал Дж.П., Дэвис М., Магущак К.А., Ресслер К.Дж. Усиление каннабиноидной нейротрансмиссии усиливает угасание условного страха. Нейропсихофармакол. 2005; 30: 516–524. [PubMed] [Академия Google]
89. Izquierdo A, Wellman CL, Holmes A. Кратковременный неконтролируемый стресс вызывает ретракцию дендритов в инфралимбической коре и устойчивость к исчезновению страха у мышей. Дж. Нейроски. 2006; 26: 5733–5738. [Бесплатная статья PMC] [PubMed] [Google Scholar]
90. Rabinak CA, Angstadt M, Sripada CS, Abelson JL, Liberzon I, Milad MR, Phan KL. Каннабиноидное облегчение воспоминаний об исчезновении страха у людей. Нейрофармакология. 2013; 64: 396–402. [Бесплатная статья PMC] [PubMed] [Google Scholar]
91. Izquierdo I, Quillfeldt JA, Zanatta MS, Quevedo J, Schaeffer E, Schmitz PK, Medina JH. Последовательная роль гиппокампа и миндалевидного тела, энторинальной коры и теменной коры в формировании и восстановлении памяти для торможения избегания у крыс. Евр Джей Нейроски. 1997; 9: 786–793. [PubMed] [Google Scholar]
Евр Джей Нейроски. 1997; 9: 786–793. [PubMed] [Google Scholar]
92. Лутц Б. Эндоканнабиноидная система и обучение вымиранию. Мол Нейробиол. 2007; 36: 92–101. [PubMed] [Google Scholar]
93. Hill MN, Gorzalka BB. Эндоканнабиноидная система и лечение аффективных и тревожных расстройств. Лекарственные мишени для нейролептиков ЦНС. 2009 г.;8:451–458. [PubMed] [Google Scholar]
94. Trezza V, Campolongo P. Эндоканнабиноидная система как возможная мишень для лечения как когнитивных, так и эмоциональных особенностей посттравматического стрессового расстройства (ПТСР). Фронт Behav Neurosci. 2013;7:100. [Бесплатная статья PMC] [PubMed] [Google Scholar]
95. Monfils MH, Cowansage KK, Klann E, LeDoux JE. Границы угасания-реконсолидации: ключ к стойкому ослаблению воспоминаний о страхе. Наука. 2009; 324:951–955. [Бесплатная статья PMC] [PubMed] [Google Scholar]
96. Schiller D, Monfils MH, Raio CM, Johnson DC, Ledoux JE, Phelps EA. Предотвращение возвращения страха у людей с помощью механизмов обновления реконсолидации. Природа. 2010; 463:49–53. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Природа. 2010; 463:49–53. [Бесплатная статья PMC] [PubMed] [Google Scholar]
97. Jones CE, Monfils MH. Использование реконсолидации и угасания для ослабления воспоминаний о страхе на животных моделях. В: Альберини CM, редактор. Реконсолидация памяти. Том. 1. Академическая пресса; Сан-Диего: 2013. стр. 165–184. [Google Scholar]
98. Фелпс Э.А., Шиллер Д. Реконсолидация у человека. В: Альберини CM, редактор. Реконсолидация памяти. Том. 1. Академическая пресса; Сан-Диего: 2013. стр. 185–212. [Академия Google]
99. Xue YX, Luo YX, Wu P, Shi HS, Xue LF, Chen C, Zhu WL, Ding ZB, Bao YP, Shi J, Epstein DH, Shaham Y, Lu L. Процедура извлечения-угасания памяти для предотвратить тягу к наркотикам и рецидивы. Наука. 2012; 336: 241–245. [Бесплатная статья PMC] [PubMed] [Google Scholar]
100. van Oostrom I, Dollfus S, Brazo P, Abadie P, Halbecq I, Thery S, Marie RM. Вербальное обучение и память у больных шизофренией и болезнью Паркинсона. Психиатрия рез. 2003; 117: 25–34. [PubMed] [Академия Google]
[PubMed] [Академия Google]
101. Чирилло М.А., Сейдман Л.Дж. Вербальная декларативная дисфункция памяти при шизофрении: от клинической оценки до генетики и механизмов мозга. Обзор нейропсихологии. 2003; 13:43–77. [PubMed] [Google Scholar]
102. Тулопулуанд Т., Мюррей Р.М. Дефицит вербальной памяти у пациентов с шизофренией: важная будущая цель лечения. Эксперт преподобный Нейротер. 2004; 4:43–52. [PubMed] [Google Scholar]
103. Стефан К.Е., Балдевег Т., Фристон К.Дж. Синаптическая пластичность и разъединение при шизофрении. Биол психиатрия. 2006; 59: 929–939. [PubMed] [Google Scholar]
104. Favalli G, Li J, Belmonte-de-Abreu P, Wong AH, Daskalakis ZJ. Роль bdnf в патофизиологии и лечении шизофрении. J Psychiatr Res. 2012; 46:1–11. [PubMed] [Google Scholar]
105. Ангелуччи Ф., Брене С., Мате А.А. Bdnf при шизофрении, депрессии и соответствующих животных моделях. Молекулярная психиатрия. 2005; 10: 345–352. [PubMed] [Google Scholar]
106. Muglia P, Vicente AM, Verga M, King N, Macciardi F, Kennedy JL. Ассоциация между геном bdnf и шизофренией. Молекулярная психиатрия. 2003; 8: 146–147. [PubMed] [Академия Google]
Ассоциация между геном bdnf и шизофренией. Молекулярная психиатрия. 2003; 8: 146–147. [PubMed] [Академия Google]
107. Отри А.Е., Монтеджиа Л.М. Нейротрофический фактор головного мозга и нервно-психические расстройства. Фармакологические обзоры. 2012; 64: 238–258. [Бесплатная статья PMC] [PubMed] [Google Scholar]
108. Вытилингам М., Хейм С., Ньюпорт Дж., Миллер А.Х., Андерсон Э., Бронен Р., Браммер М., Стаиб Л., Верметтен Э., Чарни Д.С., Немерофф С.Б., Бремнер Джей Ди. Детская травма, связанная с меньшим объемом гиппокампа у женщин с большой депрессией. Am J Психиатрия. 2002;159:2072–2080. [Бесплатная статья PMC] [PubMed] [Google Scholar]
109. Хэкман Д.А., Фарах М.Дж., Мини М.Дж. Социально-экономический статус и мозг: механистические выводы из исследований человека и животных. Нат Рев Нейроски. 2010; 11: 651–659. [Бесплатная статья PMC] [PubMed] [Google Scholar]
110. Pratchett LC, Yehuda R. Основы посттравматического стрессового расстройства: приводит ли ранняя жизненная травма к посттравматическому стрессовому расстройству у взрослых? Дев психопат. 2011; 23: 477–491. [PubMed] [Google Scholar]
2011; 23: 477–491. [PubMed] [Google Scholar]
111. Фергюссон Д.М., Лински М., Хорвуд Л.Дж. Подростковые результаты усыновления: 16-летнее продольное исследование. Журнал детской психологии и психиатрии и смежных дисциплин. 1995;36:597–615. [PubMed] [Google Scholar]
112. Беккет С., Моган Б., Раттер М., Касл Дж., Колверт Э., Грутьюз С., Креппнер Дж., Стивенс С., О’Коннор Т.Г., Сонуга-Барк Э.Дж. Сохраняются ли последствия ранней тяжелой депривации на когнитивные функции в раннем подростковом возрасте? Результаты исследования усыновленных детей из Англии и Румынии. Развитие ребенка. 2006; 77: 696–711. [PubMed] [Google Scholar]
113. Sanchez MM, Ladd CO, Plotsky PM. Ранний неблагоприятный опыт как фактор риска развития более поздней психопатологии: данные моделей грызунов и приматов. Дев психопат. 2001;13:419–449. [PubMed] [Google Scholar]
114. Heim C, Plotsky PM, Nemeroff CB. Важность изучения вклада раннего неблагоприятного опыта в нейробиологические данные о депрессии. Нейропсихофармакол. 2004; 29: 641–648. [PubMed] [Google Scholar]
Нейропсихофармакол. 2004; 29: 641–648. [PubMed] [Google Scholar]
115. Hanson M, Godfrey KM, Lillycrop KA, Burdge GC, Gluckman PD. Пластичность развития и происхождение неинфекционных заболеваний: теоретические соображения и эпигенетические механизмы. Прогресс в биофизике и молекулярной биологии. 2011; 106: 272–280. [PubMed] [Академия Google]
116. Чжан Т.И., Лабонте Б., Вэнь С.Л., Турецки Г., Мини М.Дж. Эпигенетические механизмы ранней регуляции окружающей средой экспрессии генов глюкокортикоидных рецепторов гиппокампа у грызунов и человека. Нейропсихофармакол. 2013; 38:111–123. [Бесплатная статья PMC] [PubMed] [Google Scholar]
117. Szyf M, Weaver IC, Champagne FA, Diorio J, Meaney MJ. Материнское программирование экспрессии и фенотипа стероидных рецепторов посредством метилирования ДНК у крыс. Передний нейроэндокринол. 2005; 26: 139–162. [PubMed] [Академия Google]
118. Шампанское Ф.А., Керли Дж.П. Эпигенетические механизмы, опосредующие долгосрочное влияние материнской заботы на развитие. Neurosci Biobehav Rev. 2009; 33: 593–600. [PubMed] [Google Scholar]
Neurosci Biobehav Rev. 2009; 33: 593–600. [PubMed] [Google Scholar]
119. Фрейд С. Три эссе по теории сексуальности. В: Strachey TJ, редактор. Стандартное издание полного собрания психологических сочинений Зигмунда Фрейда. Том. 7. Хогарт Пресс; 1953. [Google Scholar]
120. Кэмпбелл Б.А., Спир Н.Е. Онтогенез памяти. Psychol Rev. 1972; 79: 215–&. [PubMed] [Академия Google]
121. Копье Н.Е., Парсонс П. Анализ реактивационного лечения: Онтогенные детерминанты облегченного забывания. Лоуренс Эрлбаум Ассошиэйтс; Hillsdale, New Jersey: 1976. [Google Scholar]
122. Hayne H. Развитие детской памяти: последствия детской амнезии. Dev Rev. 2004; 24:33–73. [Google Scholar]
123. Терр Л. Что происходит с ранними воспоминаниями о травме? Исследование двадцати детей в возрасте до пяти лет во время задокументированных травмирующих событий. Журнал Американской академии детской и подростковой психиатрии. 1988;27:96–104. [PubMed] [Google Scholar]
124. Gaensbauer TJ.