Понятие о генотипе и фенотипе.
Генотип— вся совокупность имеющихся у организма генов.
Фенотип— совокупность реализованных (т.е. внешних) генетически детерминированных признаков, т.е. индивидуальное (в определенных условиях внешней среды) проявление генотипа. При изменении условий существования фенотип бактерий изменяется при сохранении генотипа.
Изменчивость у бактерий может быть ненаследуемой (модификационной) и генотипической (мутации, рекомбинации).
Временные, наследственно не закрепленные изменения, возникающие как адаптивные реакции бактерий на изменения окружающей среды, называются модификациями (чаще — морфологические и биохимические модификации). После устранения причины бактерии реверсируют к исходному фенотипу.
Стандартное
проявление модификации — распределение
однородной популяции на две или более
двух типов- диссоциация. Пример- характер
роста на питательных средах: S- (гладкие)
колонии, R- (шероховатые) колонии, M-
(мукоидные, слизистые) колонии, D-
(карликовые) колонии.
Мутации — скачкообразные изменения наследственного признака. Могут быть спонтанные и индуцированные, генные (изменения одного гена) и хромосомные (изменения двух или более двух участков хромосомы).
Одновременно у бактерий имеются различные механизмы репарации мутаций, в том числе с использованием ферментов — эндонуклеаз, лигаз, ДНК- полимеразы.
Генетические рекомбинации — изменчивость, связанная с обменом генетической информации. Генетические рекомбинации могут осуществляться путем трансформации, трансдукции, конъюгации, слияния протопластов.
1.Трансформация — захват и поглощение фрагментов чужой ДНК и образование на этой основе рекомбинанта.
2.Трансдукция
— перенос генетического материала фагами
(умеренными фагами — специфическая
трансдукция).
3.Конъюгация — при непосредственном контакте клеток. Контролируется tra (transfer) опероном. Главную роль играют конъюгативные F- плазмиды.
Схема бактериальной трансформации и трансдукции
Трансформация — перенос генетического материала, заключающийся в том, что бактерия-реципиент захватывает (поглощает) из внешней среды фрагменты чужеродной ДНК.
Трансдукция — перенос генетического
материала от клетки-донора клетке-реципиенту
с помощью бактериофагов.
клетка-донор образует F-пили.
2— F-пили прикрепляются к клетке-реципиенту.
3— Мобильная плазмида разрывается и одна цепь ДНК перемещается в клетку-реципиент.

4— В обеих клетках синтезируется вторая цепь ДНК и образуются F-пили. Обе клетки могут быть донорами.

Геном вирусов содержит или РНК, или ДНК (РНК- и ДНК- вирусы соответственно). Выделяют позитивную (+) РНК, обладающую матричной активностью и соответственно- инфекционными свойствами, и негативную ( — ) РНК, не проявляющую инфекционные свойства, которая для воспроизводства должна транскрибироваться (превращаться) в + РНК. Механизмы репродукции различных вирусов очень сложные и существенно отличаются. Основные их схематические варианты представлены ниже:
1. вирионная (матричная) +РНК комплементарная -РНК (в рибосомах) вирионная +РНК.
2. — РНК вирусная (информационная) +РНК — РНК (формируется на геноме зараженной клетки).
3. однонитевая ДНК: +ДНК +ДНК -ДНК +ДНК -ДНК +ДНК +ДНК.
4.
ретровирусная однонитевая РНК: РНК
ДНК (провирус) РНК.
5. двунитевая ДНК: разделение нитей ДНК и формирование на каждой комплементарной нити ДНК.
Генофонд вирусов создается и пополняется из четырех основных источников:
двух внутренних (мутации, рекомбинации) и двух внешних (включение в геном генетического материала клетки хозяина, поток генов из других вирусных популяций).
Комплементация
Фенотипическое смешивание — при заражении клетки близкородственными вирусами с образованием вирионов с гибридными капсидами, кодируемыми геномами двух вирусов.
Популяционная
изменчивость вирусов связана с двумя
разнонаправленными процессами — мутациями
и селекцией, связанными с внешней средой
как индуктором мутаций и фактором
стабилизирующего отбора. Гетерогенность
вирусных популяций — адаптационный
генетический механизм, способствующий
пластичности (устойчивости,
приспособляемости) популяций, фактор
эволюции и сохранения видов во внешней
среде.
Генофонд вирусных популяций сохраняется за счет нескольких механизмов:
— восстановления изменчивости за счет мутаций;
— резервирующих механизмов (возможность перехода любых, даже негативных мутаций в следующую генерацию)- комплементация, рекомбинация;— буферных механизмов (образование дефектных вирусных частиц, иммунных комплексов и др.), способствующие сохранению вируса в изменяющихся внешних условиях.
Гены: словарь терминов
ВАЖНО!
Информацию из данного раздела нельзя использовать для самодиагностики и самолечения. В случае боли или иного обострения заболевания диагностические исследования должен назначать только лечащий врач. Для постановки диагноза и правильного назначения лечения следует обращаться к Вашему лечащему врачу.
Аллели – (от греч. allelon друг друга, взаимно) различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологичных (парных) хромосом, контролирующие один и тот же белок.
Аутосомно-доминантный тип наследования – мутантный аллель (вариант) доминирует над нормальным аллелем (вариантом), т.е. проявляется как в гомозиготном, так и в гетерозиготном состоянии; патологическая наследственность прослеживается в родословной «по вертикали»; по крайней мере, один из родителей имеет проявление данной мутации. При этом мутантный ген расположен в аутосоме (неполовой хромосоме) и наследование не сцеплено с полом.
Аутосомно-рецессивный тип наследования – нормальный аллель (вариант) подавляет проявление мутантного аллеля (варианта), т.е. мутация может проявиться, только находясь в гомозиготном состоянии. При этом мутантный ген расположен в аутосоме (неполовой хромосоме) и наследование не сцеплено с полом.
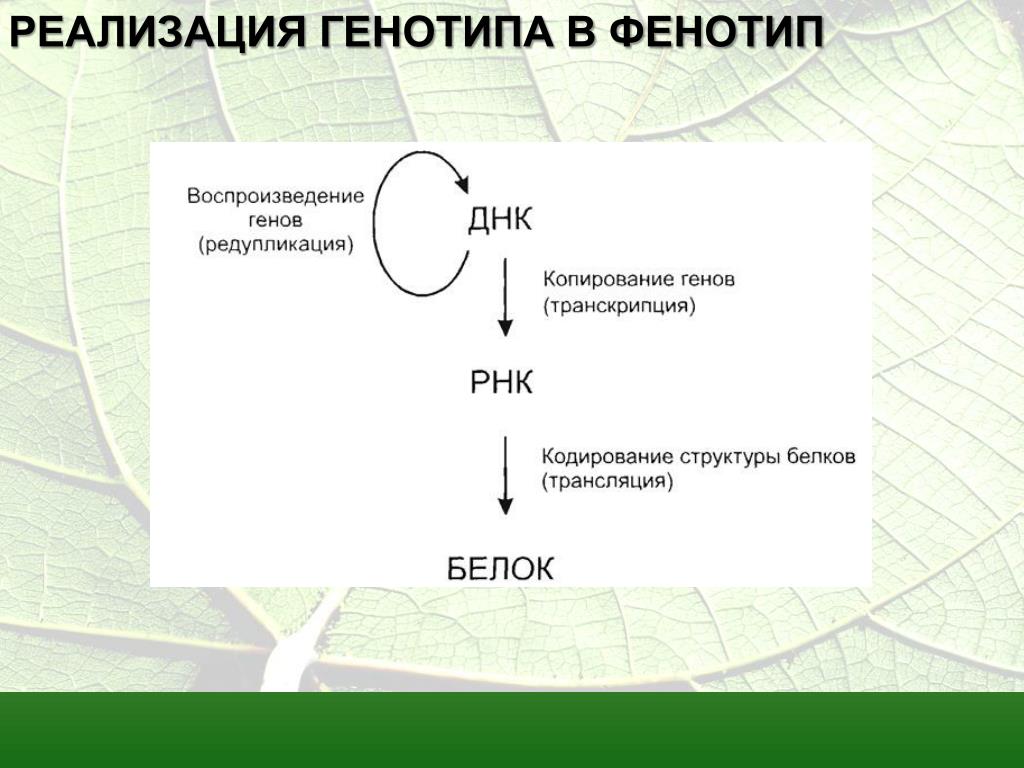
Ген – элементарная единица наследственности, наименьший неделимый элемент наследственного материала, который может быть передан от родителей потомству как целое и который определяет признаки, свойства или физиологическую функцию организма. На молекулярном уровне — это участок молекулы ДНК, кодирующий первичную структуру белков и РНК.
На молекулярном уровне — это участок молекулы ДНК, кодирующий первичную структуру белков и РНК.
Генетический вариант полиморфизма – в широком смысле аналогичен понятию аллеля; здесь, в узком смысле – одна из двух разновидностей гена, различающихся по одному генетическому полиморфизму. Генетические варианты полиморфизма обычно отличаются аминокислотной последовательностью белкового продукта гена или уровнем экспрессии гена.
Генный полиморфизм, полиморфизм (здесь, в узком смысле) – структурное различие альтернативных вариантов гена (обычно нормального и мутантного). Возникновение вариантов гена обусловлено мутациями. Применительно к понятию «генный полиморфизм» обычно рассматриваются нейтральные мутации, не приводящие к заметным нарушениям функции гена, тогда как «мутациями» обычно называют изменения в гене, приводящие к выраженному нарушению работы гена.
Генотип – (здесь, в узком смысле) сочетание генетических вариантов (аллелей), расположенных на гомологичных хромосомах.
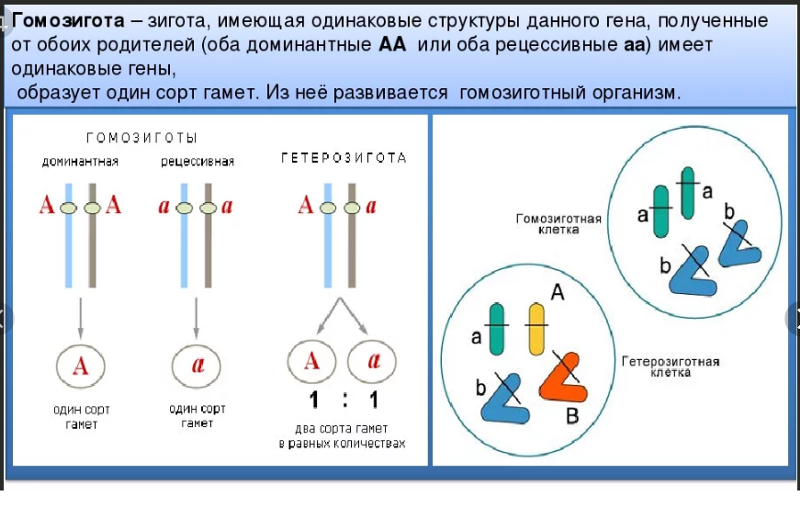
Гетерозиготный – содержащий разные аллели (генетические варианты) в соответствующих локусах гомологичных хромосом.
Гомозиготный – содержащий одинаковые аллели (генетические варианты) в соответствующих локусах гомологичных хромосом.
Гомологичные хромосомы – парные хромосомы из диплоидного набора, одинаковые по форме, размерам и набору генов.
Делеция – утрата в результате мутации сегмента ДНК размером от одного нуклеотида до субхромосомного фрагмента, включающего несколько генов. В случае генных полиморфизмов рассматриваются делеции, ограниченные одним геном.
Диплоидный набор – набор хромосом в соматических клетках организма, который содержит два гомологичных набора хромосом, из которых один передается от одного родителя, а второй — от другого.
Доминантный аллель – аллель, проявляющийся в фенотипе гетерозиготных особей.
Инсерция – вставка сегмента ДНК размерами от одного нуклеотида до субхромосомного фрагмента, включающего несколько генов. В случае генных полиморфизмов рассматриваются инсерции, ограниченные одним геном.
Интрон – область гена, разделяющая экзоны и не несущая информации об аминокислотной последовательности белкового продукта.
Миссенс-мутация — мутация, приводящая к подстановке несоответствующей аминокислоты в полипептидную цепь.
Мультифакториальные заболевания – заболевания, вызываемые взаимодействием множества наследственных и внешних факторов, например, ишемическая болезнь сердца, инфаркт миокарда (ИМ), инсульт, некоторые формы рака, психические заболевания и др.
Мутантный вариант полиморфизма – вариант полиморфизма, возникший вследствие мутации из своего предшественника – нормального варианта.
Мутация – изменение нуклеотидной последовательности ДНК. Чаще всего мутации представляют собой однонуклеотидные замены – миссенс-мутации. Влияние мутации на функцию гена может варьировать от ее полного нарушения до незначительного влияния или отсутствия влияния.
Нормальный вариант полиморфизма – наиболее распространенный в популяции вариант полиморфизма, который является предшественником других вариантов, возникающих вследствие мутаций.
Нуклеотид – структурная единица нуклеиновых кислот. В состав ДНК входят 4 нуклеотида: аденин – А, тимин – Т, гуанин – G, цитозин – С.
Пенетрантность – количественный показатель фенотипической изменчивости проявления гена. Измеряется (обычно в %) отношением числа особей, у которых данный ген проявился в фенотипе, к общему числу особей, в генотипе которых этот ген присутствует в необходимом для его проявления состоянии (гомозиготном – в случае рецессивных генов или гетерозиготном – в случае доминантных генов). Проявление гена у 100% особей с соответствующим генотипом называется полной пенетрантностью, в остальных случаях – неполной пенетрантностью. Неполная пенетрантность свойственна проявлению генов, связанных с мультифакториальными заболеваниями: болезнь развивается только у части лиц, в генотипе которых присутствует аномальный ген; у остальных же наследственное предрасположение к болезни остаётся нереализованным.
Полиморфизм гена – многообразие нуклеотидных последовательностей гена, в том числе его аллельных форм.
Прогностический – понятие, характеризующее заключение о предстоящем развитии и исходе, основанное на специальном исследовании.
Промотор – участок молекулы ДНК, к которому присоединяются молекулы РНК-полимеразы, что сопровождается инициацией транскрипции соответствующих генов; как правило, промотор расположен на операторном конце оперона; каждый ген (или оперон) имеет свой промотор, контролирующий его транскрипцию.
Рецессивный аллель – аллель, не проявляющийся в фенотипе гетерозиготных особей.
Фенотип – особенности строения и жизнедеятельности организма, обусловленные взаимодействием его генотипа с условиями среды.
Хромосома – составной элемент клеточного ядра, являющийся носителем генов. В основе хромосомы лежит линейная молекула ДНК
Экзон – фрагмент гена, кодирующий аминокислотную последовательность белкового продукта данного гена.
Экспрессия гена – перенос генетической информации от ДНК через РНК к полипептидам и белкам в определенных типах клеток организма.
ВАЖНО!
Информацию из данного раздела нельзя использовать для самодиагностики и самолечения. В случае боли или иного обострения заболевания диагностические исследования должен назначать только лечащий врач. Для постановки диагноза и правильного назначения лечения следует обращаться к Вашему лечащему врачу.
Генетические алгоритмы — математический аппарат
Генетические алгоритмы предназначены для решения задач оптимизации и моделирования путём последовательного подбора, комбинирования и вариации искомых параметров с использованием механизмов, напоминающих биологическую эволюцию.
Генетические алгоритмы предназначены для решения задач оптимизации. Примером подобной задачи может служить обучение нейросети, то есть подбора таких значений весов, при которых достигается минимальная ошибка. При этом в основе генетического алгоритма лежит метод случайного поиска. Основным недостатком случайного поиска является то, что нам неизвестно сколько понадобится времени для решения задачи.
Для того, чтобы избежать таких расходов времени при решении задачи, применяются методы, открытые при изучении эволюции и происхождения видов. Как известно, в процессе эволюции выживают наиболее приспособленные особи. Это приводит к тому, что приспособленность популяции возрастает, позволяя ей лучше выживать в изменяющихся условиях.
Впервые подобный алгоритм был предложен в 1975 году Джоном Холландом (John Holland) в Мичиганском университете. Он получил название «репродуктивный план Холланда» и лег в основу практически всех вариантов генетических алгоритмов. Однако, перед тем, как мы его рассмотрим подробнее, необходимо остановится на том, каким образом объекты реального мира могут быть закодированы для использования в генетических алгоритмах.
Представление объектов
Из биологии мы знаем, что любой организм может быть представлен своим фенотипом, который фактически определяет, чем является объект в реальном мире, и генотипом, который содержит всю информацию об объекте на уровне хромосомного набора. При этом каждый ген, то есть элемент информации генотипа, имеет свое отражение в фенотипе.
При этом каждый ген, то есть элемент информации генотипа, имеет свое отражение в фенотипе.
Таким образом, для решения задач нам необходимо представить каждый признак объекта в форме, подходящей для использования в генетическом алгоритме. Все дальнейшее функционирование механизмов генетического алгоритма производится на уровне генотипа, позволяя обойтись без информации о внутренней структуре объекта, что и обуславливает его широкое применение в самых разных задачах.
В наиболее часто встречающейся разновидности генетического алгоритма для представления генотипа объекта применяются битовые строки. При этом каждому атрибуту объекта в фенотипе соответствует один ген в генотипе объекта. Ген — битовая строка, чаще всего фиксированной длины, которая представляет собой значение этого признака.
Кодирование признаков (целые числа)
Для кодирования таких признаков можно использовать самый простой вариант — битовое значение этого признака. Тогда нам будет весьма просто использовать ген определенной длины, достаточной для представления всех возможных значений такого признака. Но, к сожалению, такое кодирование не лишено недостатков.
Но, к сожалению, такое кодирование не лишено недостатков.
Основной недостаток заключается в том, что соседние числа отличаются в значениях нескольких битов, так например числа 7 и 8 в битовом представлении различаются в 4-х позициях, что затрудняет функционирование генетического алгоритма и увеличивает время, необходимое для его сходимости. Для того, чтобы избежать эту проблему лучше использовать кодирование, при котором соседние числа отличаются меньшим количеством позиций, в идеале значением одного бита.
Таким кодом является код Грея, который целесообразно использовать в реализации генетического алгоритма. Значения кодов Грея рассмотрены в таблице ниже:
| Десятичный код | Двоичное значение | Шестнадцатеричное значение | Десятичный код | Двоичное значение | Шестнадцатеричное значение |
|---|---|---|---|---|---|
| 0 | 0000 | 0h | 0 | 0000 | 0h |
| 1 | 0001 | 1h | 1 | 0001 | 1h |
| 2 | 0010 | 2h | 3 | 0011 | 3h |
| 3 | 0011 | 3h | 2 | 0010 | 2h |
| 4 | 0100 | 4h | 6 | 0110 | 6h |
| 5 | 0101 | 5h | 7 | 0111 | 7h |
| 6 | 0110 | 6h | 5 | 0101 | 5h |
| 7 | 0111 | 7h | 4 | 0100 | 4h |
| 8 | 1000 | 8h | 12 | 1100 | Ch |
| 9 | 1001 | 9h | 13 | 1101 | Dh |
| 10 | 1010 | Ah | 15 | 1111 | Fh |
| 11 | 1011 | Bh | 14 | 1110 | Eh |
| 12 | 1100 | Ch | 10 | 1010 | Ah |
| 13 | 1101 | Dh | 11 | 1011 | Bh |
| 14 | 1110 | Eh | 9 | 1001 | 9h |
| 15 | 1111 | Fh | 8 | 1000 | 8h |
Таблица 1. Соответствие десятичных кодов (слева) и кодов Грея (справа)
Соответствие десятичных кодов (слева) и кодов Грея (справа)
Таким образом, при кодировании целочисленного признака мы разбиваем его на тетрады и каждую тетраду преобразуем по коду Грея.
В практических реализациях генетических алгоритмов обычно не возникает необходимости преобразовывать значения признака в значение гена. На практике имеет место обратная задача, когда по значению гена необходимо определить значение соответствующего ему признака.
Таким образом, задача декодирования значения генов, которым соответствуют целочисленные признаки, тривиальна.
Кодирование признаков (действительные числа)
Самый простой способ кодирования, который лежит на поверхности – использовать битовое представление, хотя такой вариант имеет те же недостатки, что и для целых чисел. Поэтому на практике обычно применяют следующую последовательность действий:
- Разбивают весь интервал допустимых значений признака на участки с требуемой точностью.
- Принимают значение гена как целочисленное число, определяющее номер интервала (используя код Грея).
- В качестве значения параметра принимают число, являющиеся серединой этого интервала.

Рассмотрим вышеописанную последовательность действий на примере.
Допустим, что значения признака лежат в интервале [0, 1]. При кодировании использовалось разбиение участка на 256 интервалов. Для кодирования их номера нам потребуется, таким образом, 8 бит. Допустим, значение гена: 00100101bG (заглавная буква G показывает, что используется кодирование по коду Грея). Для начала, используя код Грея, найдем соответствующий ему номер интервала:
25hG->36h->54d.
Теперь посмотрим, какой интервал ему соответствует… После несложных подсчетов получаем интервал [0.20703125, 0.2109375]. Значит значение нашего параметра будет (0.20703125+0.2109375)/2=0.208984375.
Кодирование нечисловых данных
При кодировании нечисловых данных необходимо предварительно преобразовать их в числа.
Определение фенотипа по генотипу
Для того чтобы определить фенотип объекта (то есть значения признаков, описывающих объект) нам необходимо только знать значения генов, соответствующим этим признакам, то есть генотип объекта. При этом совокупность генов, описывающих генотип объекта, представляет собой хромосому. В некоторых реализациях ее также называют особью.
При этом совокупность генов, описывающих генотип объекта, представляет собой хромосому. В некоторых реализациях ее также называют особью.
Таким образом, в реализации генетического алгоритма хромосома представляет собой битовую строку фиксированной длины. При этом каждому участку строки соответствует ген. Длина генов внутри хромосомы может быть одинаковой или различной. Чаще всего применяют гены одинаковой длины.
Рассмотрим пример хромосомы и интерпретации ее значения. Допустим, что у объекта имеется 5 признаков, каждый закодирован геном длинной в 4 элемента. Тогда длина хромосомы будет 5*4=20 бит:
| 0010 | 1010 | 1001 | 0100 | 1101 |
|---|
Теперь мы можем определить значения признаков:
| Признак | Значение гена | Двоичное значение признака | Десятичное значение признака |
|---|---|---|---|
| 1 | 0010 | 0011 | 3 |
| 2 | 1010 | 1100 | 12 |
| 3 | 1001 | 1110 | 14 |
| 4 | 0100 | 0111 | 7 |
| 5 | 1101 | 1001 | 9 |
Основные генетические операторы
Как известно, в теории эволюции важную роль играет то, каким образом признаки родителей передаются потомкам. В генетических алгоритмах за передачу признаков родителей потомкам отвечает оператор, который называется скрещивание (его также называют кроссовер или кроссинговер). Этот оператор определяет передачу признаков родителей потомкам.
В генетических алгоритмах за передачу признаков родителей потомкам отвечает оператор, который называется скрещивание (его также называют кроссовер или кроссинговер). Этот оператор определяет передачу признаков родителей потомкам.
Действует он следующим образом:
- из популяции выбираются две особи, которые будут родителями;
- определяется (обычно случайным образом) точка разрыва;
- потомок определяется как конкатенация части первого и второго родителя.
Рассмотрим функционирование этого оператора:
| Хромосома_1 | 0000000000 |
| Хромосома_2 | 1111111111 |
Допустим, разрыв происходит после 3-го бита хромосомы, тогда:
| Хромосома_1 | 0000000000 | >> | 000 | 1111111 | Результирующая_хромосома_1 |
| Хромосома_2 | 1111111111 | >> | 111 | 0000000 | Результирующая_хромосома_2 |
Затем с вероятностью 0. 5 определяется одна из результирующих хромосом в качестве потомка.
5 определяется одна из результирующих хромосом в качестве потомка.
Следующий генетический оператор предназначен для того, чтобы поддерживать разнообразие особей с популяции. Он называется оператором мутации. При использовании данного оператора каждый бит в хромосоме с определенной вероятностью инвертируется.
Кроме того, используется еще и так называемый оператор инверсии, который заключается в том, что хромосома делится на две части, и затем они меняются местами. Схематически это можно представить следующим образом:
| 000 | 1111111 | >> | 1111111 | 000 |
В принципе для функционирования генетического алгоритма достаточно этих двух генетических операторов, но на практике применяют еще и некоторые дополнительные операторы или модификации данных операторов.
Например, кроссовер может быть не одноточечный (как было описано выше), а многоточечный, когда формируется несколько точек разрыва (чаще всего две). Кроме того, в некоторых реализациях алгоритма оператор мутации представляет собой инверсию только одного случайно выбранного бита хромосомы.
Кроме того, в некоторых реализациях алгоритма оператор мутации представляет собой инверсию только одного случайно выбранного бита хромосомы.
Схема функционирования
Теперь, зная как интерпретировать значения генов, перейдем к описанию функционирования генетического алгоритма. Рассмотрим схему функционирования генетического алгоритма в его классическом варианте:
- Инициировать начальный момент времени t=0. Случайным образом сформировать начальную популяцию, состоящую из k особей B_0={A_1,A_2,…,A_k}
- Вычислить приспособленность каждой особи F_{A_i}=fit(A_i), i=1…k и популяции в целом F_t=fit(B_t) (также иногда называемую термином фитнес). Значение этой функции определяет насколько хорошо подходит особь, описанная данной хромосомой, для решения задачи.
- Выбрать особь A_c из популяции A_c=Get(B_t).
- С определенной вероятностью (вероятностью кроссовера P_c) выбрать вторую особь из популяции A_{c_1}=Get(B_t) и произвести оператор кроссовера A_c=crossing(A_c,A_{c_1}).
- С определенной вероятностью (вероятностью мутации P_m) выполнить оператор мутации A_c=mutation(A_c).
- С определенной вероятностью (вероятностью инверсии P_i) выполнить оператор инверсии A_c=inversion(A_c).
- Поместить полученную хромосому в новую популяцию insert(B_{t+1},A_c).
- Выполнить операции, начиная с пункта 3, k раз.
- Увеличить номер текущей эпохи t=t+1.
- Если выполнилось условие останова, то завершить работу, иначе переход на шаг 2.

Теперь рассмотрим подробнее отдельные этапы алгоритма.
Наибольшую роль в успешном функционировании алгоритма играет этап отбора родительских хромосом на шагах 3 и 4. При этом возможны различные варианты. Наиболее часто используется метод отбора, называемый рулеткой. При использовании такого метода вероятность выбора хромосомы определяется ее приспособленностью, то есть P_{Get{A_i}}=Fit(A_i)/Fit(B_t). Использование этого метода приводит к тому, что вероятность передачи признаков более приспособленными особями потомкам возрастает.
Другой часто используемый метод — турнирный отбор. Он заключается в том, что случайно выбирается несколько особей из популяции (обычно 2) и победителем выбирается особь с наибольшей приспособленностью.
Кроме того, в некоторых реализациях алгоритма применяется так называемая стратегия элитизма, которая заключается в том, что особи с наибольшей приспособленностью гарантировано переходят в новую популяцию. Использование элитизма обычно позволяет ускорить сходимость генетического алгоритма. Недостаток использования стратегии элитизма в том, что повышается вероятность попадания алгоритма в локальный минимум.
Другой важный момент – определение критериев останова. Обычно в качестве них применяются или ограничение на максимальное число эпох функционирования алгоритма, или определение его сходимости, обычно путем сравнивания приспособленности популяции на нескольких эпохах и остановки при стабилизации этого параметра.
Другие материалы по теме:
Loginom Community Edition — аналитика, доступная каждому
Новые возможности Loginom 6. 4. Вебинар
4. Вебинар
Часть генотипа определяющая фенотип называется. Понятие «генотип» и «фенотип»
Генотип и фенотип — это такие понятия, с которыми знакомятся подростки в последних классах общеобразовательной школы. Но не все понимают, что означают эти слова. Мы можем догадываться, что это какая-то классификация характеристик людей. Чем же отличаются эти созвучные названия?
Генотип человека
Генотипом называют все наследственные характеристики человека, то есть совокупность генов, расположенных в хромосомах. Генотип формируется в зависимости от задатков и адаптационных механизмов особи. Ведь каждый живой организм находится в определенных условиях. Животные, птицы, рыбы, простейшие и другие виды живых организмов приспосабливаются к тем условиям, где они обитают. Так и человек, живя в южной части Земного шара, может легко переносить высокую температуру воздуха или слишком низкую посредством цвета кожи. Такие адаптационные механизмы срабатывают не только относительно географического расположения субъекта, но и других условий, одним словом это и называют генотипом.
Что такое фенотип?
Чтобы знать, что такое генотип и фенотип, нужно знать определение этих понятий. С первым понятием уже мы разобрались, а что же означает второе? Фенотип включает в себя все свойства и признаки организма, которые он приобрел в процессе развития. Рождаясь, человек уже имеет свой набор генов, которые определяют его приспосабливаемость к внешним условиям. Но в процессе жизни, под влиянием внутренних и внешних факторов, гены могут мутировать, видоизменяться, поэтому появляется качественно новая структура характеристик человека — фенотип.
История возникновения этих понятий
Что такое генотип и фенотип можно понять, узнав историю возникновения этих научных терминов. В начале ХХ века активно изучалась наука о строении живого организма и биология. Мы помним о теории эволюции и возникновения человека Чарльза Дарвина. Он первым выдвинул Временную гипотезу об отделении клеток в организме (геммулы), из которых впоследствии могла появиться другая особь, так как это половые клетки.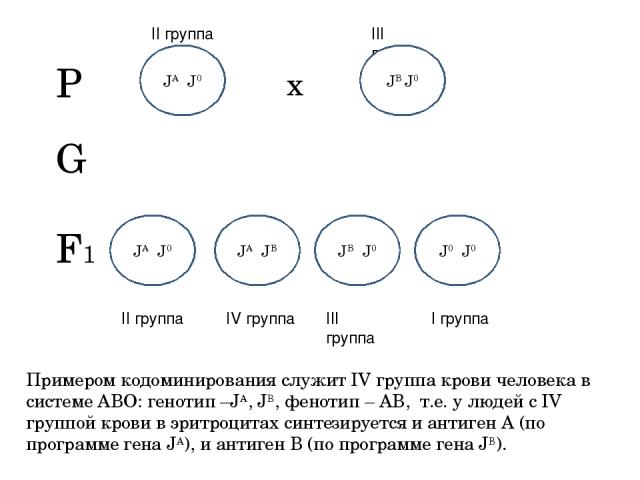 Таким образом, Дарвин развивал теорию о пангенезе.
Таким образом, Дарвин развивал теорию о пангенезе.
Спустя 41 год, в 1909 году ученый ботаник Вильгельм Иогансен на основе уже известного в те годы понятия «генетика» (введенном в 1906 году) ввел в терминологию науки новое понятие — «ген». Ученый заменил им многие слова, которыми пользовались его коллеги, но которые не отражали всю суть врожденных свойств живого организма. Это такие слова, как «детерминанта», «зачатка», «наследственный фактор». В этот же период Иогансен ввел и понятие «фенотип», подчеркивая наследственный фактор в предыдущем научном термине.
Генотип и фенотип человека — в чем разница?
Выделяя два понятия о свойствах и характеристиках живого организма, Иогансен четко определил разницу между ними.
- Гены передаются потомству индивидуумом. Фенотип же особь получает в ходе жизненного развития.
- Генотип и фенотип отличаются еще тем, что гены у живого существа появляются вследствие соединения двух наборов наследственной информации. Фенотип появляется на базе генотипа, претерпевая различные изменения и мутации. Эти изменения происходят под воздействием внешних условий существования живого организма.
- Генотип определяется путем проведения сложного анализа ДНК, фенотип индивидуума можно увидеть при анализе основных критериев внешнего вида.
 Эти изменения происходят под воздействием внешних условий существования живого организма.
Эти изменения происходят под воздействием внешних условий существования живого организма.Нужно отметить, что живые организмы имеют разный уровень приспосабливаемости и чувствительности к окружающим их условиям. От этого зависит, насколько сильно фенотип будет изменен в процессе жизни.
Отличие людей между собой по генотипу и фенотипу
Хоть мы и принадлежим к одному биологическому виду, но между собой сильно отличаемся. Нет двух одинаковых людей, генотип и фенотип каждого будет индивидуальным. Это проявляется, если поместить абсолютно разных людей в одинаково несвойственные для них условия, например, эскимоса отправить в селения Южной Африки, а жителя Зимбабве попросить пожить в условиях тундры. Мы увидим, что этот эксперимент не увенчается успехом, так как эти два человека привыкли обитать в свойственных им географических широтах. Первым отличием людей по гено- и фенотипическим особенностям является адаптация к климато-географическим факторам.
Первым отличием людей по гено- и фенотипическим особенностям является адаптация к климато-географическим факторам.
Следующее отличие продиктовано историко-эволюционным фактором. Оно заключается в том, что в результате миграций населения, войн, культуры определенных народностей, их смешения, сформировались этносы, имеющие свою религию, национальные характеристики и культуру. Поэтому можно увидеть явные различия между стилем и способом жизни, к примеру, славянина и монгола.
Отличия людей также могут быть по социальному параметру. Здесь учитывается уровень культуры людей, образования, социальных притязаний. Недаром существовало такое понятие, как «голубая кровь», свидетельствовавшее о том, что генотип и фенотип дворянина и простолюдина значительно отличались.
Последним критерием различий между людьми является экономический фактор. В зависимости от обеспечения человека, семьи и общества возникают потребности, а, следовательно, и различия между индивидами.
Здравствуйте,
уважаемые читатели блога репетитора биологии по Скайпу .
Вот такая получается «петрушка», если не сказать хуже. Очередной раз сталкиваюсь с тем, что основополагающие понятия генетики в учебниках преподносятся так, что разобраться в них бывает трудно.
Эту статью меня так и подмывало назвать сначала «Фенотип и генотип». Понятно, что фенотип вторичен от генотипа. Но если сам термин «генотип» учащиеся чаще всего могут истолковать правильно, то относительно понятия «фенотипа», как выясняется, нет четкого представления.
Да откуда же ему быть «четкому», если определения фенотипа в учебной литературе носят такой расплывчатый характер.
«Фенотип — совокупность всех внешних признаков организма, определяемых генотипом и условиями окружающей среды». Или «Фенотип — совокупность всех внешних и внутренних признаков и свойств организма, зависящих от генотипа и условий внешней среды».
А если действительно и «внешних», и «внутренних», а это на самом деле так, то в чем тогда отличие фенотипа от генотипа?
Всё же придется начинать не с «хвоста», а с «головы». Уверен, пройдет пара минут и вы, несколько уточнив для себя, что такое «генотип организма», сможете получить более четкое представление и о «фенотипе».
Уверен, пройдет пара минут и вы, несколько уточнив для себя, что такое «генотип организма», сможете получить более четкое представление и о «фенотипе».
Часто термины «признак» и «ген» мы используем как синонимы
Говорят, «генотип — совокупность всех признаков организма». И вот тут то важно понять самое главное — именно к определению генотипа такое определение вносит дополнительную путаницу. Да, действительно, информация о любом признаке закодирована в каком-либо гене (или совокупности генов) организма.
Но всех генов очень много, весь генотип организма огромен, а в течение жизни данной особи или отдельной клетки реализуется (то есть служит образованию каких-либо определенных признаков) лишь незначительная часть генотипа.
Поэтому правильным будет запомнить, что «генотип — совокупность всех генов организма». А уж какие из этих генов реализуются в течение жизни организма в его фенотипе , то есть послужат образованию каких-либо признаков — это зависит как от взаимодействия множества этих генов, так и от конкретных условий окружающей среды.
Таким образом, если правильно понимать, что собой представляет генотип, то не остается и лазейки для путаницы в терминах, обозначающих, что такое «генотип», а что такое «фенотип».
Понятно, что «фенотип — это совокупность всех реализовавшихся в течение жизни организма генов, послуживших образованию конкретных признаков данного организма в определенных условиях среды».
Поэтому на протяжении жизни организма, под действием меняющихся условий среды, фенотип может изменяться, хотя он и базируется на том же самом неизменном генотипе. А в каких границах может меняться фенотип?
Норма реакции
Эти границы для фенотипа четко очерчены генотипом и носят название «нормы реакции». В фенотипе ведь не может проявиться ничего того, чего бы не было уже «записано» ранее в генотипе.
Чтобы лучше понять, что вкладывается в понятие «нормы реакции», разберем на конкретных примерах возможного проявления «широкой» или «узкой» нормы реакции.
Вес (масса) коровы и удойность коровы, какой признак имеет более широкую, а какой более узкую норму реакции?
Понятно, что вес взрослой коровы определенной породы как ее хорошо ни корми не может превысить, к примеру, 900 кг, а при плохом содержании — не может быть меньше 600 кг.
А удойность? При оптимальном содержании и кормлении удойность может меняться от каких-то максимально возможных для данной породы величин, она может упасть до 0, при неблагоприятных условиях содержания. Значит масса коровы имеет довольно узкую норму реакции, а удойность — очень широкую.
Пример с картофелем. Любому очевидно, что «вершки» имеют довольно узкую норму реакции, а масса клубней — очень широкую.
Думаю, теперь всё «устаканилось». Генотип — множество всех генов организма, это весь его потенциал на что он может быть способен в жизни. А фенотип — лишь проявление небольшой части этого потенциала, реализация лишь части генов организма в ряд конкретных признаков в течение его жизни.
Наглядным примером реализации в течение жизни организма части его генотипа в фенотип, являются однояйцевые близнецы. Имея абсолютно одинаковый генотип, в первые годы жизни они почти неотличимы друг от друга фенотипически. Но взрослея, имея сначала незначительные отличия в поведении, в каких-то привязанностях, отдавая предпочтение тому или иному роду деятельности, эти близнецы становятся довольно отличимыми и фенотипически: по выражению лица, строению тела.
В конце этой заметки, я бы хотел вот на что ещё обратить ваше внимание. Слово генотип для изучающих основы генетики имеет как бы два смысла. Выше мы разобрали значение «генотипа» в широком его понимании.
Но для уяснения законов генетики, при решении генетических задач, под словом генотип подразумевают лишь сочетание каких-то конкретных отдельных аллелей одной (моногибридное скрещивание) или двух (дигибридное скрещивание) пар определенных генов, контролирующих проявление конкретного одного или двух признаков.
То есть, и фенотип то у нас при этом какой-то усеченный, говорим «фенотип организма», а сами изучили механизм наследования лишь одного, двух его признаков. В широком же смысле термин «фенотип» относится к любым морфологическим, биохимическим, физиологическим и поведенческим характеристикам организмов.
P.S. В связи с характеристиками понятий «генотип» и «фенотип», уместным было бы здесь разобрать вопрос о наследственной и ненаследственной формах изменчивости организмов. Ну да ладно, об этом как раз и поговорим в .
Ну да ладно, об этом как раз и поговорим в .
***************************************************************
У кого есть вопросы по статье к репетитору биологии по Скайпу , замечания, пожелания — прошу в писать комментарии.
Генотип — вся совокупность имеющихся у организма генов.
Фенотип — совокупность реализованных (т.е. внешних) генетически детерминированных признаков, т.е. индивидуальное (в определенных условиях внешней среды) проявление генотипа.
Генотип – сбалансированная система взаимодействующих генов.
У организмов, размножающихся половым путем, генотип формируется в результате слияния геномов двух родительских половых клеток. Он представляет собой двойной набор генов, заключенных в геноме данного вида. Так как при каждом акте оплодотворения взаимодействующие гаметы несут определенные и часто разные аллели генов, генотип каждого отдельного организма представляет собой оригинальный двойной набор аллелей генов. Таким образом, гены, представленные в геноме уникальными нуклеотидными последовательностями, в генотипе присутствуют в двойной дозе.
Однако многие гены, особенно у эукариот, в результате амплификации присутствуют в геноме в виде нескольких копий (гены гистонов, тРНК, рРНК). Они занимают разное место в геноме, но определяют возможность развития одного и того же признака. Такие нуклеотидные последовательности присутствуют в генотипе в виде многих двойных доз.
Наконец, так как геномы гамет разного пола отличаются друг от друга по набору генов, заключенных в половых хромосомах, в генотипе встречаются гены, представленные лишь одной дозой. Например, у некоторых видов два пола имеют разное число гетерохромосом — XX или ХО. Следовательно, генотипы особей гетерогаметного пола ХО содержат гены Х-хромосомы не в двойной, а в единственной дозе.. Чаще два пола различаются по набору гетерохромосом XX или XY. Ввиду того что морфология этих хромосом различна и одна из них часто крупнее, многие гены имеются лишь в одной гетерохромосоме и отсутствуют или неактивны в другой. В результате в генотипе особей гетерогаметного пола XY гены, расположенные в негомологичных участках Х- и Y-хромосом, встречаются в одной дозе.
У женщин половой хроматин (тельце Барра) имеет вид темного тельца (отмечено стрелкой), располагающегося у оболочки ядра
Таким образом, сформировавшийся в процессе эволюции геном каждого отдельного вида представляет собой совокупность генетических единиц, представленных в нем в строго определенных дозах. В результате и генотипы особей и их клеток — сбалансированные по дозам генов системы.
Значение поддержания определенного дозового соотношения генов в генотипе для формирования видовых характеристик подтверждается возникшим в процессе эволюции механизмом инактивации одной из Х-хромосом у гомогаметного пола XX. Это приводит дозу активно функционирующих Х-генов у данного пола в соответствие с их дозой у гетерогаметного пола ХО или XY.
У млекопитающих гомогаметным является женский пол XX, а гетерогаметным -мужской XY. У мышей такая инактивация происходит на 3-6-е сутки эмбрионального развития. У человека на 16-е сутки во всех клетках женского эмбриона одна из Х-хромосом образует тельце полового хроматина (тельце Барра), которое может быть обнаружено вблизи ядерной мембраны интерфазных клеток в виде хорошо окрашивающегося гетерохроматинового образования (рис. 3.77).
3.77).
Ввиду того что гены, расположенные в инактивированной Х-хромосоме, не функционируют, в генотипе каждой клетки организма гомогаметного пола в диплоидном наборе остальных генов экспрессируется лишь одна доза Х-генов. Так как инактивация Х-хромосомы происходит, когда организм уже представляет собой многоклеточное образо
Аллельные и неаллельные гены.
Аллельные гены – гены, расположенные в одинаковых участках гомологичных хромосом и контролирующие развитие вариаций одного признака.
Неаллельные гены – расположены в разных участках гомологичных хромосом, контролируют развитие разных признаков.
Генотип — это совокупность всех генов организма, являющихся его наследственной основой.
Фенотип — совокупность всех признаков и свойств организма, которые выявляются в процессе индивидуального развития в данных условиях и являются результатом взаимодействия генотипа с комплексом факторов внутренней и внешней среды.
Фенотип в общем случае — это то, что можно увидеть (окрас кошки), услышать,
ощутить (запах), а также поведение животного. Договоримся, что мы будем
рассматривать фенотип только с точки зрения окраса.
Договоримся, что мы будем
рассматривать фенотип только с точки зрения окраса.
Что касается генотипа, то о нем чаще всего говорят, имея в виду некую небольшую группу генов. Пока же давайте считать, что наш генотип состоит всего лишь из одного гена W (в следующих параграфах мы будем последовательно добавлять к нему другие гены).
У гомозиготного животного генотип совпадает с фенотипом, а у гетерозиготного — нет.
Действительно, в случае генотипа WW , обе аллели отвечают за белый окрас, и кошка будет белой. Аналогично ww — обе аллели отвечают за не белый окрас, и кошка будет не белой.
А вот в случае генотипа Ww кошка внешне (фенотипически) будет белой, однако в своем генотипе она будет нести рецессивную аллель не белого окраса w .
Каждый биологический вид имеет свойственный только ему фенотип. Он формируется в
соответствии с наследственной информацией, заложенной в генах. Однако в
зависимости от изменений внешней среды состояние признаков варьирует от
организма к организму, в результате чего возникают индивидуальные различия —
изменчивость.
На основе изменчивости организмов появляется генетическое разнообразие форм. Различают изменчивость модификационную, или фенотипическую, и генетическую, или мутационную.
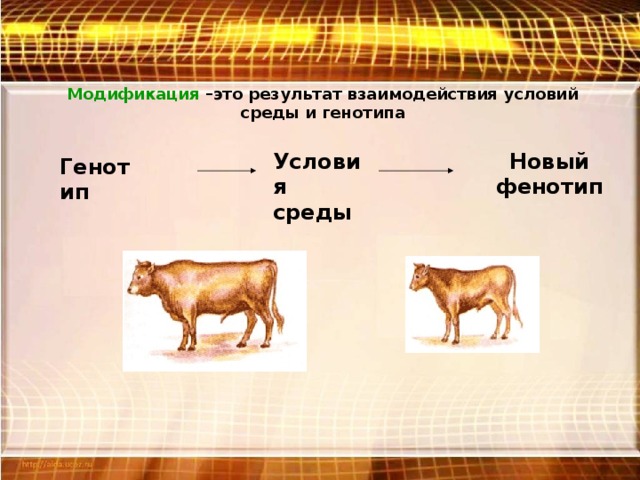
Модификационная изменчивость не вызывает изменений генотипа, она связана с реакцией данного, одного и того же генотипа на изменение внешней среды: в оптимальных условиях выявляется максимум возможностей, присущих данному генотипу. Модификационная изменчивость проявляется в количественных и качественных отклонениях от исходной нормы, которые не передаются по наследству, а носят лишь приспособительный характер, например, усиление пигментации кожи человека под действием ультрафиолетовых лучей или развития мышечной системы под действием физических упражнений и т.д.
Степень варьирования признака у организма, то есть пределы модификационной
изменчивости называются нормой реакции. Таким образом, фенотип формируется в
результате взаимодействия генотипа и факторов среды, Фенотипические признаки не
передаются от родителей к потомкам, наследуется лишь норма реакции, то есть
характер реагирования на изменение окружающих условий.
Генетическая изменчивость бывает комбинативной и мутационной.
Комбинативная изменчивость возникает в результате обмена гомологичными участками гомологичных хромосом в процессе мейоза, что приводит к образованию новых объединений генов в генотипе. Возникает в результате трех процессов:
1) независимого расхождения хромосом в
процессе мейоза;
2) случайного соединения их при
оплодотворении;
3) обмена участками гомологичных хромосом
или конъюгации.
Мутационная изменчивость. Мутациями называют скачкообразные и устойчивые
изменения единиц наследственности — генов, влекущие за собой изменения
наследственных признаков. Они обязательно вызывают изменения генотипа, которые
наследуются потомством и не связаны со скрещиванием и рекомбинацией генов.
Существуют хромосомные и генные мутации. Хромосомные мутации связаны с
изменением структуры хромосом. Это может быть изменение числа хромосом кратное
или не кратное гаплоидному набору (у растений — полиплоидия, у человека —
гетероплоидия). Примером гетероплоидии у человека может быть синдром Дауна (одна
лишняя хромосома и в кариотипе 47 хромосом), синдром Шерешевского — Тернера
(отсутствует одна Х-хромосома, 45). Такие отклонения в кариотипе человека
сопровождаются расстройством здоровья, нарушение психики и телосложения,
снижением жизнеспособности и др.
Примером гетероплоидии у человека может быть синдром Дауна (одна
лишняя хромосома и в кариотипе 47 хромосом), синдром Шерешевского — Тернера
(отсутствует одна Х-хромосома, 45). Такие отклонения в кариотипе человека
сопровождаются расстройством здоровья, нарушение психики и телосложения,
снижением жизнеспособности и др.
Генные мутации — затрагивают структуру самого гена и влекут за собой изменение
свойств организма (гемофилия, дальтонизм, альбинизм и др.). Генные мутации
возникают как в соматических, так и в половых клетках.
Мутации, возникающие в половых клетках, передаются по наследству. Их называют
генеративными мутациями. Изменения в соматических клетках вызывают соматические
мутации, распространяющиеся на ту часть тела, которая развивается из
изменившейся клетки. Для видов, размножающихся половым путем, они не имеют
существенного значения, для вегетативного размножения растений они важны.
Слово «фенотип» имеет греческое происхождение и переводится (дословно) «обнаруживаю», «являю». Каково же практическое значение данного понятия?
Каково же практическое значение данного понятия?
Что такое фенотип? Определение
Под фенотипом следует понимать комплекс характеристик, которые присущи индивиду на конкретном этапе развития. Формируется эта совокупность на основе генотипа. Для диплоидных организмов характерно проявление Точнее определяя, что такое фенотип, следует, говорить о совокупности внутренних и внешних признаков организма, которые были приобретены в процессе
Общие сведения
Несмотря на достаточно точное фенотип, его концепция имеет ряд неопределенностей. Большинство структур и молекул, которые кодируются генетическим материалом, не обнаруживаются во внешнем виде организма. При этом они являются частью фенотипа. Примером может являться фенотип крови людей. В связи с этим, по мнению ряда авторов, определение должно включать в себя и те характеристики, которые могут быть получены с применением диагностических, медицинских или технических процедур. Более радикальное дальнейшее расширение может содержать приобретенное поведение, а при необходимости и влияние организма на среду обитания и остальные организмы. Так, к примеру, резцы и можно принимать за их фенотип.
Так, к примеру, резцы и можно принимать за их фенотип.
Основные характеристики
Определяя, что такое фенотип, можно говорить о некотором «выносе» генетических сведений навстречу факторам окружающей среды. При первом приближении следует рассматривать две характеристики:
- Мерность фенотипа. Этот признак указывает на количество направлений «выноса», которое характеризует количество факторов окружающей среды.
- Второй признак указывает на уровень чувствительности фенотипа к окружающим условиям. Называется эта степень — дальностью.
В комплексе эти характеристики свидетельствуют о богатстве и разновидности фенотипа. Чем многомернее совокупность индивидуальных особенностей, чем чувствительнее признаки и чем дальше они от генотипа, тем он богаче. Так, например, если сравнивать фенотип бактерии, аскариды, лягушки, человека, то «богатство» в этой цепочке повышается. Это значит, что фенотип человека богаче.
Историческая справка
В 1909 году Вильгельм Иогансен (датский ученый) впервые — в комплексе с концепцией генотипа — предложил определение фенотипа. Это позволило отличать наследственность от результата ее реализации. Идею о различиях можно проследить и в работах Менделя и Вейсмана. При этом последний отличал соматические и репродуктивные клетки в Полученный от родителей хромосомный набор содержится в Хромосомы несут комплекс генов, характерных для конкретного вида в целом и определенного организма в частности. В генах содержится информация о белках, способных синтезироваться, а также о механизмах, которые, собственно, определяют и регулируют синтез. Что же при этом происходит? При онтогенезе последовательно включаются гены и синтезируются те белки, которые ими закодированы. В итоге происходит формирование и развитие всех свойств и признаков организма, составляющих его фенотип. Другими словами, получается некий «продукт» от реализации генетической программы, содержащейся в генотипе.
Это позволило отличать наследственность от результата ее реализации. Идею о различиях можно проследить и в работах Менделя и Вейсмана. При этом последний отличал соматические и репродуктивные клетки в Полученный от родителей хромосомный набор содержится в Хромосомы несут комплекс генов, характерных для конкретного вида в целом и определенного организма в частности. В генах содержится информация о белках, способных синтезироваться, а также о механизмах, которые, собственно, определяют и регулируют синтез. Что же при этом происходит? При онтогенезе последовательно включаются гены и синтезируются те белки, которые ими закодированы. В итоге происходит формирование и развитие всех свойств и признаков организма, составляющих его фенотип. Другими словами, получается некий «продукт» от реализации генетической программы, содержащейся в генотипе.
Влияние внешних условий на развитие индивидуальных признаков
Следует отметить, что генотип не является однозначным фактором, определяющим фенотип. В той или иной степени формирование совокупности индивидуальных признаков будет зависеть и от среды пребывания, то есть от внешних факторов. В разных условиях фенотипы имеют резкое отличие. Так, например, вид бабочек «арашния» в год дает два потомства. Те особи, которые вышли из перезимовавших куколок (весенние), резко отличаются от тех, которые появились в летнее время. Так же может отличаться и фенотип растения. Например, на открытом пространстве сосны развесистые, а в лесу — стройные и высокие. У водяного лютика форма листа зависит от того, где он пребывает — на воздухе или же в воде.
В той или иной степени формирование совокупности индивидуальных признаков будет зависеть и от среды пребывания, то есть от внешних факторов. В разных условиях фенотипы имеют резкое отличие. Так, например, вид бабочек «арашния» в год дает два потомства. Те особи, которые вышли из перезимовавших куколок (весенние), резко отличаются от тех, которые появились в летнее время. Так же может отличаться и фенотип растения. Например, на открытом пространстве сосны развесистые, а в лесу — стройные и высокие. У водяного лютика форма листа зависит от того, где он пребывает — на воздухе или же в воде.
Взаимоотношения фенотипов и генотипов
Способность к изменениям, которая предусмотрена генетической программой, называется нормой реакции. Как правило, чем разнообразнее условия, в которых обитает вид, тем шире эта норма. В том случае, когда окружающая среда резко отличается от той, к которой приспособлен вид, происходит нарушение в развитии организмов, и они погибают. В признаках фенотипа не всегда отражаются рецессивные аллели.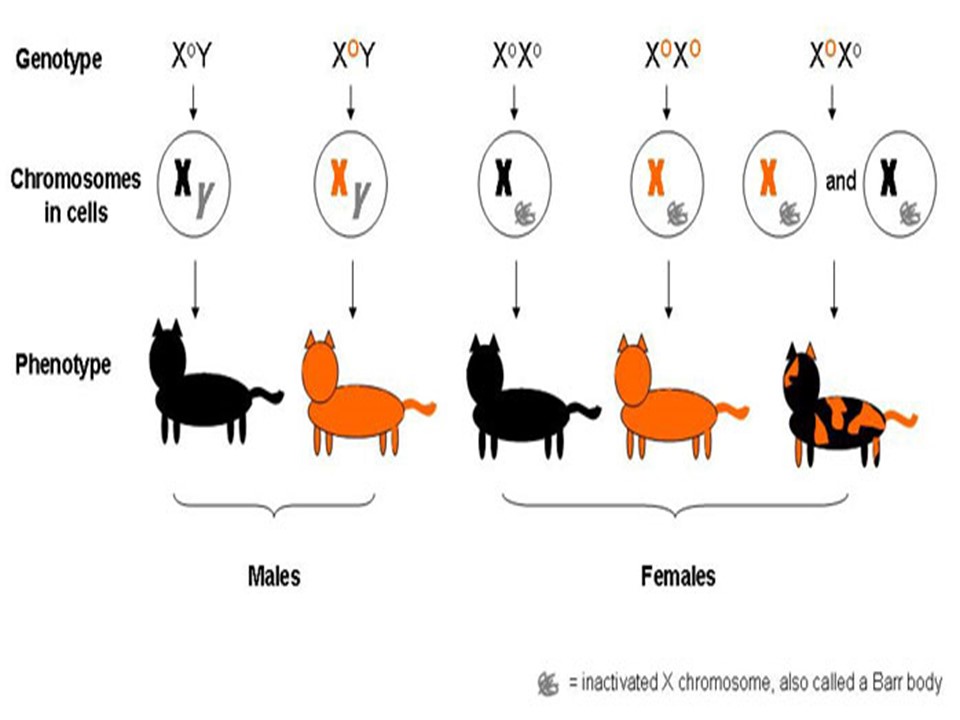 Но при этом они сохраняются и могут перейти к потомству. Эти сведения позволяют лучше понять эволюционный процесс. В участвуют только фенотипы, потомству же передаются и остаются далее в популяции генотипы. Взаимодействие не исчерпывается взаимоотношениями рецессивных и доминантных аллелей — взаимодействуют многие гены друг с другом.
Но при этом они сохраняются и могут перейти к потомству. Эти сведения позволяют лучше понять эволюционный процесс. В участвуют только фенотипы, потомству же передаются и остаются далее в популяции генотипы. Взаимодействие не исчерпывается взаимоотношениями рецессивных и доминантных аллелей — взаимодействуют многие гены друг с другом.
Основы генетики
Автор статьи Бусыгин С.А.
ВведениеГенетика как наука по своей сути тесно связана с эволюционным учением с одной стороны, и с цитологией и молекулярной биологией — с другой. Как следствие, генетика необходима для правильного понимания биологических процессов на всех уровнях организации живой материи. В этом кратком пособии представлены наиболее часто встречающиеся задачи олимпиадного уровня по генетике и необходимые для их решения указания, сформулированы общеупотребимые понятия и продемонстрирован надлежащий путь восприятия биологической реальности с позиций современной науки.
Данный текст предполагает наличие у читателей первичных знаний по генетике, а именно знакомства с основными понятиями генетики, используемой при решении генетических задач символикой и законами Менделя.
В современной генетике для исследования закономерностей наследования применяют следующие методы:
- Гибридологический – создание систем скрещиваний, которые позволяют проследить закономерности наследования признаков путем строгого подбора родителей, различающихся по контрастным признакам, строгого количественного учета распределения признаков у гибридов и индивидуальной оценки потомства в ряду поколений
- Генеалогический – составление и анализ родословных
- Близнецовый – изучение близнецов позволяет определить, насколько степень проявления признака зависит от генотипа и от внешней среды
- Молекулярно-биологический – изучение первичной структуры молекул наследования на уровне от генов и до хромосом
- Популяционно-генетический – изучение генетической структуры популяций и связанных с ней эволюционных закономерностей.
Дезоксирибонуклеиновой кислоте отведена центральная роль в процессе хранения информации у всех клеточных организмов. Именно поэтому методы молекулярной биологии, работающих конкретно с этим полимером, так разнообразны: секвенирование последовательности нуклеотидов, полимеразная цепная реакция, гель-электрофорез, рестрикционный анализ и саузерн блоттинг.
Именно поэтому методы молекулярной биологии, работающих конкретно с этим полимером, так разнообразны: секвенирование последовательности нуклеотидов, полимеразная цепная реакция, гель-электрофорез, рестрикционный анализ и саузерн блоттинг.
- Секвенирование – процесс определения последовательности азотистых оснований в цепочке ДНК. В настоящее время используется для получения полных геномов организмов и задач сходного масштаба.
- Полимеразная цепная реакция – процесс получения многочисленных копий участка ДНК (около тысячи пар оснований), заключенного между специально подобранными короткими последовательностями (праймерами). Обычно это подготовительный этап для других методов.
- Гель-электрофорез – способ разделения линейных фрагментов ДНК в агарозном геле в зависимости от их длины (более короткие мигрируют быстрее) за счет энергии электрического поля и последующей визуализации при помощи флуоресцентных красителей, специфически взаимодействующих с ДНК. Часто параллельно с исследуемыми образцами форез проходит набор линейных фрагментов ДНК известной длины, что позволяет точнее оценить длину исходных фрагментов.
- Рестрикционный анализ – изучение ДНК с применением эндонуклеаз рестрикции, то есть ферментов, которые вносят в ДНК двуцепочечный разрыв, но только в тех местах, где встречается специфическая для каждой конкретной рестриктазы последовательность нуклеотидов.
- Саузерн-блоттинг – способ поиска в исследуемой ДНК определенных последовательностей, комплементарных известной последовательности ДНК-метки. Этому этапу предшествует гель-электрофорез. Метод помогает в поиске гомологичных генов и копий гена в геноме.
С помощью этих методов можно обосновать различия генотипов организмов на молекулярном уровне и визуализировать различные мутации. Комбинируя гель-электрофорез с использованием рестриктаз, можно картировать ДНК. Для этого сопоставляют результаты гидролиза исследуемого фрагмента каждой рестриктазой в отдельности и их комбинациями. Затем эти фрагменты разделяют с помощью электрического поля и оценивают их размеры. Результаты совместного гидролиза показывают, содержится ли сайты узнавания разных рестриктаз внутри фрагмента, вычленяемого определенной рестриктазой. Если они присутствуют, то такой фрагмент исчезает в геле и превращается в два и более субфрагмента с такой же суммарной длиной. Сопоставив размеры фрагментов, можно определить локализацию сайтов рестрикции.
Если они присутствуют, то такой фрагмент исчезает в геле и превращается в два и более субфрагмента с такой же суммарной длиной. Сопоставив размеры фрагментов, можно определить локализацию сайтов рестрикции.
Пример олимпиадной задачи на построение рестрикционных карт (заключительный этап олимпиады Ломоносов по биологии 2014/15 учебного года)
У многих бактерий для защиты от вирусов есть специальные ферменты – рестриктазы. Они расщепляют ДНК по определённым последовательностям, которые в ДНК бактерий данного вида отсутствуют или модифицированы присоединением к основанию метильной группы. Эти ферменты называют по первым буквам латинского названия рода и вида бактерии, например, Есо – Еsсherichia coli – рестриктаза из кишечной палочки. При действии такого фермента на очищенную ДНК разрывы происходят в строго определённых местах, и образуются фрагменты ДНК определённой длины.
Сравнивая расщепление исследуемой ДНК различными рестриктазами и их комбинациями, можно определить относительное расположение точек расщепления и построить рестрикционную карту данной последовательности ДНК. Из клеток бактерий выделили небольшую кольцевую ДНК – плазмиду, несущую ген устойчивости к пенициллину. Расщепление этой плазмиды тремя рестриктазами дало следующие фрагменты (см. таблицу). По этим данным постройте рестрикционнную карту плазмиды, расположив на ней все точки расщепления. Ответ обоснуйте и оформите по образцу (как на рис.).
Из клеток бактерий выделили небольшую кольцевую ДНК – плазмиду, несущую ген устойчивости к пенициллину. Расщепление этой плазмиды тремя рестриктазами дало следующие фрагменты (см. таблицу). По этим данным постройте рестрикционнную карту плазмиды, расположив на ней все точки расщепления. Ответ обоснуйте и оформите по образцу (как на рис.).
Решение
Один из подходов к решению этой задачи – рассмотреть положение сайтов рестрикции Sal и Hind относительно друг друга. Sal даёт два фрагмента одинаковой длины (5 тыс. п.н.), после обработки Sal+Hind получаются фрагменты 4; 3; 2 и 1. Далее с помощью элементарной арифметики можно определить, что один из фрагментов Sal разрезается Hind на фрагменты 4 и 1 (4 + 1 = 5), а другой – на фрагменты 3 и 2 (3 + 2= 5).
Рестриктаза Hind даёт фрагменты 6 и 4 тыс. п. н. После обработки Sal+Hind фрагмент 6 тыс. п.н. разрезается на фрагменты 4 и 2 тыс. п. н., а фрагмент 4 тыс. п.н. – на 3 и 1 соответственно. Этих данных достаточно для начала построения рестрикционной карты плазмиды. Для удобства обозначим разными цветами каждую из пар, состоящих из сайта рестрикции Sal и близлежащего сайта Hind.
Для удобства обозначим разными цветами каждую из пар, состоящих из сайта рестрикции Sal и близлежащего сайта Hind.
Теперь можно установить взаимное расположение сайтов Sal и Ava. При совместном действии Sal+Ava получается четыре фрагмента: 3,5 тыс. п. н.; 3 тыс. п. н., 2 тыс. п. н. и 1,5 тыс. п. н. Очевидно, что один из фрагментов Sal разрезается Ava на 3,5 и 1,5 тыс. п. н. (3,5 + 1,5 = 5), а второй – на 3 и 2 тыс. п. н. (3 + 2 = 5) соответственно. Аналогично можно построить карту для Sal и Ava.
Теперь построим еще одну вспомогательную карту для Hind и Ava. Очевидно, что фрагмент Hind длиной 6 тыс. п. н. разрезается Ava на фрагменты 4 и 2 тыс. п. н., а фрагмент Hind 4 тыс. п. н. разрезается Ava на фрагменты 2,5 и 1,5 тыс. п. н. При этом фрагменты 4 и 1,5 должны оказаться рядом (как и фрагменты 2 и 2,5). Это следует из данных по обработке плазмиды только Ava. Карта по Hind и Ava выглядит следующим образом.
Для окончательного решения необходимо совместить все полученные рестрикционные карты. При этом мы можем столкнуться с ситуацией, когда одну из карт придётся либо симметрично отобразить, либо повернуть на некоторый угол. Есть две возможности такого совмещения.
При этом мы можем столкнуться с ситуацией, когда одну из карт придётся либо симметрично отобразить, либо повернуть на некоторый угол. Есть две возможности такого совмещения.
- Сайт Hind (1) на рис. В соответствует сайту Hind*, помеченному зелёным цветом на рис. А.
- Сайт Hind (1) на рис. В соответствует сайту Hind, помеченному розовым цветом на рис. А.
Рассмотрим обе возможности.
- В случае Hind(1)=Hind* сайты рестрикции Sal должны находиться на расстоянии 2 (Sal*) и 3 тыс. п. н. (Sal). Тогда точка рестрикции Sal окажется на расстоянии 0,5 тыс. п. н. от Ava (1) (см. рис. В). При совместном действии на ДНК плазмиды Sal+Ava должен получиться фрагмент длиной 0,5 тыс. п. н. Однако это противоречит условию: получаются фрагменты короче 1,5 тыс. п. н.
- В случае Hind(1)=Hind сайты рестрикции Sal должны находиться на расстоянии 1 (Sal) и 4 тыс. п. н. (Sal*). В этом случае сайт Sal окажется на расстоянии 1,5 от сайта Ava(1), а сайт Sal* – на расстоянии 2,0 от сайта Ava(2). Таким образом, сопоставляя рис. Б и рис. В, мы находим, что Ava(1)=Ava*, тогда как Ava(2)=Ava, и мы должны отобразить карту на рис. Б зеркально. Итоговая рестрикционная карта выглядит следующим образом.
 п. н.
п. н.Ответ: рис.
Если карта зеркально симметрична приведённой в ответе, и/или повёрнута на некоторый угол, это не является ошибкой. Задачу также можно было начинать решать с построения начальной карты для любой другой рестриктазы.
Структура гена и его действие через продукт
Подавляющее большинство генов проявляют свою функцию через кодируемый ими белок (структурные гены). В ходе процесса трансляции согласно последовательности оснований нуклеиновых кислот в соответствии с генетическим кодом образуется последовательность аминокислот. Генетический код обладает следующими свойствами:
В ходе процесса трансляции согласно последовательности оснований нуклеиновых кислот в соответствии с генетическим кодом образуется последовательность аминокислот. Генетический код обладает следующими свойствами:
- Триплетность – каждая аминокислота кодируется последовательностью из трех нуклеотидов
- Однозначность – каждый триплет кодирует одну аминокислоту либо сигнал конца трансляции
- Вырожденность – большинству аминокислот соответствует несколько разных триплетов
- Знаки препинания отсутствуют внутри гена и обязательно встречаются в конце его последовательности
- Универсальность – одинаковые триплеты кодируют одинаковые аминокислоты у различных форм жизни
Проще всего отслеживать те белки, которые катализируют какую-то химическую реакцию. Например, изучая мутантные клетки, не способные к синтезу какого-либо вещества, можно установить последовательность химических реакций в метаболическом пути. В олимпиадной практике это может выглядеть как задача про тест на синтрофизм. Рассмотрим такую задачу, предложенную на теоретическом туре B международной олимпиады по биологии 2014 года.
Рассмотрим такую задачу, предложенную на теоретическом туре B международной олимпиады по биологии 2014 года.
Три мутантных штамма бактерий TrpB-, TrpE- и TrpD-, дефективные по одной из ступеней пути биосинтеза триптофана, были нанесены штрихами на чашку Петри (см. рисунок ниже). В среде была ограниченная концентрация триптофана, позволившая вырасти тонким штрихам, прежде чем триптофан в среде кончился. Но некоторые фрагменты штрихов продолжили свой рост и стали толще. В ходе синтеза триптофана происходит превращение хоризмата в антранилат, индол, и, в конце концов, триптофан.
Установите истинность следующих утверждений:
- Из результатов следует, что промежуточные соединения активно секретируются клетками культуры
- У TrpD- мутация в ферменте, катализирующем превращения индола в триптофан
- TrpE- способен синтезировать триптофан, если в среде есть антранилат или индол
- Индол будет накапливаться в среде, в которой TrpB- клетки расположены в непосредственной близости от TrpD-
Решение: Тест на синтрофизм основан на том, что мутация с утратой функции влечет за собой прекращение дальнейшего использования промежуточного метаболита, занимающего место в цепи биосинтеза непосредственно перед блокированной стадией. В результате промежуточный метаболит накапливается в мутантной клетке и может выделяться в ростовую среду. Такие мутантные клетки способны поддерживать с помощью выделяемого ими метаболита рост других мутантных клеток, у которых блокированы более ранние этапы данной цепи биосинтеза. Лимитированное количество конечного продукта пути биосинтеза, в данном случае триптофана, необходимо для поддержания слабого роста штрихов мутантных клеток, позволяющего им выделять в среду диффундирующие метаболиты. Обильный рост на концах штрихов клеток TrpD и TrpE объясняется тем, что они получают необходимый для их роста метаболит от клеток TrpB, а клетки TrpE еще и от TrpD. Поскольку клетки TrpB поддерживают рост TrpD и TrpE, ген TrpB занимает место в процессе биосинтеза триптофана после TrpD и TrpE. Клетки TrpD поддерживают рост TrpE, следовательно, ген TrpD расположен после гена TrpE. Порядок генов в контроле процесса биосинтеза триптофана:
В результате промежуточный метаболит накапливается в мутантной клетке и может выделяться в ростовую среду. Такие мутантные клетки способны поддерживать с помощью выделяемого ими метаболита рост других мутантных клеток, у которых блокированы более ранние этапы данной цепи биосинтеза. Лимитированное количество конечного продукта пути биосинтеза, в данном случае триптофана, необходимо для поддержания слабого роста штрихов мутантных клеток, позволяющего им выделять в среду диффундирующие метаболиты. Обильный рост на концах штрихов клеток TrpD и TrpE объясняется тем, что они получают необходимый для их роста метаболит от клеток TrpB, а клетки TrpE еще и от TrpD. Поскольку клетки TrpB поддерживают рост TrpD и TrpE, ген TrpB занимает место в процессе биосинтеза триптофана после TrpD и TrpE. Клетки TrpD поддерживают рост TrpE, следовательно, ген TrpD расположен после гена TrpE. Порядок генов в контроле процесса биосинтеза триптофана:
Теперь рассмотрим истинность утверждений:
- Из результатов следует, что промежуточные соединения активно секретируются из клеток. Неверно, так как эксперимент не был связан с активным транспортом.
- У TrpD- мутация в ферменте, катализирующем превращения индола в триптофан. Неверно, так как у TrpD мутация в ферменте, превращающем антранилат в индол.
- TrpE- способен синтезировать триптофан, если в среде есть антранилат или индол. Верно, так как у него нарушена более ранняя стадия биосинтетического пути.
- Индол будет накапливаться в среде, в которой TrpB- клетки расположены в непосредственной близости от TrpD-. Неверно. Хотя TrpB- клетки выделяют в окружающую среду именно индол, он не будет накапливаться, так как TrpD способен синтезировать триптофан из индола и будет его потреблять из среды.
 Неверно, так как эксперимент не был связан с активным транспортом.
Неверно, так как эксперимент не был связан с активным транспортом.В этом разделе рассматриваются случаи, когда признак кодируется одним геном, гены не входят в группы сцепления, а их продукты никак не взаимодействуют друг с другом. Но тем не менее, кажущаяся простота таких задач может негативно отразиться на способности их решать, если не помнить о следующих сложностях:
- Полное/неполное доминирование. В случае если фенотип гетерозиготы совпадает с фенотипом доминантной гомозигиты, аллельные гены взаимодействуют по принципу полного доминирования. В случае неполного доминирования фенотип гетерозиготы не совпадает с фенотипами гомозигот и представляет собой среднее (промежуточное) между доминантным и рецессивным фенотипом.
- Пенетратность – отношение частоты проявления фенотипа к частоте генотипа
- Плейотропизм – множественный эффект генов за счет многообразия ролей продукта. Пример: синдром Морфана. Мутация в гене фибриллина-1 приводит к развитию паучьих пальцев, сдвигу хрусталика и пролапсу сердечных клапанов у носителя.
- Множественный аллелизм – наличие двух и более вариантов аллеля у гена
- Кодоминирование – формирование у гетерозиготы фенотипа, отличного от фенотипа гомозигот, в результате присутствия продуктов обоих аллелей. Например, четвертая группа крови у людей возникает из-за присутствия на поверхности эритроцитов обоих вариантов антигенов
- Сверхдоминирование – гетерозиготы обычно более сильные и лучше приспособлены по сравнению с обеими гомозиготами. В качестве примера можно привести серповидноклеточную анемию у человека в регионах с сильной угрозой малярии. Рецессивные гомозиготы по этому признаку погибают в детстве, а в эритроцитах гетерозигот плазмодию размножаться труднее, чем в нормальных. Доминантные гомозиготы имеют обычные эритроциты.
 В случае если фенотип гетерозиготы совпадает с фенотипом доминантной гомозигиты, аллельные гены взаимодействуют по принципу полного доминирования. В случае неполного доминирования фенотип гетерозиготы не совпадает с фенотипами гомозигот и представляет собой среднее (промежуточное) между доминантным и рецессивным фенотипом.
В случае если фенотип гетерозиготы совпадает с фенотипом доминантной гомозигиты, аллельные гены взаимодействуют по принципу полного доминирования. В случае неполного доминирования фенотип гетерозиготы не совпадает с фенотипами гомозигот и представляет собой среднее (промежуточное) между доминантным и рецессивным фенотипом. В качестве примера можно привести серповидноклеточную анемию у человека в регионах с сильной угрозой малярии. Рецессивные гомозиготы по этому признаку погибают в детстве, а в эритроцитах гетерозигот плазмодию размножаться труднее, чем в нормальных. Доминантные гомозиготы имеют обычные эритроциты.
В качестве примера можно привести серповидноклеточную анемию у человека в регионах с сильной угрозой малярии. Рецессивные гомозиготы по этому признаку погибают в детстве, а в эритроцитах гетерозигот плазмодию размножаться труднее, чем в нормальных. Доминантные гомозиготы имеют обычные эритроциты.Еще нужно обращать внимание на то, заданы ли генотипы в задаче прямо или косвенно. Проиллюстрируем это на примере задачи из регионального этапа ВОШ по биологии 16/17 учебного года
У собак часто встречается рецессивная глухота, не сцепленная с полом. Признак определяется одним геном. Вы завели пару собак с нормальным слухом, при этом, несмотря на то что все их родители имели нормальный слух, и у самца, и у самки были сибсы (братья и сёстры) с глухотой. Какова вероятность рождения глухого щенка у этой пары?
Чтобы найти вероятность рождения глухого щенка, для начала определим генотипы родителей и старшего поколения. Так как у родителей есть глухие сибсы (рецессивные гомозиготы), а всё старшее поколение здорово, оба родителя этой пары собак гетерозиготны. Так как исходная пара здорова, то при таком фенотипе для каждого из них существует вероятность того, что они гетерозиготны, которая равна 2/3, и 1/3 – что они доминантные гомозиготы. Щенок будет глухим, только если он родиться от гетерозиготных родителей (вероятность того, что пара гетерозиготна, равна 4/9) и получит от каждого рецессивный ген (вероятность 1/4). Следовательно, итоговая вероятность искомого события 1/9.
Так как исходная пара здорова, то при таком фенотипе для каждого из них существует вероятность того, что они гетерозиготны, которая равна 2/3, и 1/3 – что они доминантные гомозиготы. Щенок будет глухим, только если он родиться от гетерозиготных родителей (вероятность того, что пара гетерозиготна, равна 4/9) и получит от каждого рецессивный ген (вероятность 1/4). Следовательно, итоговая вероятность искомого события 1/9.
Задача из заключительного этапа ВОШ по биологии 14/15
В попытках вывести чистую линию коротконогих кур селекционер десять поколений скрещивал коротконогих кур друг с другом, но при этом всегда около трети цыплят имели обычные ноги и примерно две трети — короткие ноги. Это связано с тем, что:
- ген коротконогости находится на Х-хромосоме и проявляется у самцов; Неверно, так как про пол ничего не сказано. Отношение полов у кур 1:1
- ген коротконогости летален в гомозиготе и проявляется у гетерозигот; Верно, пример плейотропного действия гена.
- ген коротконогости по-разному проявляет себя у самцов и самок; Неверно, соотношение полов у кур 1:1, про пол потомков ничего не сказано.
- коротконогость связана с несколькими независимо наследуемыми генами. Неверно, расщепление при нескольких независимых генах не такое простое.
Множество биологических процессов состоит из нескольких стадий. Кроме того, в геноме могут присутствовать регуляторные элементы, влияющие на возможность проявления признака. Традиционно к неаллельным взаимодействиям относят следующие явления:
- Комплементарность – признак формируется при сочетании продуктов доминантных аллелей. Пример задания из заключительного этапа ВОШ по биологии 14/15.
В школьной лаборатории учащиеся старших классов проводили изучение синтеза пигмента в лепестках некоторого вида растений. Для эксперимента использовали два сорта, обладающие цветками с белыми венчиками. Сначала ребята измельчали лепестки, после чего изготавливали экстракты. При смешивании полученных растворов первоначально бесцветная жидкость со временем приобретала пурпурный оттенок. Ознакомьтесь со схемой эксперимента. Известно, что за признак отвечают два гена. Какое расщепление следует ожидать в F2 в случае скрещивания растений сорта №1 и сорта №2 друг с другом? Считайте, что оба сорта являются чистыми линиями (т.е. гомозиготны по всем генам).
Сначала ребята измельчали лепестки, после чего изготавливали экстракты. При смешивании полученных растворов первоначально бесцветная жидкость со временем приобретала пурпурный оттенок. Ознакомьтесь со схемой эксперимента. Известно, что за признак отвечают два гена. Какое расщепление следует ожидать в F2 в случае скрещивания растений сорта №1 и сорта №2 друг с другом? Считайте, что оба сорта являются чистыми линиями (т.е. гомозиготны по всем генам).
Решение: Так как линии гомозиготные, разумно предположить, что у них нарушены разные ступени биохимического процесса, и один из сортов накапливает бесцветный предшественник пигмента, который обрабатывается ферментом, содержащимся во втором сорте. В F1 присутствует дигетерозигота пурпурного цвета, а в F2 будут окрашенными только особи, содержащие хотя бы по одному доминантному гену каждого аллеля. Заполним решетку Пеннета для этого скрещивания. Посчитав соотношение фенотипов, получим ответ 9:7
- Эпистаз – вид взаимодействия, при котором одна пара подавляет другую. Если ген-подавитель доминантный, то эпистаз тоже доминантный, в противном случае наблюдается рецессивный эпистаз. Пример задачи из заключительного этапа ВОШ по биологии 14/15.
 Если ген-подавитель доминантный, то эпистаз тоже доминантный, в противном случае наблюдается рецессивный эпистаз. Пример задачи из заключительного этапа ВОШ по биологии 14/15.
Если ген-подавитель доминантный, то эпистаз тоже доминантный, в противном случае наблюдается рецессивный эпистаз. Пример задачи из заключительного этапа ВОШ по биологии 14/15.На рисунке представлена схема взаимодействия молекулярных продуктов генов I и II, объясняющая наследование окраски плодов некоторого растения. Известно, что ген I может быть представлен двумя аллелями – G и g. Аллель G отвечает за синтез мРНК, несущей информацию о строении функционально исправного белка. Аллель g кодирует мРНК, на которой синтезируется неактивный фермент. Известно, что ген II бывает представлен также двумя аллелями W и w. Аллель W кодирует регуляторную РНК, способную связываться с мРНК гена I. Продукт аллеля w не способен связываться с мРНК гена I. В клетке концентрация РНК гена II сильно выше концентрации мРНК гена I. Какое расщепление следует ожидать в потомстве от скрещивания дигетерозигот?
Решение: Так как концентрация РНК гена II превосходит концентрацию мРНК гена II, то даже в случае одновременного синтеза продуктов аллелей W и w связыванию помех не будет и мРНК деградирует. Это означает, что здесь наблюдается рецессивный эпистаз. Составим решетку Пеннета. По ней мы можем расщепление при скрещивании дигетерозигот, которое равно 13:3
Это означает, что здесь наблюдается рецессивный эпистаз. Составим решетку Пеннета. По ней мы можем расщепление при скрещивании дигетерозигот, которое равно 13:3
- Полимерия – взаимодействие неаллельных генов, доминантные аллели которых однозначно влияют на развитие какого-то признака. Кумулятивная полимерия – выраженность признака тем сильнее, чем больше в генотипе доминантных аллелей. Некумулятивная – признак проявляется при наличии хотя бы одного доминантного аллеля. К примеру, если ген с полным доминированием удвоится, то он будет попадать под закономерности некумулятивной полимерии.
Кроме генов, отвечающих за развитие половых признаков, в половых хромосомах есть и другие гены, важные для организмов обоих полов. По определению, сцепленное с полом наследование – это наследование неполовых признаков, за которые отвечают гены, расположенные в половых хромосомах. Специфичность этого типа наследования выражается в том, что у гетерогаметного пола (содержащего в клетках две разные половые хромосомы) признаки, за которые отвечают расположенные в негомологичных участках половых хромосом гены, будут проявляться в любом случае, так как представлены в генотипе в единственном экземпляре.
Пример задачи из заключительного этапа ВОШ по биологии.
У кузнечиков дикого типа цвет тела зеленый, а розовый цвет тела обусловлен рецессивной мутацией гена, локализованного в Х хромосоме. При скрещивании розовой самки с самцом дикого типа в потомстве: а) все самки будут зеленые, а все самцы розовые; б) половина всех самцов и всех самок будут розовые; в) все потомство будет розовым независимо от пола; г) все потомство будет зеленым независимо от пола.
Решение: Большинство насекомых, кроме некоторых исключений, в том числе чешуекрылых, имеют XY половые хромосомы, то есть самцы – гетерогаметный пол у кузнечиков. Следовательно, самцы получают Х-хромосому от матери и только от нее. У матери обе Х-хромосомы содержат мутантный ген, поэтому все самцы будут розовые. Самки в любом случае получат доминантный аллель от отца, и из-за рецессивности признака будут зеленые. Ответ: А
Сцепленное наследование и кроссинговерКаждый организм имеет огромное количество признаков, по сравнению с которым число хромосом невелико.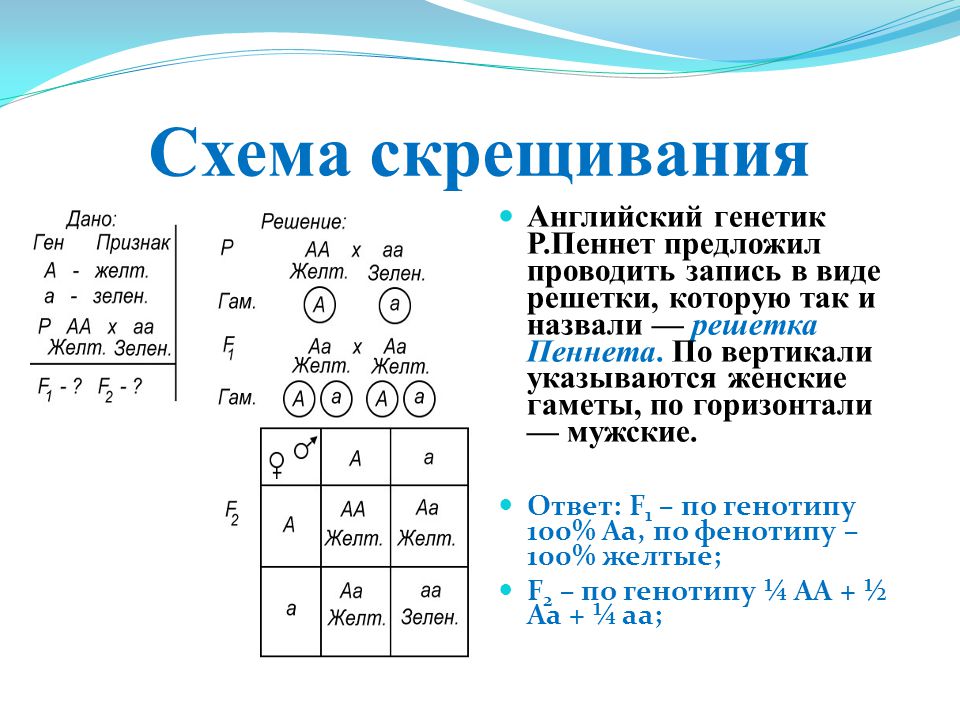 Получается, что каждая хромосома несёт целую группу генов. Гены, которые расположены близко в одной хромосоме, в большинстве случаев наследуются вместе. Однако конъюгация хромосом в профазе мейоза может привести к гомологичной рекомбинации и образованию рекомбинантных генотипов, частота которых будет иной по сравнению с несцепленными генами. Расстояние между генами на хромосоме измеряется на основе частоты образования кроссоверных гамет и, как следствие, потомков с рекомбинантным фенотипом. Если вероятность рекомбинации генов в мейозе составляет 1, то расстояние между ними считается равным 1 сантиморганиде. Обратите внимание, что гены, расстояние между которыми больше 50 сантиморганид, более склонны следовать закономерностям Менделевского наследования, и для них кроссинговером можно пренебречь.
Получается, что каждая хромосома несёт целую группу генов. Гены, которые расположены близко в одной хромосоме, в большинстве случаев наследуются вместе. Однако конъюгация хромосом в профазе мейоза может привести к гомологичной рекомбинации и образованию рекомбинантных генотипов, частота которых будет иной по сравнению с несцепленными генами. Расстояние между генами на хромосоме измеряется на основе частоты образования кроссоверных гамет и, как следствие, потомков с рекомбинантным фенотипом. Если вероятность рекомбинации генов в мейозе составляет 1, то расстояние между ними считается равным 1 сантиморганиде. Обратите внимание, что гены, расстояние между которыми больше 50 сантиморганид, более склонны следовать закономерностям Менделевского наследования, и для них кроссинговером можно пренебречь.
Пример задания из международной олимпиады по биологии 2010 года
Следующая таблица представляет результаты скрещивания растений с тремя сцепленными генами: ген F определяет окраску цветков, ген S определяет окраску семян, ген L определяет высоту растения. У каждого гена имеется по два аллеля, среди которых один аллель проявляет полное доминирование над другим аллелем. Растения с доминантным фенотипом высокие и имеют красные цветки и желтые семена; растения с рецессивным фенотипом низкие и имеют белые цветки и зеленые семена. Допустим, что кроссинговер между двумя генами произошел только один раз.
У каждого гена имеется по два аллеля, среди которых один аллель проявляет полное доминирование над другим аллелем. Растения с доминантным фенотипом высокие и имеют красные цветки и желтые семена; растения с рецессивным фенотипом низкие и имеют белые цветки и зеленые семена. Допустим, что кроссинговер между двумя генами произошел только один раз.
Решение: Сначала разберем скрещивание с рецессивной гомозиготой, которая образует только гаметы вида fs. В этом случае фенотип полностью определяется аллелями, полученными от второго родителя. Так как суммарная частота фенотипов красные цветки/желтые семена и белые цветки/зеленые семена (родительских фенотипов) максимальна, значит у первого родителя сцеплены оба доминантных и оба рецессивных аллеля, и он преимущественно образует гаметы FS и fs. Тогда процент кроссинговера между F и S равен 2, и расстояние между ними в геноме равно 2. Для разбора второго скрещивания нарисуем решетку Пеннета с учетом частот образования гамет.
Низкое растение с зелеными семенами может получиться, только если встретятся две ls гаметы. Вероятность их встречи равна квадрату вероятности образования этих гамет. Значит, гаметы ls образуются с вероятностью 10 %. Пусть гаметы LS, Ls и lS образуются с вероятностью x, y и z соответственно. Запишем в клетки решетки Пеннета вероятности получения этих генотипов. Сложив числа в клетках с фенотипом высокое растение/желтые семена, получим уравнение z2+0.2z=0.24 Единственный положительный корень z=0.4. Аналогично, y=0.4, а х=1-0,1-y-z=0.1 (так как в сумме частоты всех 4 гамет равны 1). Получаем, что у скрещиваемой с самой собой дигетерозиготы хромосомы Ls и lS, а расстояние между ними равно 20.
Необходимо установить истинность следующих утверждений:
- S расположен ближе к L чем к F. Неверно по нашим подсчетам.
- Некоторые высокие растения F1 с зелеными семенами являются результатом кроссинговера. Верно, к примеру с генотипом Llss, где ls образовалось в результате кросинговера.
- Кроссинговер происходит в профазе мейоза I. Верно
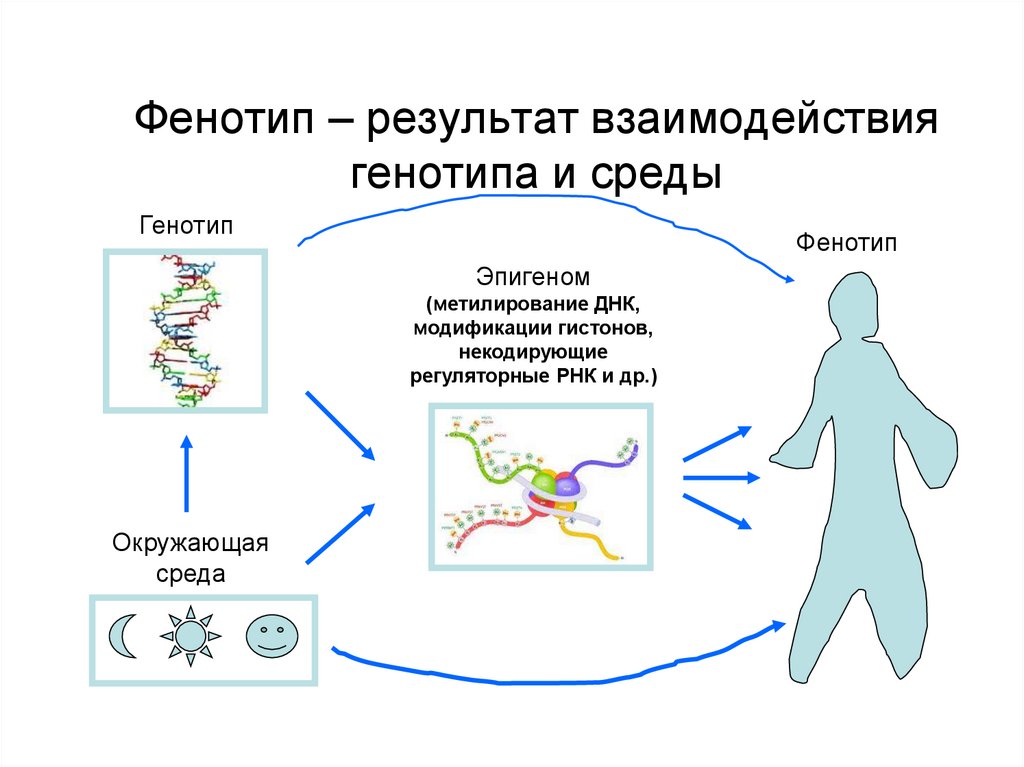
Сколько генотипов можно наблюдать у растений F1, имеющих высокий рост и желтые семена? Четыре: LLSS, LLSs, LlSS, LlSs
Определите расстояние на генетической карте между генами L и S. (Одна единица карты = расстояние в 1% рекомбинации): 20, расчеты выполнены выше.
Хромосомные мутации и геномные перестройкиМутации могут затрагивать не только несколько нуклеотидов, но и структуру хромосом и генома, изменяя взаиморасположение генов и их количество. Типы хромосомных мутаций:
- Делеция – утрата участка хромосомы
- Инверсия – поворот участка хромосомы на 180˚
- Дупликация – удвоение участка хромосомы
- Инсерция – перенос участка хромосомы на другое место
- Транслокация – перенос участка одной хромосомы или целой хромосомы на другую
Хромосомные мутации вызывают хромосомные болезни, к примеру, синдром кошачьего крика, вызываемый делецией в коротком плече хромосомы 15, синдром петрушки (Ангельмана), вызываемый делецией в длинном плече 15 хромосомы, полученной от матери. Такие мутации не подчиняются законам менделевского наследования.
Такие мутации не подчиняются законам менделевского наследования.
Геномные мутации возникают при изменении числа хромосом. Это происходит при нарушении хода митоза и мейоза. Чаще всего происходит увеличение или уменьшение хромосомного набора на 1 хромосому (реже 2 и более). Если хромосома исчезает из диплоидного набора, говорят о нулесомии, остается в единичном экземпляре – моносомия, в тройном – трисомия и так далее. Из-за важной роли фиксированного количества аутосом в онтогнезе, большинство переживших младенчество людей с геномными мутациями несут аномалии в половых хромосомах. Исключение – синдром Дауна (трисомия по 21 хромосоме). Среди аномалий половых хромосом такие синдромы как:
- Синдром Шерешевского-Тернера (моносомия Х-хромосомы)
- Синдром полисомии Х-хромосомы у женщин
- Синдром Кляйнфельтера (полисомия Х и Y-хромосом у лиц мужского пола)
- Синдром полисомии Y-хромосомы.
Задача из отборочного этапа всесибирской олимпиады школьников по биологии за 2014/15 учебный год.
Несколько лет назад в медико-генетическую консультацию г. Новосибирска обратилась супружеская пара, у которой неоднократно рождались нежизнеспособные дети с разнообразными уродствами. Анализ хромосом супругов показал, что они являются примером редчайшего события – они оба носители сбалансированных транслокаций. У жены было обнаружено слияние 21 и 15 хромосом, а у мужа – транслокация между 5 и 9 хромосомами.
Какова вероятность рождения здорового ребенка у этой пары? Какова вероятность того, что все потомки этого ребенка будут здоровы?
Решение: Здоровым ребенок будет в том случае, если обе слившиеся гаметы будут сбалансированы, т. е. либо не иметь транслокаций вообще, либо иметь сбалансированные транслокации.
Вероятность образования яйцеклетки без транслокации – 1/6 (см. таблицу ниже), вероятность образования яйцеклетки с 22 хромосомами (слившимися 21 и 15 хромосомами) – тоже 1/6. Всего вероятность благоприятного кариотипа яйцеклетки 1/3.
Вероятность образования сперматозоида без транслокации 1/4 и сперматозоида с двумя хромосомами с транслокацией (5/9 и 9/5), но без нормальных 9 и 5 хромосом — 1/4. Всего вероятность благоприятного кариотипа сперматозоида – 1/2.
Здоровый ребенок может родиться в результате встречи яйцеклетки и сперматозоида с благоприятными кариотипами. Поэтому вероятность этого события равна произведению посчитанных ранее вероятностей: 1/3 × 1/2 = 1/6
Здоровые потомки у ребенка этой пары будут только в том случае, если его кариотип не будет содержать транслокаций. Вероятность нормального кариотипа у него 1/6 × 1/4 = 1/24, т. е. в 4 раза меньше.
Все возможные результаты мейоза 1 у женщины:
Набор хромосом в исходной клетке: 15 21 Т(15-21)
«1» в таблице означает присутствие хромосомы в клетке, 0 – отсутствие.
Желтые – нормальные гаметы (1/6)
Голубые – с транслокацией Т(15-21), но сбалансированные (1/6)
Все возможные результаты мейоза 1 у мужчины:
Набор хромосом в исходной клетке: 5 9 5/9 9/5
Желтые – нормальные гаметы (1/4)
Голубые – сбалансированные с транслокацией (1/4)
Популяционная генетика – раздел генетики, изучающий изменение частоты аллелей в результате мутагенеза, естественного отбора на репродуктивный успех и случайного перераспределения генов на уровне популяций и видов организмов. Пример таких закономерностей – эффект основателя (снижение генетического разнообразия при формировании новой популяции из малого количества индивидов) и эффект бутылочного горлышка (сокращение генофонда при критическом уменьшении и последующем восстановлении численности популяции). Для вычислений в основном используется закон Харди-Вайенберга, работающий в идеальной популяции (нет естественного отбора, мутагенеза, дрейфа генов, популяция бесконечно большая, замкнутая, а спаривание происходит случайно). Он позволяет связать частоту аллелей с частотой генотипов с помощью формулы p2+2pq+q2=1, где p – частота одного аллеля, q – частота альтернативного аллеля; первое слагаемое в формуле – частота гомозигот по первому аллелю, второе – частота гетерозигот, а третье – частота гомозигот по альтернативному аллелю. Эти величины остаются постоянными и поддерживаются из поколения в поколение. Закон также справедлив в случае множественного аллелизма.
Пример таких закономерностей – эффект основателя (снижение генетического разнообразия при формировании новой популяции из малого количества индивидов) и эффект бутылочного горлышка (сокращение генофонда при критическом уменьшении и последующем восстановлении численности популяции). Для вычислений в основном используется закон Харди-Вайенберга, работающий в идеальной популяции (нет естественного отбора, мутагенеза, дрейфа генов, популяция бесконечно большая, замкнутая, а спаривание происходит случайно). Он позволяет связать частоту аллелей с частотой генотипов с помощью формулы p2+2pq+q2=1, где p – частота одного аллеля, q – частота альтернативного аллеля; первое слагаемое в формуле – частота гомозигот по первому аллелю, второе – частота гетерозигот, а третье – частота гомозигот по альтернативному аллелю. Эти величины остаются постоянными и поддерживаются из поколения в поколение. Закон также справедлив в случае множественного аллелизма.
Пример задания из международной олимпиады 2011 года
На отдаленном острове доктор Йех открыл новый вид растений, цветы у которого либо белые, либо голубые. Растения этого вида главным образом перекрестно опыляются насекомыми. Генетические исследования показали, что белый цвет цветков является рецессивным, а голубой — доминантным. Статистический анализ показал, что 91% этих растений на острове имеют голубые цветки. Если скрестить два случайным образом выбранных растения с голубыми цветками, какова приблизительная вероятность того, что их потомство F1 будет иметь белые цветки?
Растения этого вида главным образом перекрестно опыляются насекомыми. Генетические исследования показали, что белый цвет цветков является рецессивным, а голубой — доминантным. Статистический анализ показал, что 91% этих растений на острове имеют голубые цветки. Если скрестить два случайным образом выбранных растения с голубыми цветками, какова приблизительная вероятность того, что их потомство F1 будет иметь белые цветки?
Решение: 9% растений на острове имеют белые цветки, то есть являются рецессивными гомозиготами. Тогда частота аллеля белых цветков в популяции 0,09=0,3. Следовательно, частота аллеля голубых цветков 1-0,3=0,7. По закону Харди-Вайенберга, считая популяцию идеальной, частота гетерозигот 2*0,3*0,7=0,42. От двух случайно выбранных растений с голубыми цветами можно получить потомство с белыми цветками, только если оба родителя гетерозиготы. Вероятность этого события представляет собой квадрат вероятности случайного выбора гетерозиготы из голубых растений, которая равна отношению частоты встречаемости гетерозиготного генотипа и голубоцветочного фенотипа, то есть (6/13)2≈0,21
ЗаключениеАвтор надеется, что данный материал помог систематизировать существующие знания и закрыть пробелы в сферах, в которых читатели не были уверены.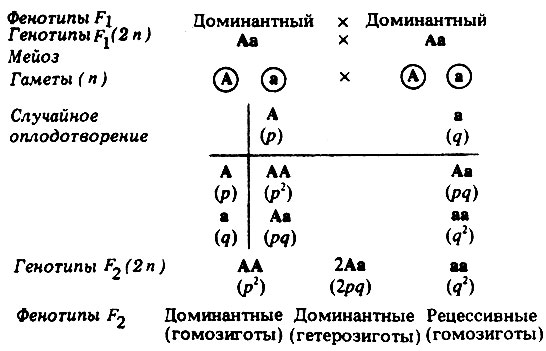 Удачи в дальнейшем изучении биологии и участии в соревнованиях и олимпиадах!
Удачи в дальнейшем изучении биологии и участии в соревнованиях и олимпиадах!
# Генетика
# 11 класс
Влияние генотипа и среды на фенотип
Напомним еще раз смысл понятий генотипа и фенотипа. Генотип — это совокупность всех генов данного организма; фенотип — это совокупность всех признаков организма.
Известно, что при одном и том же фенотипе организмы могут иметь разный генотип. Например, в опытах Менделя растения, генотип которых содержал аллели AA, и растения, генотип которых содержал аллели Aа, по фенотипу не отличались друг от друга. Может ли быть обратная ситуация, когда генотипы у организмов одинаковые, а фенотипы разные? В частности, в какой мере фенотип определяется генотипом, а в какой — влияниями среды? Этот вопрос часто обсуждается на бытовом уровне применительно к характеру или поведению людей. При этом бытуют две точки зрения.
Согласно одной из них, особенности человека целиком определяются его
генотипом.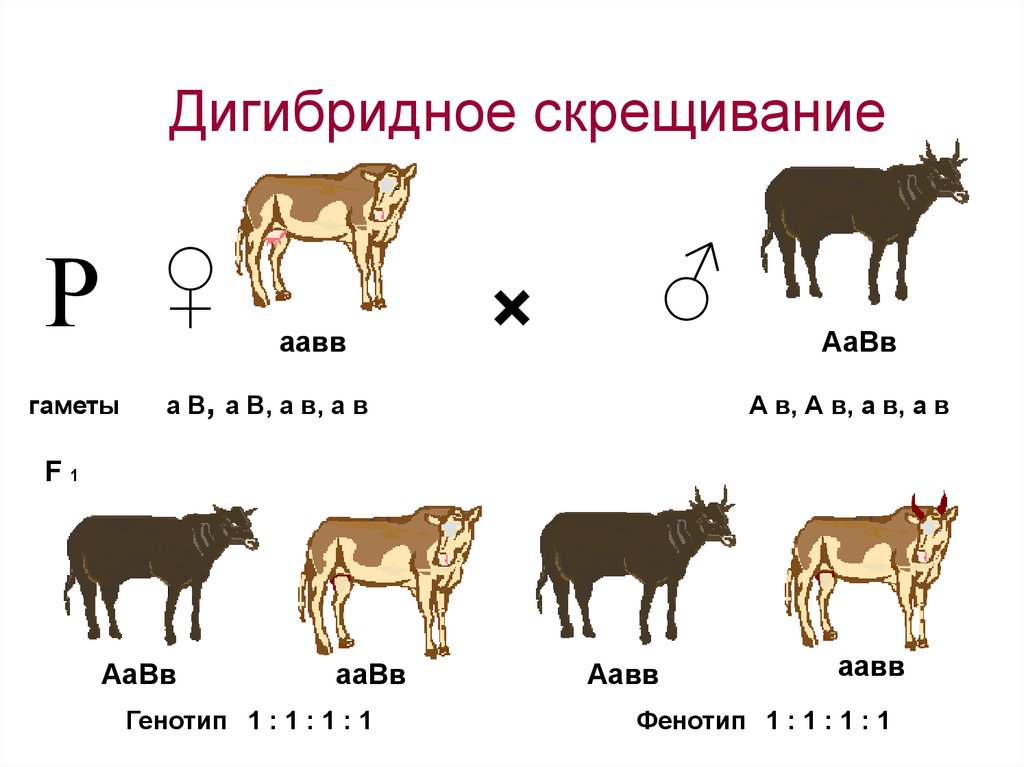 Поведение задано наследственностью, с которой ничего нельзя
поделать. Согласно другой точке зрения, наследственность в поведении людей
играет незначительную роль по сравнению с условиями жизни и, особенно, с
воспитанием.
Поведение задано наследственностью, с которой ничего нельзя
поделать. Согласно другой точке зрения, наследственность в поведении людей
играет незначительную роль по сравнению с условиями жизни и, особенно, с
воспитанием.
Рассмотрим влияние наследственности и среды на более простые признаки, чем поведение людей. Даже для таких признаков возможны разные варианты.
— Некоторые признаки полностью определяются генотипом и не зависят от условий среды. К ним относятся, например, группы крови и многие генетические заболевания .
— Другие признаки зависят и от генотипа и от среды. Например, рост
человека зависит от его генотипа (вспомните работу
Гальтона
). В то же время рост зависит и от условий среды, в частности от питания в
период роста. Цвет кожи в значительной мере определяется генотипом. Но цвет
кожи людей с одинаковым генотипом очень сильно зависит от времени их
пребывания на солнце (
рис. 122
).
Рассмотрим несколько характерных примеров влияния среды на проявления генов.
1. Еще на самом раннем периоде развития генетики было обнаружено, что признак может оказаться доминантным или рецессивным в зависимости от условий, в которых развивается организм. В 1915 г. Морган показал на дрозофиле, что при выращивании в сухом воздухе обычное для «дикого» типа распределение полос на брюшке дрозофилы доминирует над ненормальным и, напротив, при избыточной влажности доминирует ненормальное распределение полос. Наблюдения такого рода еще раз показали различия между генотипом и фенотипом: при одном и том же генотипе фенотип зависел от внешнцх условий.
2. Влияние внешней среды на фенотип можно продемонстрировать на примере
общественных насекомых. У пчел и муравьев из неоплодотворенных яиц
развиваются самцы, а из оплодотворенных — самки. Однако фенотип этих самок
зависит от условий развития: при одних условиях развивается плодовитая
самка, а при других — бесплодная рабочая пчела.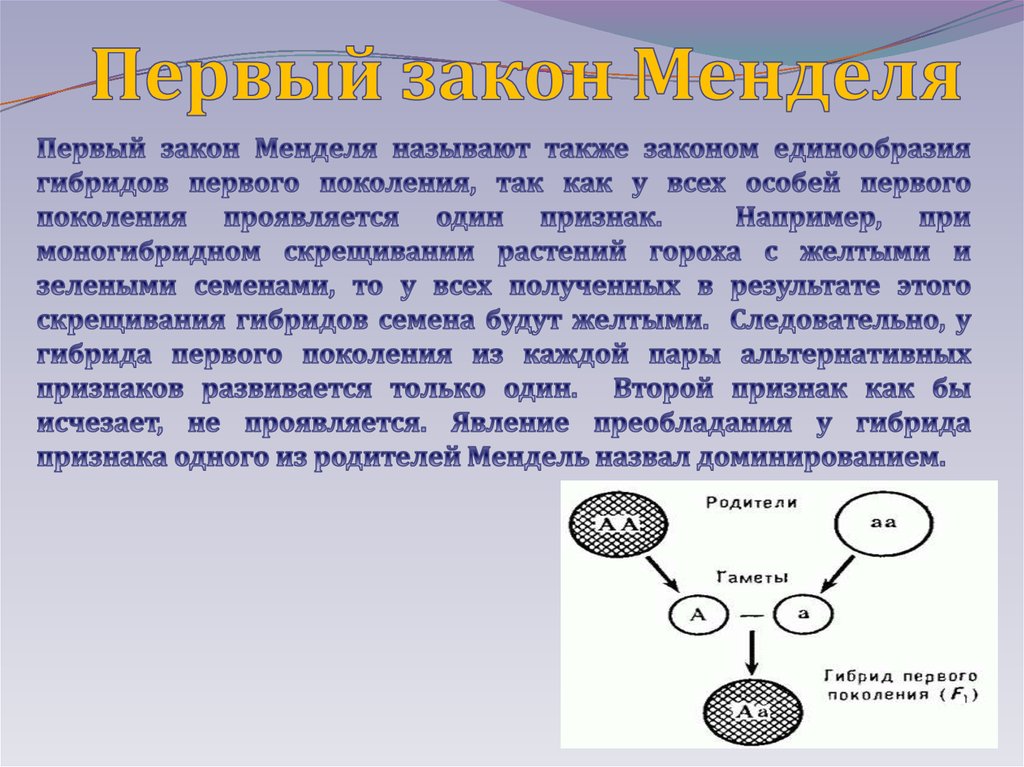 У
муравьев
существуют разные «касты» бесплодных особей. Основную часть
населения муравейника составляют рабочие муравьи, которые строят
муравейник, добывают пищу, выкармливают личинок и выполняют всякую другую
работу. У многих видов муравьев имеются «солдаты» — муравьи с
крупной головой, защищенной толстым хитином, и с особо мощными челюстями.
Рабочие муравьи и солдаты — это недоразвитые самки, они бесплодны. Почему
же из одних яиц, отложенных самкой, выводятся рабочие особи, из других —
солдаты, из третьих — крылатые половые особи: самцы и самки? Еще в 1910 г.
исследователь муравьев
Вассман
убрал из гнезда самку. Оказалось, что после этого рабочие муравьи
начинают откладывать яйца! Этим опытом было показано, что наличие самки
тормозит откладку яиц рабочими особями. При дальнейшем изучении выяснилось,
что кроме веществ, которые тормозят развитие новых самок, в муравейнике
циркулируют вещества, которые наоборот стимулируют развитие яичников у
рабочих особей и у личинок.
У
муравьев
существуют разные «касты» бесплодных особей. Основную часть
населения муравейника составляют рабочие муравьи, которые строят
муравейник, добывают пищу, выкармливают личинок и выполняют всякую другую
работу. У многих видов муравьев имеются «солдаты» — муравьи с
крупной головой, защищенной толстым хитином, и с особо мощными челюстями.
Рабочие муравьи и солдаты — это недоразвитые самки, они бесплодны. Почему
же из одних яиц, отложенных самкой, выводятся рабочие особи, из других —
солдаты, из третьих — крылатые половые особи: самцы и самки? Еще в 1910 г.
исследователь муравьев
Вассман
убрал из гнезда самку. Оказалось, что после этого рабочие муравьи
начинают откладывать яйца! Этим опытом было показано, что наличие самки
тормозит откладку яиц рабочими особями. При дальнейшем изучении выяснилось,
что кроме веществ, которые тормозят развитие новых самок, в муравейнике
циркулируют вещества, которые наоборот стимулируют развитие яичников у
рабочих особей и у личинок. Эти вещества вырабатываются специальными
железами рабочих муравьев. В нормальных условиях рабочие муравьи кормят
этими веществами матку и личинок, из которых развиваются самцы и самки.
Если матки в муравейнике нет, эти вещества получают в основном личинки.
Если же личинок мало, то рабочие муравьи кормят этими веществами друг друга
и тогда начинают откладывать яйца. Таким образом, было выяснено, что
развитие личинок зависит от того, какой корм они получат от рабочих
муравьев и какие добавки окажутся в корме. Точно так же у
пчел
от характера пищи и добавок зависит, разовьется личинка в рабочую пчелу
или же в матку.
Эти вещества вырабатываются специальными
железами рабочих муравьев. В нормальных условиях рабочие муравьи кормят
этими веществами матку и личинок, из которых развиваются самцы и самки.
Если матки в муравейнике нет, эти вещества получают в основном личинки.
Если же личинок мало, то рабочие муравьи кормят этими веществами друг друга
и тогда начинают откладывать яйца. Таким образом, было выяснено, что
развитие личинок зависит от того, какой корм они получат от рабочих
муравьев и какие добавки окажутся в корме. Точно так же у
пчел
от характера пищи и добавок зависит, разовьется личинка в рабочую пчелу
или же в матку.
3. У горностаевых кроликов мех белый, но отдельные части тела — лапы, уши,
кончик морды и хвост — черные. Если на спине кролика, которая покрыта белой
шерстью, выстричь какой-то участок и содержать кролика при пониженной
температуре, на этом участке вырастает черная шерсть. Конечно, такие пятна
черной окраски на необычном месте потомкам этого кролика по наследству не
передаются.
Приведенные примеры показывают, что в действительности в очень многих случаях наследуется не признак как таковой, а способность к развитию данного признака при соответствующих условиях внешней среды, которая и передается от поколения к поколению.
Рассмотрим еще раз понятие чистая линия . Группы животных и растений, потомки которых в ряде поколений не изменяются по внешнему виду и не дают расщепления, названы чистыми линиями (иногда это понятие применяют только к потомству самоопылителей). Теперь можно дополнить определение: организмы, относящиеся к чистой линии, гомозиготны по аллелям , определяющим изучаемые признаки.
Датский генетик
Иоганнсен
поставил опыты для определения возможности отбора в чистых линиях. Он
видел, что у данного растения — гороха, принадлежащего к чистой линии,
имелись горошины разного размера: мелкие, средние и крупные. Иоганнсен
сажал самые мелкие горошины и самые крупные и получал от них потомство.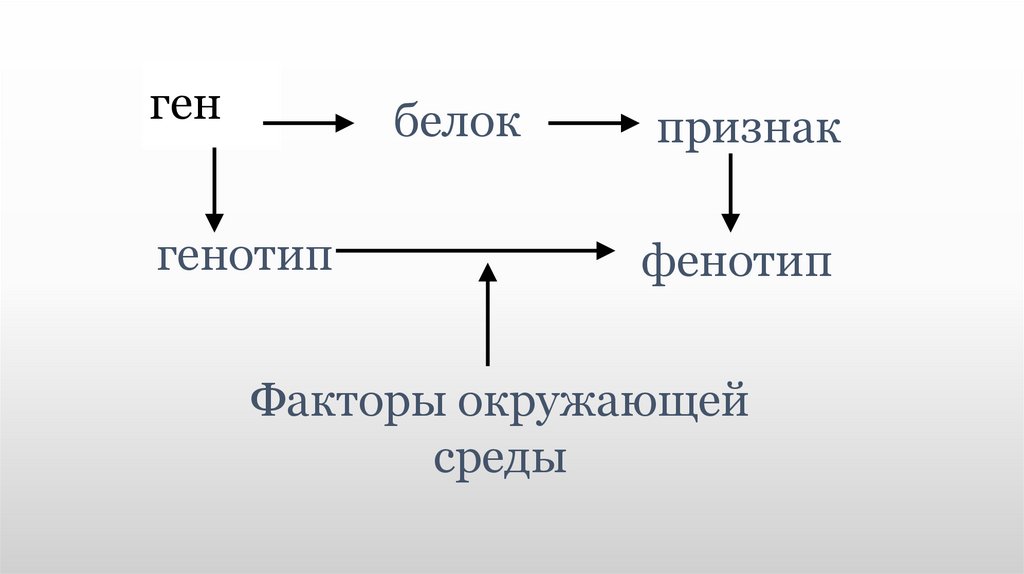 С
растений, выросших из самых мелких горошин, брались опять самые мелкие, а с
растений, выросших из крупных горошин, брались самые крупные. После такой
процедуры, проводившейся для ряда поколений, оказалось, что соотношение
горошин разного размера (мелких, средних и крупных) было одинаковым у
отбираемых растений, выросших из самых мелких семян, и выросших из самых
крупных семян; при этом оно не отличалось от соотношения, которое было у
исходного родительского растения. Размеры горошин были обусловлены разными
случайными причинами (одни формировались, когда было больше солнца, другие,
когда было больше влаги и т.д.). Но генотип у всех растений был одинаков и
отбор не мог изменить соотношение размера горошин. Это показало, что
селекционерам нецелесообразно вести отбор среди потомков чистых линий.
Разброс размеров горошин, возникавший под влиянием случайных причин,
подчинялся определенной закономерности. Больше всего было горошин
некоторого среднего размера. Меньше всего было особенно мелких и особенно
крупных горошин.
С
растений, выросших из самых мелких горошин, брались опять самые мелкие, а с
растений, выросших из крупных горошин, брались самые крупные. После такой
процедуры, проводившейся для ряда поколений, оказалось, что соотношение
горошин разного размера (мелких, средних и крупных) было одинаковым у
отбираемых растений, выросших из самых мелких семян, и выросших из самых
крупных семян; при этом оно не отличалось от соотношения, которое было у
исходного родительского растения. Размеры горошин были обусловлены разными
случайными причинами (одни формировались, когда было больше солнца, другие,
когда было больше влаги и т.д.). Но генотип у всех растений был одинаков и
отбор не мог изменить соотношение размера горошин. Это показало, что
селекционерам нецелесообразно вести отбор среди потомков чистых линий.
Разброс размеров горошин, возникавший под влиянием случайных причин,
подчинялся определенной закономерности. Больше всего было горошин
некоторого среднего размера. Меньше всего было особенно мелких и особенно
крупных горошин. Распределение горошин по размеру является одним из
примеров нормального распределения.
Распределение горошин по размеру является одним из
примеров нормального распределения.
Вернемся теперь к поведению человека. Здесь возникают важные вопросы, которые давно вызывают споры. Например, бывает ли человек от рождения умным или глупым? Бывают ли врожденные преступники? Или ум — результат хорошего воспитания, а преступность — результат плохого. Однако ответы на эти вопросы очень трудны. Во-первых, трудно измерять уровень интеллекта человека и характеристики его поведения. Во-вторых, трудно выяснить, какие гены имеют отношение к поведению и как различаются люди по этим генам. В-третьих, трудно сравнить или уравнять условия воспитания разных людей.
Тем не менее, некоторые результаты исследований этой проблемы заслуживают
внимания, например, полученные в работах по изучению наследования
интеллекта. Для определения уровня интеллекта разработан ряд тестов.
Применение этих тестов к близким родственникам, которые воспитывались
вместе или отдельно, и к неродственным людям, которые воспитывались вместе
или отдельно, показало следующее. Во-первых, чем ближе родство людей, тем
ближе их уровни интеллекта, даже если они воспитываются раздельно. Особенно
сходны между собой однояйцевые близнецы (Идея использования близнецов для
генетических исследований была предложена
Ф.Гальтоном
). Это значит, что генотип играет заметную роль в определении интеллекта.
Во-вторых, не родственники, воспитанные вместе, имеют более сходные
показатели интеллекта, чем такие же не родственники, воспитывающиеся
раздельно. Это показывает, что и среда (воспитание) отчасти определяет
уровень интеллекта. Для большинства людей влияние наследственности и среды
сопоставимо.
Во-первых, чем ближе родство людей, тем
ближе их уровни интеллекта, даже если они воспитываются раздельно. Особенно
сходны между собой однояйцевые близнецы (Идея использования близнецов для
генетических исследований была предложена
Ф.Гальтоном
). Это значит, что генотип играет заметную роль в определении интеллекта.
Во-вторых, не родственники, воспитанные вместе, имеют более сходные
показатели интеллекта, чем такие же не родственники, воспитывающиеся
раздельно. Это показывает, что и среда (воспитание) отчасти определяет
уровень интеллекта. Для большинства людей влияние наследственности и среды
сопоставимо.
Ссылки:
- Модификационная изменчивость
- ГЕНОТИП И ФЕНОТИП
Дифференциальный взгляд на отношения генотип-фенотип
- Список журналов
- Фронт Жене
- PMC4437230
Передняя жен. 2015 г.; 6: 179.
2015 г.; 6: 179.
Опубликовано в Интернете 19 мая 2015 г. doi: 10.3389/fgene.2015.00179
, 1, * , 2 и 3
Информация об авторе Примечания к статье Информация об авторском праве и лицензии Отказ от ответственности между генотипами и фенотипами. Здесь мы вновь подчеркиваем старую точку зрения, согласно которой отношения генотип-фенотип (GP) лучше всего рассматривать как связь между двумя различиями, одно на генетическом уровне, а другое на фенотипическом уровне. На сегодняшний день в биологических исследованиях преобладает мнение, что несколько генов взаимодействуют с несколькими переменными окружающей среды (такими как абиотические факторы, культура или симбионты) для создания фенотипа. Часто проблема связи генотипов и фенотипов формулируется в терминах карт генотипов и фенотипов, и такие графические представления неявным образом уводят нас от дифференциального взгляда на отношения ГП. Здесь мы показываем, что дифференциальный взгляд на отношения GP является полезной объяснительной основой в контексте всепроникающей плейотропии, эпистаза и эффектов окружающей среды. В таких случаях уместно рассматривать отношения ГП как различия, встроенные в различия. Мышление с точки зрения различий проясняет сравнение между влиянием окружающей среды и генетики на фенотипы и помогает лучше понять связь между генотипами и фенотипами.
В таких случаях уместно рассматривать отношения ГП как различия, встроенные в различия. Мышление с точки зрения различий проясняет сравнение между влиянием окружающей среды и генетики на фенотипы и помогает лучше понять связь между генотипами и фенотипами.
Ключевые слова: генотип, фенотип, генетика, сложный признак, GxE, GxG. человек человеком». В противовес0041 генотип , наследственный материал, передаваемый гаметами. Начиная с раннего предположения, что генотипы и фенотипы образуют два фундаментально разных уровня биологической абстракции (Johannsen, 1911), задача состояла в том, чтобы понять, как они взаимодействуют друг с другом, как генотипы отображаются на фенотипы. За последние 15 лет более 1000 примеров изменений последовательности ДНК были связаны с естественными невредными фенотипическими различиями между особями или видами у эукариот (Martin and Orgogozo, 2013b). У человека OMIM 9Каталог 0015 ® (онлайн-менделевское наследование у человека, http://omim. org/), в котором собраны генетические детерминанты фенотипов, связанных с заболеванием, насчитывает более 4300 записей и в общей сложности 2493 опубликованных полногеномных ассоциативных исследования (GWAS). обнаружение множества участков в геноме, которые статистически связаны со сложными признаками (Welter et al., 2014). Поскольку обнаружение причинно-следственных связей между генетическими и фенотипическими вариациями ускоряется, пересмотр наших концептуальных инструментов может помочь нам найти объединяющие принципы в рое данных. Здесь мы размышляем о взаимосвязи между генотипами и фенотипами и адресуем это эссе биологам, которые готовы попытаться бросить вызов своему нынешнему пониманию фенотипов. Выделим одну полезную точку зрения — дифференциальную. Затем мы показываем, что эта простая структура остается полезной в контексте всепроникающей плейотропии, эпистаза и воздействия окружающей среды.
org/), в котором собраны генетические детерминанты фенотипов, связанных с заболеванием, насчитывает более 4300 записей и в общей сложности 2493 опубликованных полногеномных ассоциативных исследования (GWAS). обнаружение множества участков в геноме, которые статистически связаны со сложными признаками (Welter et al., 2014). Поскольку обнаружение причинно-следственных связей между генетическими и фенотипическими вариациями ускоряется, пересмотр наших концептуальных инструментов может помочь нам найти объединяющие принципы в рое данных. Здесь мы размышляем о взаимосвязи между генотипами и фенотипами и адресуем это эссе биологам, которые готовы попытаться бросить вызов своему нынешнему пониманию фенотипов. Выделим одну полезную точку зрения — дифференциальную. Затем мы показываем, что эта простая структура остается полезной в контексте всепроникающей плейотропии, эпистаза и воздействия окружающей среды.
Мутации, выделенные из лабораторных штаммов, сыграли важную роль в понимании карты GP. В классической схеме мутацию сравнивают с эталоном дикого типа, и ее фенотипические эффекты используются для вывода о функции гена. Эта структура часто приводит к семантическому сокращению: от генетического изменения, вызывающего вариацию фенотипа, часто бывает удобно ассимилировать соответствующий ген в качестве причинной детерминанты признака (Keller, 2010; Figure ). Часто можно встретить заголовки, выражающие эти упрощения, трубящие перед широкой аудиторией об открытии гена «долголетия» или «благополучия», которые приносят научную точность в жертву психологическому воздействию. В соответствии с этим следует ли называть ген, мутация которого летальна, «геном жизни»? На самом деле эти упрощенные формулировки означают, что вариация данного гена вызывает вариацию данного фенотипа (Dawkins, 1982; Schwartz, 2000; Waters, 2007). На самом деле ген сам по себе не может ни вызвать наблюдаемый фенотипический признак, ни быть необходимым и достаточным для появления наблюдаемых признаков.
В классической схеме мутацию сравнивают с эталоном дикого типа, и ее фенотипические эффекты используются для вывода о функции гена. Эта структура часто приводит к семантическому сокращению: от генетического изменения, вызывающего вариацию фенотипа, часто бывает удобно ассимилировать соответствующий ген в качестве причинной детерминанты признака (Keller, 2010; Figure ). Часто можно встретить заголовки, выражающие эти упрощения, трубящие перед широкой аудиторией об открытии гена «долголетия» или «благополучия», которые приносят научную точность в жертву психологическому воздействию. В соответствии с этим следует ли называть ген, мутация которого летальна, «геном жизни»? На самом деле эти упрощенные формулировки означают, что вариация данного гена вызывает вариацию данного фенотипа (Dawkins, 1982; Schwartz, 2000; Waters, 2007). На самом деле ген сам по себе не может ни вызвать наблюдаемый фенотипический признак, ни быть необходимым и достаточным для появления наблюдаемых признаков. Гены нуждаются в клеточной среде, совместном действии множества других генов, а также в определенных физико-химических условиях, чтобы оказывать заметное влияние на организмы ( Рисунок ). Например, пигментация каштановых волос у одного человека является результатом не только генов, кодирующих ферменты, синтезирующие пигмент, но и наличия клеток, продуцирующих пигменты соответствующих молекул субстрата (таких как тирозин для меланина), и количества полученного вещества. солнечный свет (Liu et al., 2013). Таким образом, генетический редукционистский подход, который исследует лишь несколько генетических параметров среди множества причинных факторов, бесполезен для полного решения широкого вопроса о том, что делает волосы коричневыми, что порождает определенную биологическую структуру или процесс в целом. Тем не менее, генетический редукционизм может быть вполне подходящим для идентификации генетических локусов, где изменение вызывает фенотипическое различие (9).0029 Рисунок ).
Гены нуждаются в клеточной среде, совместном действии множества других генов, а также в определенных физико-химических условиях, чтобы оказывать заметное влияние на организмы ( Рисунок ). Например, пигментация каштановых волос у одного человека является результатом не только генов, кодирующих ферменты, синтезирующие пигмент, но и наличия клеток, продуцирующих пигменты соответствующих молекул субстрата (таких как тирозин для меланина), и количества полученного вещества. солнечный свет (Liu et al., 2013). Таким образом, генетический редукционистский подход, который исследует лишь несколько генетических параметров среди множества причинных факторов, бесполезен для полного решения широкого вопроса о том, что делает волосы коричневыми, что порождает определенную биологическую структуру или процесс в целом. Тем не менее, генетический редукционизм может быть вполне подходящим для идентификации генетических локусов, где изменение вызывает фенотипическое различие (9).0029 Рисунок ). Различие в цвете волос между двумя людьми в некоторых случаях может быть связано с их генетическими различиями. Заметим, однако, что не все фенотипические изменения можно отнести к генетическим изменениям. Разница в цвете волос также может быть вызвана негенетическими факторами, такими как возраст, интенсивность солнечного излучения или окраска волос, или сочетанием генетических и негенетических различий.
Различие в цвете волос между двумя людьми в некоторых случаях может быть связано с их генетическими различиями. Заметим, однако, что не все фенотипические изменения можно отнести к генетическим изменениям. Разница в цвете волос также может быть вызвана негенетическими факторами, такими как возраст, интенсивность солнечного излучения или окраска волос, или сочетанием генетических и негенетических различий.
Открыть в отдельном окне
Схематические изображения звеньев ГП. (А) Традиционное представление в классическом генетическом редукционизме. (B) Интегративный взгляд на биологию развития. (C) Схема экспериментального подхода в генетике. (D) Схема экспериментального подхода в эволюционной генетике. (E) Один из примеров таксономически надежной связи GP: SLC45A2 и внутривидовые различия в пигментации у тигров и кур. Отношения ГП обозначены пунктирными линиями на панелях 9. 0029 (Германия) .
0029 (Германия) .
В то время как современная генетика находилась в зачаточном состоянии, Альфред Стертевант сформулировал вопрос о карте ВП простыми словами: «одной из центральных проблем биологии является проблема дифференцировки — как яйцо развивается в сложный многоклеточный организм? Это, конечно, традиционная главная проблема эмбриологии; но он также появляется в генетике в форме вопроса: как гены производят свои эффекты?» (Стёртевант, 1932). Долгое время некоторые генетики, возможно, думали, что они анализируют морфогенетические механизмы, лежащие в основе формирование фенотипических признаков, в то время как их экспериментальный подход фактически заключался в выявлении генов, отсутствие или изменение которых (мутации, делеции, дупликации, перестройки и т. д.) приводит к фенотипическим различиям (сравните рис. с рис. ). На самом деле предложение «ваши волосы каштановые» можно интерпретировать либо как абсолютное наблюдение (описание определенного набора молекул, содержащих определенные уровни темного пигмента эумеланина и светлого пигмента феомеланина), либо как имплицитную ссылку на другие возможности. (он коричневый, а не другого цвета). Заблуждения возникают из-за того, что фенотипы обычно определяются относительно возможностей, которые не сформулированы явно. Наше сознание и наш язык часто склонны путать объекты, изменчивость которых рассматривается, с самой изменчивостью (Keller, 2010), и важно напомнить, что в генетике интересующие нас объекты (например, данный генотип, аллель или фенотип) заслуживают определения относительно к другому эталонному состоянию.
(он коричневый, а не другого цвета). Заблуждения возникают из-за того, что фенотипы обычно определяются относительно возможностей, которые не сформулированы явно. Наше сознание и наш язык часто склонны путать объекты, изменчивость которых рассматривается, с самой изменчивостью (Keller, 2010), и важно напомнить, что в генетике интересующие нас объекты (например, данный генотип, аллель или фенотип) заслуживают определения относительно к другому эталонному состоянию.
Таким образом, классический генетический редукционистский подход по своей сути неспособен объяснить все факторы, ответственные за наблюдаемые характеристики в живом мире (Stotz, 2012), но является мощным и актуальным методом анализа генетических рычагов наследуемой фенотипической изменчивости. Сосредоточение внимания на фенотипической изменчивости между особями, а не на абсолютных признаках, присутствующих в отдельных организмах, является ключом к лучшему пониманию генетических причин фенотипического разнообразия.
Мышление с точки зрения различий делает очевидным абстрактную сущность, которая включает в себя как генетический, так и фенотипический уровни. Этот объект состоит из вариации генетического локуса (два аллеля), связанного с ним фенотипического изменения (два различных фенотипических состояния) и их взаимосвязей ( Рисунок ). Мы втроем называем совокупность этих элементов «гефе», но здесь мы называем это просто «отношением генотип-фенотип» (отношением ГП). Мы покажем, что отношения ГП — это гораздо больше, чем простое и слабо определенное взаимодействие между двумя уровнями организации: это причинно-следственная связь, которая облегчает наше понимание фенотипического разнообразия.
Генетическая часть взаимосвязи GP
В современных базах данных аннотаций генома ген обычно определяется как фрагмент нуклеиновых кислот, который транскрибируется и кодирует РНК или полипептид с известной или предполагаемой функцией (Gerstein et al. др., 2007). Генетический локус, лежащий в основе фенотипических различий, не обязательно является геном в строгом смысле; он может охватывать определенную пару оснований, кодирующую область, регуляторную область цис- или распространяться на весь ген с его цис -регуляторные области или даже кластер генов ( Таблица ). Как ранее отмечалось другими (Falk, 1984; Gilbert, 2000; Stern, 2000; Moss, 2003; Griffiths and Stotz, 2013), концепция гена в биологии развития и в современных базах данных аннотаций генома отличается от концепции гена в эволюционная биология. Здесь акцент делается не на сам ген, как он определен в геномных базах данных, а скорее на индивидуальном функциональном разделении генома на определяющие различия локусы. Генотипическая часть родства GP может принимать форму различных аллелей: разные кодоны, кодирующие разные аминокислоты, вставки/делеции в последовательности, кодирующей белок, расходящиеся версии конкретных цис- регуляторный элемент, наличие/отсутствие вставок транспозонов, количество копий генов в пределах генного кластера, склонного к структурной изменчивости, и т.
др., 2007). Генетический локус, лежащий в основе фенотипических различий, не обязательно является геном в строгом смысле; он может охватывать определенную пару оснований, кодирующую область, регуляторную область цис- или распространяться на весь ген с его цис -регуляторные области или даже кластер генов ( Таблица ). Как ранее отмечалось другими (Falk, 1984; Gilbert, 2000; Stern, 2000; Moss, 2003; Griffiths and Stotz, 2013), концепция гена в биологии развития и в современных базах данных аннотаций генома отличается от концепции гена в эволюционная биология. Здесь акцент делается не на сам ген, как он определен в геномных базах данных, а скорее на индивидуальном функциональном разделении генома на определяющие различия локусы. Генотипическая часть родства GP может принимать форму различных аллелей: разные кодоны, кодирующие разные аминокислоты, вставки/делеции в последовательности, кодирующей белок, расходящиеся версии конкретных цис- регуляторный элемент, наличие/отсутствие вставок транспозонов, количество копий генов в пределах генного кластера, склонного к структурной изменчивости, и т. д. В геноме не все нуклеотидные участки связаны с фенотипической изменчивостью. Например, вероятно, существуют фрагменты нуклеотидных последовательностей, в том числе так называемая мусорная ДНК (Graur et al., 2015), присутствие которых не оказывает никакого влияния на наблюдаемые характеристики организма, кроме того, что они реплицируются и, возможно, транскрибируются. Существуют также генетические локусы, которые могли быть связаны с фенотипической изменчивостью в прошлом и больше не связаны с фенотипической изменчивостью. Например, генетическая изменчивость в кодирующих областях, связывающих гистоновую ДНК, могла быть важной во время ранней эволюции эукариотических клеток, но эти генетические локусы больше не содержат фенотипически релевантных вариаций, кроме летальных мутаций. Таким образом, в геноме есть участки нуклеотидов, которые абсолютно необходимы для жизни, но сами по себе не содержат жизнеспособных фенотипически релевантных вариаций.
д. В геноме не все нуклеотидные участки связаны с фенотипической изменчивостью. Например, вероятно, существуют фрагменты нуклеотидных последовательностей, в том числе так называемая мусорная ДНК (Graur et al., 2015), присутствие которых не оказывает никакого влияния на наблюдаемые характеристики организма, кроме того, что они реплицируются и, возможно, транскрибируются. Существуют также генетические локусы, которые могли быть связаны с фенотипической изменчивостью в прошлом и больше не связаны с фенотипической изменчивостью. Например, генетическая изменчивость в кодирующих областях, связывающих гистоновую ДНК, могла быть важной во время ранней эволюции эукариотических клеток, но эти генетические локусы больше не содержат фенотипически релевантных вариаций, кроме летальных мутаций. Таким образом, в геноме есть участки нуклеотидов, которые абсолютно необходимы для жизни, но сами по себе не содержат жизнеспособных фенотипически релевантных вариаций.
Таблица 1
Несколько примеров отношений GP.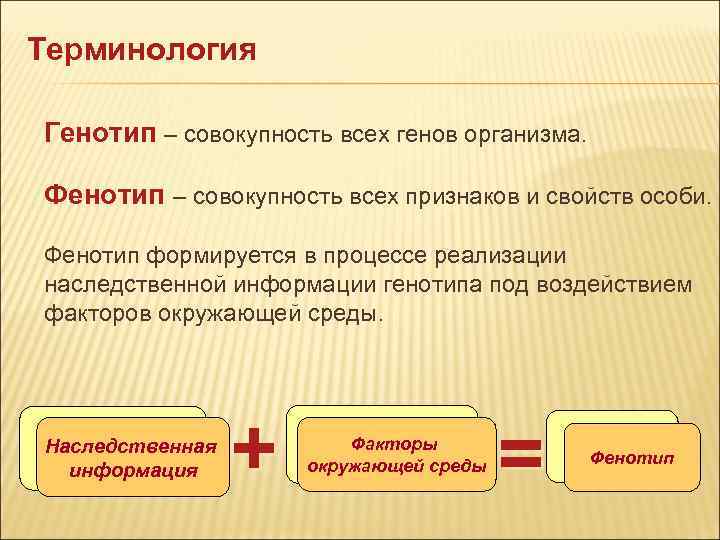
| Genetic locus | Phenotypic trait | Organisms | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Two coding sites in the ABO HBGG gene | A/B blood group | Human, chimpanzee, gibbon | ||||||||||||||||||||||||||||||||||||
| Цис -регуляторный элемент в гене лактазы | Способность переваривать молоко | Различные популяции человека | ||||||||||||||||||||||||||||||||||||
| Количество дупликаций генов амилазы AMY1 или AMY2B | Способность переваривать крах.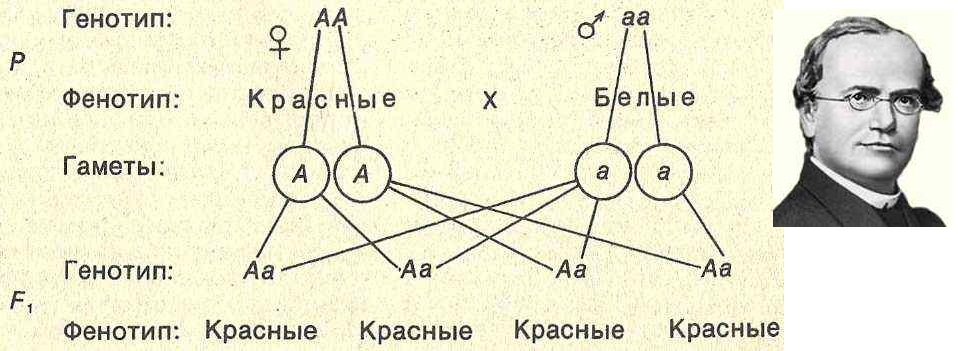 синтазы синтазы | Способность продуцировать каротиноиды | Гороховая тля, паутинный клещ, галлица, нематода | |||||||||||||||||||||||||||||||||||
| Кодирующие сайты в генах опсина | Цветовое зрение | ЧЕЛОВЕКА, Китоисты, Рыбы, Бабочки | ||||||||||||||||||||||||||||||||||||
| Кодирующая область F 3’5’h | Цветочная пигментация | соевые сои, горох, годовой Phlox, Potato, 37| . |  FRIGIDA FRIGIDA Flowering time | Thale cress, oilseed rape | | |||||||||||||||||||||||||||||||||
| Coding region of BADh3 | Fragrance | Soybean, rice | ||||||||||||||||||||||||||||||||||||
| Coding region of HMA3 | Heavy metal tolerance | Thale cress, rice | ||||||||||||||||||||||||||||||||||||
| Coding and cis -regulatory regions of myostatin | Muscle size | Cattle, sheep, dog, pig, horse, human | ||||||||||||||||||||||||||||||||||||
| Цис -регуляторные области в комплексе ахете-щитка | Количество и положение сенсорных щетинок | 0041 pitx1 genePelvis morphology | Stickleback fish | |||||||||||||||||||||||||||||||||||
| Coding region of SLC45A2 | Pigmentation of eye, hair, and skin | Human, tiger, chicken | ||||||||||||||||||||||||||||||||||||
| Coding region of Mc1R | Пигментация волос и кожи, но не глаз | Человек, мышь, крупный рогатый скот, курица, морская свинка, лошадь, лиса, свинья, овца, собака, кролик, медведь, ягуар, ягуарунди, белка, птицы, прыткая ящерица | ||||||||||||||||||||||||||||||||||||
| Cis -regulatory regions of tan and ebony | Pigmentation pattern | Fruitflies | ||||||||||||||||||||||||||||||||||||
| Cis -regulatory elements in the shavenbaby gene | Position and number of trichomes | Плодовые мушки | ||||||||||||||||||||||||||||||||||||
| Цис — регуляторные элементы в гене optix | Красный рисунок на крыльях бабочки | Длиннокрылые бабочки | ||||||||||||||||||||||||||||||||||||
| Coding region of hemoglobin alpha and beta chain genes | Resistance to hypoxia | Human, llama, crocodile, deer mouse, waterfowl | ||||||||||||||||||||||||||||||||||||
| Coding region of Ace | Resistance to organophosphate insecticides | Картофельный жук, тля, комары, комнатная муха, плодовая муха, восточная плодовая муха | ||||||||||||||||||||||||||||||||||||
| Наличие/отсутствие вставки CypA в0041 TRIM5a gene | Resistance to retrovirus | Owl monkey, macaque, Old World monkeys | ||||||||||||||||||||||||||||||||||||
Coding sites in the Nav1. 4 gene 4 gene | Resistance to tetrodotoxin or saxotoxin | Snakes, pufferfish, clam | ||||||||||||||||||||||||||||||||||||
| Кодирующая область TAS2R38 | Чувствительность к горечи | Человек, шимпанзе | ||||||||||||||||||||||||||||||||||||
| Цис -регуляторные и кодирующие области в Агути | Пигментация кожи и шерсти | Человек, мышь-олень, крупный рогатый скот, свинья, собака, кошка, лошадь, лиса, домашняя лиса, перепел, овца в среде с низким содержанием глюкозы | Дрожжи | |||||||||||||||||||||||||||||||||||
| Количество дупликаций гена CCL3L1 | Восприимчивость к ВИЧ-инфекции и скорость прогрессирования СПИДа после инфицирования | Человек | ||||||||||||||||||||||||||||||||||||
| CIS -Регуляторный элемент в гене WNTA | Pigmentation Photels | Butterflies |
Открытые в Antice
1110 111111111111111111111111111111111111111111110 гг. случаи.
случаи.Фенотипическая часть взаимосвязи ГП
Фенотипическая часть взаимосвязи ГП относится к разновидности вариации (цвет волос, уровень устойчивости к токсинам и т. д.), а не к состоянию (светлые волосы, дегустация фенилтиокарбамида и т. д.). .; Таблица ).
Фенотип, связанный с генетическим изменением, не обязательно ограничивается организмом, содержащим генетическую мутацию. Например, различие между лево- и правозакрученными раковинами у улитки Lymnaea peregra определяется одним генетическим локусом с материнским эффектом: генотип матери, а не самой особи, отвечает за направление скручивание раковины (Boycott et al., 1931). В других случаях причинное генетическое изменение связано с бактериями-симбионтами: термостойкость тли может различаться у разных особей из-за точечной мутации в их бактериальном симбионте (Dunbar et al., 2007). Определенные фенотипические эффекты могут проявляться и на более высоком уровне, чем организм, несущий генетическое изменение (Dawkins, 19). 82), одним из показательных случаев является социальная организация муравьиной колонии (Wang et al., 2013).
82), одним из показательных случаев является социальная организация муравьиной колонии (Wang et al., 2013).
Дифференциальная часть родства GP
Как определено выше, родство GP охватывает генетическое различие и фенотипическое различие. Связь различия как на генетическом, так и на фенотипическом уровне довольно абстрактна и может соответствовать трем различным различиям в живом мире: (#1) различие между двумя репродуктивно изолированными таксонами (живущими или вымершими), (# 2) различие, сегрегирующее внутри популяции, и (3) различие, впервые появившееся в ходе эволюции, между организмом, имеющим наследственный аллель/признак, и его прямым потомком, развившим новый аллель/признак. Следует отметить, что изменчивость фенотипа не всегда сразу следует за появлением новой вызывающей мутации, но может появиться позже из-за единичного набора аллелей, сегрегирующих в популяции. Например, новый фенотип рецессивных панцирей появился в популяции пресноводной колюшки, когда рецессивный 9Аллель 0041 EDA , уже присутствующий на криптических уровнях, оказался в гомозиготном состоянии у одного человека (Colosimo et al. , 2005; Jones et al., 2012). Главный концептуальный прорыв, сделанный Чарльзом Дарвином, заключался в том, чтобы связать изменчивость среди особей внутри скрещивающейся группы (различие № 2) с изменчивостью между таксономическими группами в пространстве и времени (различие № 1; Левонтин, 1974а).
, 2005; Jones et al., 2012). Главный концептуальный прорыв, сделанный Чарльзом Дарвином, заключался в том, чтобы связать изменчивость среди особей внутри скрещивающейся группы (различие № 2) с изменчивостью между таксономическими группами в пространстве и времени (различие № 1; Левонтин, 1974а).
Отметим также, что определенные фенотипические изменения могут проявляться на уровне всего организма, когда «причинная» мутация сопровождается дополнительными соматическими мутациями, которые весьма вероятны. Например, у женщин, несущих аллель дикого типа и мутантный аллель BRCA1 , клетки могут продуцировать белки BRCA1 дикого типа, поскольку они несут одну копию аллеля BRCA1 дикого типа. Тем не менее, у этих женщин риск развития рака груди или яичников к 70 годам составляет до 80% по сравнению с женщинами, имеющими две копии дикого типа BRCA1 , из-за появления дополнительных вредных мутаций в диком типе BRCA1 . аллель в их соматических клетках молочной железы (Narod and Foulkes, 2004).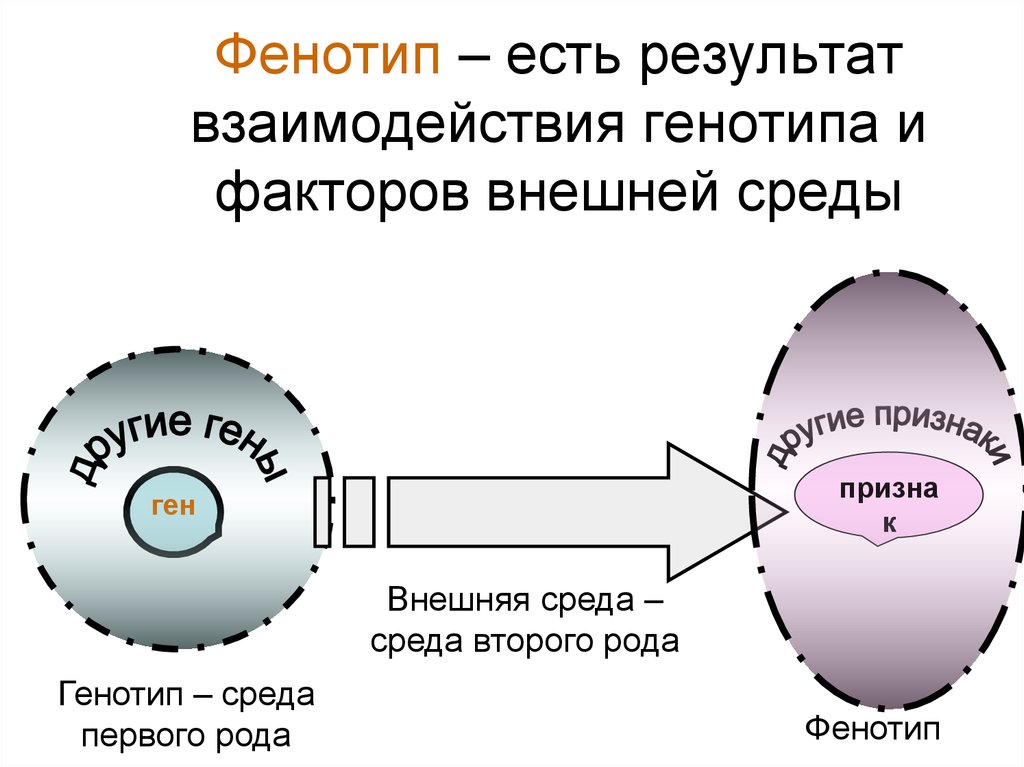
Важно отметить, что разница GP всегда определяется относительно интересующей популяции или таксона (Sober, 1988). В менее развитых с медицинской точки зрения странах люди, несущие две дефектные копии гена фенилаланингидроксилазы, имеют серьезные проблемы со здоровьем, включая судороги и умственную отсталость. Напротив, в большинстве развитых стран такие люди диагностируются при рождении и имеют нормальную продолжительность жизни с нормальным умственным развитием благодаря диете с ограничением фенилаланина (Armstrong and Tyler, 1955). Таким образом, взаимосвязь с GP, включающая мутацию с дефектом фенилаланингидроксилазы, зависит от контекста: мутация связана с проблемами со здоровьем в менее развитых в медицинском отношении странах, но не в других странах. Этот пример показывает, что причинно-следственная связь между генетическим изменением и связанным с ним фенотипическим изменением может скрывать множество встроенных параметров (таких как медицинская практика в случае фенилаланингидроксилазы) в пределах при прочих равных условиях допущение «при прочих равных условиях».
Таким образом, связь GP лучше всего рассматривать как связь между двумя вариациями, одна на уровне генотипа, а другая на уровне фенотипа. Человеческий разум может разрабатывать концепции возрастающей абстракции: концепции вещей (например, клетки), концепции изменения (например, эволюции) и концепции отношений (например, гомологии; Cassirer, 1910; Simondon, 1968). Здесь понятие отношения ВП устанавливает отношение между двумя меняет (генетические и фенотипические). В следующих абзацах мы покажем, что по сравнению с использованием интуитивных представлений о вещах этот обходной путь через усиление абстракции может оказаться более эффективным для лучшего понимания фенотипического разнообразия.
Выше мы утверждали, что при размышлениях о связи между генотипами и фенотипами всегда следует помнить о дифференциальном подходе. GWAS, представляющие собой наиболее популярный метод выявления геномных локусов, ассоциированных со сложными признаками в популяциях, основаны на анализе различий (Visscher et al. , 2012). Тем не менее, в современных исследованиях дифференцированная точка зрения иногда неявно отвергается. Когда наблюдается влияние нескольких факторов на фенотипические признаки ( Рисунок ), дифференциальная точка зрения считается слишком упрощенной, и исследователи часто предпочитают сосредотачиваться на фенотипах отдельных особей, не связывая их явно с фенотипическим эталоном.
, 2012). Тем не менее, в современных исследованиях дифференцированная точка зрения иногда неявно отвергается. Когда наблюдается влияние нескольких факторов на фенотипические признаки ( Рисунок ), дифференциальная точка зрения считается слишком упрощенной, и исследователи часто предпочитают сосредотачиваться на фенотипах отдельных особей, не связывая их явно с фенотипическим эталоном.
В большинстве современных статей проблема связи генотипа с фенотипом формулируется в терминах карт генотипа и фенотипа. Первая карта ВП была представлена Ричардом Левонтином в его книге «Генетическая основа эволюционных изменений» (Левонтин, 1974а; ).0030). Он указал средний генотип популяции как точку в пространстве всех возможных генотипов (пространство G) и средний фенотип той же популяции как соответствующую точку в пространстве всех возможных фенотипов (пространство P). Таким образом, эволюционный процесс был разбит на четыре этапа: (1) средний фенотип возникает в результате развития различных генотипов в различных условиях; (2) миграция, спаривание и естественный отбор действуют в пространстве P, чтобы изменить средний фенотип исходной популяции на средний фенотип особей, которые будут иметь потомство; (3) идентичность успешных родителей определяет, какие генотипы сохраняются; и (4) генетические процессы, такие как мутация и рекомбинация, изменяют положение в G-пространстве.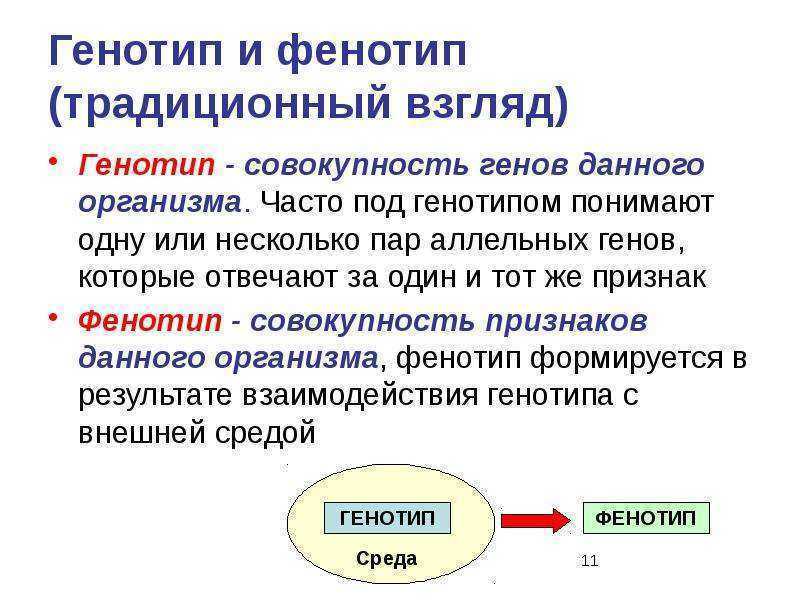
Открыть в отдельном окне
Три текущих графических представления карт GP. (A) Ранняя версия карты GP, предложенная Левонтином (1974а). (B) Карта GP, где каждая точка представляет одного человека (Houle et al., 2010; Gjuvsland et al., 2013; Salazar-Ciudad and Marín-Riera, 2013). (C) Отношения между признаками и генами, как показано Вагнером (1996). Подробности смотрите в тексте.
В другом распространенном графическом представлении ( Рисунок ), точка в пространстве G и соответствующая ей точка в пространстве P соответствуют генотипу и фенотипу отдельной особи (Fontana, 2002; Landry and Rifkin, 2012). При таком представлении абстрактный объект, который мы определили выше как отношение ГП, будет соответствовать «ходу» в пространстве генотипов, связанному с «ходом» в пространстве фенотипа (или, лучше, сумме нескольких «ходов» в пространстве генотипа, и пространства фенотипов, потому что несколько различных геномов могут нести два альтернативных аллеля данного родства GP).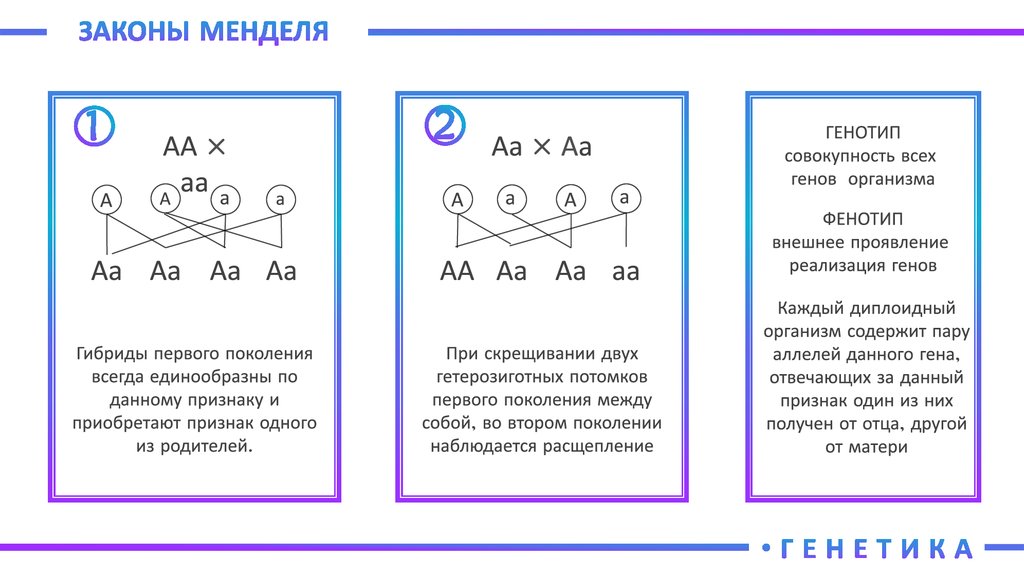 В третьем представлении, выдвинутом Вагнером (1996; Рисунок ), отдельные гены связаны с отдельными признаками.
В третьем представлении, выдвинутом Вагнером (1996; Рисунок ), отдельные гены связаны с отдельными признаками.
Хотя эти три графических представления карт ВП могут облегчить наше понимание некоторых аспектов биологии, во всех из них взаимосвязь ВП и дифференциальное представление понять нелегко. Вызывает недоумение тот факт, что первым человеком, нарисовавшим такую карту ВП, был Ричард Левонтин, красноречивый сторонник дифференциального взгляда (см., например, его предисловие к Oyama, 2000, шедевр убеждения). Поскольку эти графики сосредоточены на индивидуальных, а не на дифференциальных объектах, мы полагаем, что эти три репрезентации имплицитно побуждают нас вернуться к более интуитивной идее об одном генотипе, связанном с одним фенотипом. Упускание из виду дифференциального взгляда также может происходить с точки зрения молекулярной биологии, где белки рассматриваются как имеющие причинные эффекты сами по себе, такие как фосфорилирование субстрата или связывание с последовательностью ДНК. Из-за двух запутанных определений гена, либо как кодирующего белок, либо как вызывающего фенотипическое изменение (Griffiths and Stotz, 2013), легко перейти от дифференциального взгляда к недифференциальному взгляду на отношения GP.
Из-за двух запутанных определений гена, либо как кодирующего белок, либо как вызывающего фенотипическое изменение (Griffiths and Stotz, 2013), легко перейти от дифференциального взгляда к недифференциальному взгляду на отношения GP.
Таким образом, многие современные ментальные представления о связи между генотипом и фенотипом имплицитно отвергают дифференциальную точку зрения. Теперь мы покажем, что дифференциальная точка зрения совместима с тем фактом, что на фенотипические черты влияет сложная комбинация множества факторов, и что мы можем найти соответствующее схематическое представление взаимосвязей GP.
Разложение организма на элементарные единицы, такие как анатомические структуры, сыграло важную роль во многих биологических дисциплинах, таких как физиология, палеонтология и эволюция. Однако вопрос состоит в том, чтобы определить разложение на признаки, наиболее адекватное интересующему вопросу. Для вопросов, связанных с отношениями между органами различных особей или видов (таких как гомология), может быть уместно сохранить традиционную декомпозицию на анатомические структуры (Wagner, 2014). Рихард Левонтин и Гюнтер Вагнер определили характеры как элементы внутри организма, которые отвечают на адаптивные вызовы и представляют собой квазинезависимые единицы эволюционных изменений (Левонтин, 19).78; Вагнер, 2000). Их определение касается абсолютных признаков, наблюдаемых у отдельных организмов (например, форма крыла или количество пальцев у особи), и поэтому далеко от дифференциального взгляда. Здесь, чтобы лучше понять эволюцию и фенотипическое разнообразие живого мира, мы предлагаем разложить наблюдаемые признаки организма на множество элементарных вариаций ГП, которые накапливались в течение многих поколений, начиная с исходного состояния. Мы настаиваем на том, что с этой точки зрения персонажи — это не конкретные объекты (например, кожа), а абстрактные сущности, определяемые существованием различия между двумя возможными наблюдаемыми состояниями (например, цветом кожи). В качестве аналогии можно представить два способа изготовления изношенной кожаной обуви определенной формы. Можно либо собрать разные атомы в одну организацию, либо можно купить в магазине ботинок, а затем подвергнуть его ряду механических воздействий. Мы естественно склонны сравнивать организмы с машинами и мыслить в терминах частей, которые должны быть собраны, чтобы создать функциональное целое. Однако безудержной метафоры дизайнера или создателя недостаточно для понимания происхождения современных организмов (Coen, 2012). Чтобы понять фенотипические особенности данного организма, более эффективно разложить его на абстрактные изменения, которые происходили последовательно на протяжении эволюционного времени, а не на протяжении времени развития. Исходное состояние является гипотетическим предком изучаемого организма.
Наблюдается, что некоторые мутации (квалифицированные как плейотропные) поражают сразу несколько органов, в то время как другие изменяют только один за раз (Paaby, Rockman, 2013; Zhang, Wagner, 2013). Для плейотропных мутаций мы считаем, что связь ГП должна включать все фенотипические изменения (в разных органах, на разных стадиях и т. д.), связанные с генетическим различием. Например, мутация V370A рецептора EDAR связана не только с толщиной волос, но и с изменением плотности потовых и молочных желез в азиатских популяциях (Камберов и др., 2013). Отношения GP в таких случаях являются один-ко-многим. Рассмотрение кожи и глаза как независимых анатомических модулей человеческого тела может показаться уместным для многих эволюционных изменений, но несколько неадекватным в тех случаях, когда эти два органа одновременно развили новый признак пигментации в результате единственной мутации в Ген SLC45A2 (Liu et al., 2013). Рассуждения в терминах взаимосвязей ГП снимают проблему нахождения соответствующей декомпозиции на элементарные анатомические структуры. Сами элементарные отношения ГП представляются адекватными полунезависимыми модулями, комбинация которых может объяснить наблюдаемые характеристики организма.
В рамках дифференциальной концепции взаимоотношений ГП одним из важнейших моментов является разложение наблюдаемых признаков на ряд полунезависимых фенотипических вариаций, то есть выявление элементарных изменений, произошедших в ходе эволюции. Доступны экспериментальные подходы для разложения данного фенотипического различия на соответствующие более тонкие подвариации. Например, скрещивание растений с разной формой листьев дает потомство, которое демонстрирует составной набор промежуточных форм листьев. Анализ основных компонентов выявил элементарные изменения формы листа, которые вместе могут объяснить разницу в форме между родительскими линиями и, по-видимому, вызваны разными геномными областями (Langlade et al., 2005). Это наводит на мысль, что «сумма затемняет части». То, что мы традиционно считаем сложными признаками, может быть составлено из более простых признаков, более поддающихся генетическому анализу. Другим ярким примером является пигментация живота в Группа Drosophila dunni . Взятые как одна переменная, уровни пигментации демонстрируют сложную генетическую архитектуру, но разложение взрослых паттернов на анатомические субъединицы раскрывает дискретный генетический контроль для каждого субпризнака (Hollocher et al. , 2000). Более известным случаем является эволюция цвета тела пляжных мышей. Различие в окраске между светлыми пляжными мышами и темными мышами можно разложить на отдельные фенотипы (оттенок спины, яркость спины, ширина полосы на хвосте и дорсовентральная граница), которые все связаны с различными мутациями в Ген Agouti (Linnen et al., 2013; Рисунок ). Каждый генетический локус Agouti , по-видимому, предназначен для спецификации пигментации в данной части тела. Вместе они образуют группу тесно связанных локусов, связанных с изменениями пигментации шерсти.
Открыть в отдельном окне
Эволюция светлоокрашенных пляжных мышей обусловлена несколькими мутациями с выраженными эффектами пигментации в локусе Agouti . Темный и светлый фенотипы можно разложить на четыре фенотипических признака, которые связаны с различными однонуклеотидными полиморфизмами (SNP, цветные точки), расположенными в Агути ген. Показаны только SNP с предполагаемым коэффициентом отбора светлого аллеля выше 0,1. Кодирующие экзоны представлены темными прямоугольниками, а нетранслируемые экзоны — белыми прямоугольниками. Адаптировано из Linnen et al. (2013).
Хотя сложные признаки не всегда можно свести к набору простых взаимосвязей GP, возможно, что такие признаки, как рост взрослого человека, наиболее характерный количественный признак, который, по прогнозам, состоит из множества генетических эффектов небольшого размера (Fisher, 1930), также могут быть разложены на элементарные вариации, каждая из которых объясняет более дискретные субпризнаки. В то время как некоторые детерминанты человеческого роста, такие как LIN28B были связаны с ростом взрослого человека в разном возрасте, другие гены достигли статистической значимости только в стадийных исследованиях, посвященных росту плода и скорости роста в период полового созревания (Lettre, 2011). Другими словами, эти данные предполагают, что рост человека может быть составным признаком, который модулируется несколькими отношениями GP, каждое из которых действует на разных фазах развития.
Взаимодействие генов с окружающей средой (GxE) происходит, когда фенотипический эффект данного генетического изменения зависит от параметров окружающей среды. Сходным образом, эпистаз или взаимодействие GxG возникает, когда фенотипический эффект данного генетического изменения зависит от аллельного состояния по крайней мере одного другого локуса (Phillips, 2008; Hansen, 2013). Появляется все больше свидетельств того, что взаимодействия GxG и GxE имеют фундаментальное значение для понимания эволюции и наследования сложных признаков (Gilbert and Epel, 2009).; Хансен, 2013). Мы предполагаем, что оба явления могут быть интегрированы в базовую дифференциальную структуру GP, где взаимодействия GxG и GxE вводят уровень контекстной зависимости и приводят к различиям, встроенным в различия.
Разница в цветовой пигментации между темными и светлыми пляжными мышами, упомянутая ранее ( Рисунок ), обусловлена не только мутациями в Agouti , но и кодирующей мутацией в гене MC1R , которая уменьшает пигментацию (Steiner et и др. , 2007; Рисунок ). Эффект мутации MC1R виден только в присутствии ассоциированного со светлым цветом производного гаплотипа Agouti . Здесь считается, что локус Mc1R эпистатически взаимодействует с локусом Agouti . В этом случае мы предполагаем, что отношение GP включает не одно фенотипическое различие, а два возможных фенотипических различия (изменение пигментации шерсти или полное отсутствие изменений). Выбор между этими двумя фенотипическими различиями определяется генетическим фоном (здесь на Агути, местонахождение ). Дифференциальный взгляд, таким образом, остается относительно простым для взаимодействия двух локусов: зависимость фенотипа от контекста транслируется в выбор между двумя возможными фенотипическими различиями. Мы предполагаем, что отношения GP, включающие мутацию, подверженную множественным эпистатическим взаимодействиям, должны включать все возможные фенотипические различия, которые могут возникнуть в результате мутации во всех генетических фонах. Среди всех возможных фенотипических вариаций фенотипическое различие, которое будет наблюдаться, определяется другими генетическими локусами. В общем, взаимодействия GxG включают несколько сайтов, разбросанных по всему геному (Bloom et al., 2013).
Открыть в отдельном окне
Взаимодействие генов с окружающей средой (GxE) и GxG. (A) Мутация, кодирующая npr-1 , влияет на агрегационное поведение нематод при уровне кислорода 21%, но не при 10% (Andersen et al., 2014). (B) Мутация Mc1R, кодирующая , влияет на пигментацию тела мыши в присутствии доминантных светлых аллелей Agouti , но не в гомозиготном фоне Agouti по рецессивному темному аллелю (Steiner et al., 2007).
Примером взаимодействия GxE (см. также Рисунок ) является природный аллель потери функции brx в растениях Arabidopsis , который связан с ускоренным ростом и повышенной приспособленностью к кислым почвам, а также к сильно замедленный рост корней по сравнению с диким типом в нормальных почвах (Gujas et al. , 2012). Взаимодействия GxE обычно анализируются в форме нормы реакции , которая представляет все наблюдаемые черты с одним генотипом в различных средах (Johannsen, 1911; Sarkar, 1999). В случае взаимодействий GxE мы предполагаем, что отношения GP должны включать все возможные фенотипические изменения , которые могут быть вызваны ассоциированными генетическими изменениями в различных экспериментальных условиях. Таким образом, связанное с этим фенотипическое изменение представляет собой разницу между двумя нормами реакции. Примером из учебника является изменение правила размера и температуры у C. elegans . Как и большинство других животных, 9Нематоды 0041 C. elegans увеличиваются в размерах при низкой температуре, но лабораторный штамм дикого типа C. elegans , происходящий с Гавайских островов, не показывает изменений в размере тела при различных температурах. Аминокислотное изменение в кальций-связывающем белке ответственно за снижение способности гавайского штамма увеличиваться в размерах при низкой температуре (Kammenga et al. , 2007). Здесь норма реакции (представляющая размер тела нематоды в диапазоне температур) различается между нематодами, и связанное с ней отношение GP охватывает разницу между этими двумя наклонами.
Диапазон фенотипических вариаций, воплощенных в отношениях GP, подверженных взаимодействиям GxG и GxE, может быть весьма огромным, особенно в случаях, когда одна и та же мутация затрагивает несколько тканей, и когда на фенотипическую изменчивость каждой ткани влияют другие геномные локусы и по условиям окружающей среды. Фактически, фенотипические эффекты мутации всегда зависят от других фрагментов ДНК из того же генома, так что можно считать, что любые отношения GP испытывают эпистаз. Другими словами, генетический локус, влияющий на фенотип, никогда не действует независимо от других последовательностей ДНК. Например, данный аллель опсина будет приводить к определенным свойствам цветового зрения только в том случае, если глаз сформирован и если этот глаз получает свет во время своего развития, что позволяет сформировать эффективные зрительные нейронные цепи. Для того чтобы дифференцированный взгляд был приемлемым, мы советуем не рассматривать все возможные генетические предпосылки и условия окружающей среды, а ограничивать возможности потенциальной средой и выделять аллели, которые имеют отношение к интересующей популяции (Sober, 19).88).
Таким образом, при наличии эпистаза или взаимодействия GxE генетическое изменение связано не с одним фенотипическим отличием, а с несколькими возможными фенотипическими различиями, среди которых будет достигнуто одно, в зависимости от окружающей среды и генетического фона. Зависимость от контекста может быть схематически представлена как различия ГП, встроенные в другие различия генотипа и среды.
Дифференциальный взгляд на генетические и экологические эффекты на фенотипы
Как подчеркивают многие авторы (прежде всего Waddington, 1957; Oyama, 2000; Keller, 2010), гены и среда действуют на фенотип совместно, и в большинстве случаев невозможно отделить влияние одного от другого. Здесь мы показываем, что рассуждения с точки зрения различий помогают прояснить сравнение между генетическими и экологическими эффектами на фенотипы. Однако мы выделяем некоторые случаи, когда сравнение остается затруднительным.
По аналогии с отношением GP мы можем определить отношение среда-фенотип как вариацию среды (две среды), связанное с ней фенотипическое изменение (различные фенотипические состояния) и их взаимосвязь. Например, у многих видов черепах изменение температуры во время развития яиц связано с половыми различиями между самцами и самками (9).0029 Рисунок
) и по крайней мере шесть переходов от определения пола в окружающей среде к генетическому ( Рисунок ) произошли в филогенезе черепах (Pokorná and Kratochvíl, 2009). В этом случае средовые и генетические эффекты можно сопоставить: половые хромосомы и температура оказывают одинаковое фенотипическое влияние на черепах. Такие наблюдения привели Уэст-Эберхард (West-Eberhard, 2003, 2005) к предложению гипотезы «гены как последователи», которая предполагает, что новые фенотипические состояния с большей вероятностью возникают в первую очередь из-за изменения окружающей среды, чем из-за генетической мутации, и что мутации происходят только позже, при изменении порога экспрессии нового признака. Уэст-Эберхард (2003, 2005) экстраполировал различия, сегрегирующие внутри популяций (различие № 2), на различия, возникшие во времени во время эволюции популяции (различие № 1).Открыть в отдельном окне
Связь между средой и фенотипом по сравнению с зависимостью от GP для определения пола у черепах. (A) У некоторых видов температура во время эмбрионального развития определяет пол взрослой особи. (B) У других пол определяется половыми хромосомами.
Независимая эволюция направленной лево-правой асимметрии от симметричных предков во многих линиях предоставила основной аргумент в поддержку гипотезы «ген как последователь» (Palmer, 2004). В рамках этой схемы оговаривается, что направленная асимметрия, при которой все люди односторонние, часто развилась из состояния «случайной асимметрии», когда направленность будет зависеть от факторов окружающей среды и, таким образом, различаться между генетически идентичными людьми. Например, самая сильная клешня омара будет развиваться в зависимости от использования и имеет априори равные вероятности развития с левой или с правой стороны. Мы можем видеть, как здесь работает формула «гены как последователи»: окружающая среда запускает асимметрию, и позже в ходе эволюции некоторые генетические эффекты могут изменить ее направленность в ту или иную сторону. Но в то время как асимметрия «происходит до того, как существует генетическая изменчивость, чтобы контролировать ее», дифференциальная точка зрения ясно дает понять, что генетическое влияние на направленность несопоставимо с влиянием окружающей среды, которое запускает асимметрию. Генетическое изменение приводит к переключению между окончательным 100% односторонним состоянием и начальным состоянием, при котором 50% случаев являются правосторонними и 50% левосторонними. Напротив, два альтернативных фенотипических состояния, возникающие в результате изменчивости в окружающей среде, считаются 100% правосторонним и 100% левосторонним. Этот пример показывает, что ради точности важно явно указать различия, которые учитываются в отношениях GP.
Дифференциальное представление обеспечивает теоретическую основу, которая может помочь в разработке экспериментов для исследования соответствующих переменных: можно сравнивать различные генотипы в фиксированной среде (классическая взаимосвязь GP), сравнивать реакцию фиксированного генотипа на две разные среды (фенотипическая пластичность). ), или сравнить чувствительность двух разных генотипов к двум разным средам (где фенотипическая вариация становится различием в различии; см., например, Engelman et al., 2009).; Томас, 2010).
Были разработаны различные количественные методы, чтобы отделить генетику от эффектов окружающей среды и количественно оценить взаимодействия GxE (Lynch and Walsh, 1998). Тем не менее, в определенных ситуациях может оказаться невозможным биологически значимым образом отделить генетические эффекты от воздействий окружающей среды, даже если рассуждать в терминах различий (Lewontin, 1974b). Популяции жука Calathus melanocephalus представлены двумя морфами: длиннокрылой и короткокрылой (Schwander, Leimar, 2011). Длиннокрылая морфа развивается только из гомозиготных особей по рецессивному аллелю, сегрегирующему в популяции, и только при хороших пищевых условиях. В этом случае генетические и средовые эффекты смешиваются (9).0029 Фигуры ). В теоретическом случае популяции, состоящей только из короткокрылых гетерозиготных животных, выращенных в условиях голодания, и длиннокрылых, за разницу крыльев между особями ответственны как гены, так и среда, и невозможно оценить соотношение факторов среды и среды. генетические эффекты, поскольку гены и окружающая среда действуют на различных уровнях сложной причинно-следственной связи между генотипами и фенотипами.
Открыть в отдельном окне
Перспективы взаимосвязи среда-фенотип и отношения GP для полиморфизма длины крыла у жука Calathus melanocephalus . (A) С точки зрения взаимосвязи среда-фенотип изменение условий питания связано с изменением размера крыльев, но только в гомозиготном фоне по рецессивному аллелю ( l ) локуса размера крыльев. (B) С точки зрения взаимосвязи GP генетическое изменение в локусе размера крыла связано с изменением размера крыла, но только при хороших условиях питания.
Другой случай, который ставит под сомнение классическое различие между окружающей средой и генетикой, — это когда добавление определенных симбиотических бактерий изменяет фенотип хозяина. Мыши, которых кормили штаммом бактерий Lactobacillus , демонстрируют меньшее поведение, связанное с тревогой, по сравнению с контрольными мышами, которых кормили бульоном без бактерий (Bravo et al. , 2011). Здесь разница в поведении вызвана переключением между присутствием или отсутствием определенного кишечного симбионта. Причиной фенотипических различий является не простое изменение в последовательности ДНК, не простое изменение окружающей среды, не связанное с генетическими изменениями, а переключение между присутствием и отсутствием фактора, который можно рассматривать как фактор окружающей среды, — бактерий, — который содержит ДНК, мутации которой могут также изменить фенотип хозяина.
В заключение, рассуждения с точки зрения различий могут помочь прояснить сравнение между генетическими и экологическими эффектами на фенотипы. Тем не менее, вопросы не более чем простые. Поскольку гены и среда действуют на разных уровнях сложной причинной связи между генотипами и фенотипами, в некоторых случаях невозможно распутать обе причины.
Фенотипические различия, по-видимому, подпадают под две основные категории: либо наличие/отсутствие чего-либо (например, волосы на теле или способность переваривать молоко), либо сдвиг между двумя альтернативами, которые присутствуют одновременно (например, два цвета волос). Точно так же со стороны генотипа мутация может соответствовать присутствию/отсутствию соответствующей последовательности ДНК или полиморфизму нуклеотидов. Дифференциальная точка зрения делает очевидным, что потеря фенотипа не обязательно связана с потерей генетического материала, и наоборот. Например, эволюционное приобретение темных пигментов, покрывающих всю шерсть животных, часто связывают с утратой Ген Mc1R (Gompel, Prud’homme, 2009). Кроме того, как ранее отмечал один из нас (приложение Stern and Orgogozo, 2008), усиление или потеря фенотипа субъективны. Например, выпадение волос также можно рассматривать как приобретение обнаженного эпидермиса. Большинство эпидермальных клеток насекомых дифференцируются в одно из этих двух альтернативных состояний, и в обоих состояниях участвуют большие сети регуляции генов. Неясно, какое фенотипическое состояние представляет собой выигрыш или потерю по сравнению с другим. Даже с точки зрения генотипа определение потерь и приобретений может быть затруднено. Вставка мобильного элемента может сбить ген, тогда как делеция иногда может создать новый сайт связывания для активатора транскрипции. На самом деле, эволюционный выигрыш 9Экспрессия 0041 desatF в D. melanogaster произошла посредством серии из трех делеций, каждая из которых создавала гексамерный мотив, необходимый для экспрессии desatF (Shirangi et al., 2009).
Точно так же дифференциальная точка зрения на воздействие окружающей среды выдвигает на первый план ошибочность различия между разрешительными и инструктивными сигналами. Разрешающий сигнал связан с наличием/отсутствием фенотипа, а инструктивный сигнал — со сдвигом между двумя присутствующими альтернативами. Как утверждалось выше, эти различия на фенотипическом уровне не являются четкими.
В заключение мы предлагаем с осторожностью использовать термины «прибыль/убыток» и «поучительная/разрешительная» терминология.
Ожидается, что мутация вызовет в некоторой степени воспроизводимую фенотипическую изменчивость в популяции. Такая воспроизводимость фенотипического исхода необходима для обеспечения генетической эволюции и адаптации путем естественного отбора (Lewontin, 1974a; Kirschner and Gerhart, 1998). В самом деле, вновь образованный аллель, который будет генерировать еще один фенотип каждый раз, когда он попадает в другой организм, не будет подвергаться естественному отбору. Рассуждение с точки зрения изменчивости, а не рассмотрение аллелей как изолированных объектов, проясняет, что конкуренция происходит между аллелями, которые охватывают один и тот же генетический локус. Естественный отбор воздействует непосредственно на аллельную вариацию, которая устойчиво связана с данной фенотипической вариацией, что и является родством ГП. Таким образом, отношения ГП являются базовой единицей эволюционных изменений, на которые действует естественный отбор.
Главное открытие последних 20 лет состоит в том, что изменчивость в определенных генетических локусах приводит к сопоставимой фенотипической изменчивости не только у разных особей одной популяции, но и у чрезвычайно разных таксонов (Martin and Orgogozo, 2013b). Другими словами, определенные отношения GP являются таксономически надежными и присутствуют у большого количества видов. Это означает, что генетические и экологические предпосылки оставались относительно постоянными или неоднократно появлялись на протяжении эволюции, что позволяло генетическим локусам генерировать сходные фенотипические изменения в различных таксономических группах. Это важное открытие было совершенно неожиданным около 50 лет назад. Долгое время ожидалось, что сингулярность, наблюдаемая в живом мире, будет отражать сравнимую сингулярность на генетическом уровне, затрагивая несопоставимые и неконсервативные гены, специфичные для каждой линии (Mayr, 19).63). Как однажды предположил Майр (1963) в 1963 году: «Многое из того, что было известно о физиологии генов, делает очевидным, что поиск гомологичных генов совершенно бесполезен, за исключением очень близких родственников […]. Поговорка «Многие дороги ведут в Рим» справедлива как в эволюции, так и в повседневных делах» (Mayr, 1963). Другими словами, ожидалось, что генетические локусы, которые делают человека мужчиной, будут отличаться от тех, которые делают собаку или рыбу. Позже, в 80–90-х годах, некоторые исследователи предположили, что эволюция происходит за счет мутаций в консервативных генах, кодирующих белок (Romero-Herrera et al., 19).78; Перуц, 1983; Стюарт и др., 1987; Carroll et al., 2005), но у них было мало экспериментальных данных, подтверждающих их точку зрения (Tautz and Schmid, 1998). Накопившиеся на сегодняшний день данные о мутациях, ответственных за естественную изменчивость, ясно показывают, что разнообразие живых организмов имеет общую генетическую основу как минимум в трех пунктах. Во-первых, сравнительная биология развития показала, что животные имеют общие наборы ключевых регуляторных генов с законсервированными функциями (Wilkins, 2002, 2014; Carroll et al., 2005). Во-вторых, большинство межвидовых различий у животных и растений, для которых хотя бы частично идентифицирована лежащая в основе генетическая основа (154 случая из 160), обусловлено мутациями в гомологичных генах, и очень немногие (6/160) обусловлены новыми генами. которые, тем не менее, представляют собой дубликаты существующих генов (Martin and Orgogozo, 2013b). В-третьих, было показано, что множественные случаи сходных фенотипических изменений связаны с мутациями одних и тех же гомологичных генов в независимых линиях (9).0029 Таблица ), иногда на больших филогенетических дистанциях. Например, разница в пигментации между белыми и оранжевыми бенгальскими тиграми недавно была сопоставлена с единственной мутацией в гене белка-транспортера SLC45A2 (Xu et al., 2013), и этот ген также был связан с гипопигментацией глаз, кожи , волосы и перья у людей и кур (Xu et al., 2013; Figure ). Более ярким примером является недавняя эволюция устойчивости к токсинам у трех видов, которые разошлись более 500 миллионов лет назад – моллюска, змеи и иглобрюха – посредством замены одной и той же аминокислоты в законсервированном гене (Bricelj et al., 2005). ; Geffeney et al., 2005; Venkatesh et al., 2005; Feldman et al., 2012). Такие поразительные паттерны генетического повторения в настоящее время обнаружены для более чем 100 генов животных и растений (Martin and Orgogozo, 2013b). Несмотря на существующие методологические предубеждения в пользу консервативных генов при поиске локусов количественных признаков (Rockman, 2012; Martin and Orgogozo, 2013b), уровень генетического повторения остается поразительным и предполагает, что для эволюции по крайней мере некоторых фенотипических различий требуется относительно мало генетических путей. ведут в Рим (Stern, 2013). В наши дни не следует удивляться тому, что фрагмент ДНК, связанный со сложной окраской крыльев в одном Вид бабочек Heliconius обеспечивает сходные крылья и коллективную защиту от одних и тех же хищников при введении в геном других бабочек (Supple et al., 2014). То, что делает собаку собакой или человека человеком, теперь частично объясняется уникальным набором таксономически устойчивых отношений ГП, которые обнаруживаются во многих ветвях линии.
Определенные связи между средой и фенотипом также таксономически устойчивы. Например, у большинства таксонов на размер тела влияет питание; Дефицит железа может вызвать анемию, а некоторые токсичные соединения могут привести к летальному исходу. У экзотермов температура организма зависит от температуры окружающей среды. Учитывая устрашающее количество условий окружающей среды, которые можно представить, вероятно, невозможно определить, преобладают ли таксономически устойчивые отношения GP или таксономически устойчивые отношения среда-фенотип. Кроме того, вопрос о том, представляют ли таксономически устойчивые отношения GP исключительную и небольшую долю или значительную долю всех отношений GP, является предметом споров. В любом случае, существование таксономически устойчивых взаимосвязей GP теперь ясно и должно быть широко признано исследовательским сообществом.
Некоторые из самых поразительных учений современной биологии включают в себя открытие того, что живые существа имеют один и тот же генетический материал (ДНК или РНК), один и тот же генетический код (за некоторыми исключениями) и один и тот же базовый клеточный механизм. Таким образом, далеко не парадоксально, что индивидуальные различия строятся на сходствах, и тот факт, что определенные отношения ГП сохраняются на протяжении длительного времени эволюции, завершает картину.
Точная предсказательная сила, возникающая в результате существования таксономически надежных взаимосвязей GP, редко встречается в биологии и только начинает использовать свой полный потенциал. Сохранение связей GP на большие расстояния теперь полностью оправдывает использование подходов сравнительной генетики для решения прагматических проблем. Например, одомашнивание сельскохозяйственных культур приняло форму аналогичного давления отбора у многих видов, и теперь у нас есть экспериментальные доказательства того, что этот процесс неоднократно включал мутации в одном и том же наборе консервативных генов (Peterson et al., 19).95; Мартин и Оргогозо, 2013b). Это наблюдение открывает интересные возможности для применения, поскольку мы можем использовать эту новую совокупность генетических знаний, чтобы помочь одомашниванию будущих культур или использовать стратегии с помощью маркеров для производства и поддержания биоразнообразия сельскохозяйственных культур (Lenser and Theißen, 2013). Предсказуемость GP уже используется для идентификации штаммов, которые развили устойчивость к различным стратегиям борьбы с вредителями, причем крайние случаи нацелены на толерантность к противомалярийным препаратам у Plasmodium 9.0042 паразитов (Manske et al., 2012), устойчивость бактерий и дрожжей к антибиотикам (Fischbach, 2009; MacCallum et al., 2010) или, что еще более драматично, антропогенная эволюция устойчивости к инсектицидам у различных когорт насекомых, независимо от их статус вредителя (Ffrench-Constant et al., 2004; Martin and Orgogozo, 2013b).
Кроме того, повторяемость генетической основы фенотипической изменчивости предполагает, что клинические исследования, вероятно, также получат пользу от генетических исследований большого числа модельных видов (Robinson and Webber, 2014). Например, естественная изменчивость толерантности к метотрексату, химиотерапевтическому препарату, была картирована в 9 случаях.0041 Drosophila плодовых мух к генам, человеческие ортологи которых также связаны с реакцией пациентов на этот препарат (Kislukhin et al. , 2013), что расширяет возможности использования модельных организмов в качестве моделей заболеваний.
Один из оригинальных аспектов описания связи GP с точки зрения индивидуальных отношений GP заключается в том, что это позволяет классифицировать фенотипы в соответствии с лежащей в их основе генетической основой. На первом уровне отношения GP, затрагивающие разные регионы одного и того же гена и дающие сопоставимые фенотипические результаты, могут быть сгруппированы вместе. Простые случаи взаимодействия GxG были обнаружены между тесно связанными мутациями, как правило, внутри кодирующей последовательности или внутри цис- регуляторный элемент, когда они вызывают неаддитивный эффект на фенотип. Например, было обнаружено, что определенная мутация в энхансере вызывает различные сдвиги в паттерне экспрессии нижестоящего кодирующего гена в зависимости от соседней последовательности ДНК (Frankel et al., 2011; Rogers et al., 2013). Сходным образом было обнаружено, что аминокислотные мутации в гене гемоглобина увеличивают или уменьшают сродство к кислороду в зависимости от аллельного состояния других сайтов (Natarajan et al. , 2013). В таких случаях интуитивно понятно сгруппировать такие генетически связанные сайты вместе, поскольку все они влияют на один и тот же тип фенотипического признака.
Отсутствие пигментов меланина у животных было связано с мутациями в нескольких генах, включая OCA2 , kit-лиганд или Mc1R (обзор в Gompel and Prud’homme, 2009; Liu et al., 2013). В то время как отсутствие меланина традиционно рассматривается как одно из состояний признака, альбинизм, независимо от лежащей в основе генетической основы, мы предлагаем здесь различать OCA2 -ассоциированный альбинизм от Mc1R -ассоциированного альбинизма или альбинизма, связанного с любым другим геном. Одним из преимуществ разложения изменчивости живого мира на эти множественные элементарные отношения ЗП является то, что эти элементы затем могут быть сгруппированы в последовательно увеличивающиеся группы. Элементарные фенотипические изменения, затрагивающие разные гены, которые являются частью одного и того же генетического пути, также могут быть сгруппированы вместе как сопутствующие компоненты одного и того же механизма модуляции фенотипа. Это явно относится к сигнальным молекулам TGF-β BMP15, GDF9.и рецептор TGF-β BMPR1B, которые неоднократно связывали с изменениями функции яичников у людей и домашних овец (обзор в Luong et al., 2011).
Другим важным следствием перспективы родства GP является то, что очевидно различные фенотипические изменения, вызванные сходными генетическими локусами в различных организмах, могут быть исследованы далее, чтобы выявить то, что может быть общим основным фенотипическим изменением (Deans et al., 2015). Например, личинки мух и черви-нематоды имеют разное поведение при поиске пищи, но имеют мутации в одном и том же ортологическом гене (9).0041 для / egl-4 ) изменяют интенсивность поведения в поисках пищи у обоих организмов (Osborne et al., 1997; Mery et al., 2007; Hong et al., 2008). Таким образом, вполне вероятно, что основное поведенческое изменение, которое лежит в основе кажущихся отличительными изменений поиска пищи мухами и нематодами, представляет собой консервативное отношение GP между нематодами и мухами. Этот несколько пограничный пример иллюстрирует проблему включения широко распространенного сравнительного мышления в наше глобальное понимание биологии. Имеет ли отношение мутация в мышиной модели к заболеванию человека? Можем ли мы считать, что фенотип мыши подобен состоянию человека, если его генетическая основа иная? Мы и другие прогнозируем, что поиск ортологичных фенотипов, или «фенологов» (McGary et al., 2010), станет главной задачей современной генетики и потребует плодотворного союза между прикладной и эволюционной биологией.
В этой статье мы возвращаем дифференциальную концепцию гена (Schwartz, 2000) в нашу структуру для понимания карты GP. Дифференциальный взгляд на отношения GP помогает прояснить генетические и экологические эффекты на фенотипы и их связь. Это также открывает новые возможности для размышлений, в частности, в отношении декомпозиции наблюдаемых признаков внутри организма и представления карт GP. Кроме того, существование таксономически надежных взаимосвязей GP поощряет беззастенчивое использование сравнительной генетики для предсказания генетической основы фенотипической изменчивости в различных группах организмов, и эта предсказательная сила имеет важный потенциал для трансляционных исследований в области агрономии и клинических исследований.
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Мы глубоко признательны Мари-Анн Феликс за содержательные обсуждения и Томасу Прадеу за то, что он привлек наше внимание к нескольким важным документам. Мы также благодарим Giuseppe Baldacci, Marie-Anne Félix, Pierre-Henri Gouyon, Alexandre Peluffo, Mark Siegal, David Stern и рецензентов за их проницательные комментарии к рукописи. Исследование, которое привело к написанию этой статьи, финансировалось Европейским исследовательским советом в рамках Седьмой рамочной программы Европейского сообщества (FP7/2007-2013 Грантовое соглашение № 337579).) и от Фонда Джона Темплтона (грант № 43903).
| ГП | генотип-фенотип. |
- Андерсен Э. К., Блум Дж. С., Герке Дж. П., Кругляк Л. (2014). Вариант нейропептидного рецептора npr-1 является основной детерминантой роста и физиологии Caenorhabditis elegans . Генетика PLoS. 10:e1004156
10.1371/journal.pgen.1004156 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Армстронг М. Д., Тайлер Ф. Х. (1955). Исследования на фенилкетонурию. I. Ограничение приема фенилаланина при фенилкетонурии. Дж. Клин. Инвестировать. 34 565–580. 10.1172/JCI103105 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Bloom J. S., Ehrenreich I. M., Loo W. T., Lite T.-L. В., Кругляк Л. (2013). Поиск источников отсутствующей наследственности при скрещивании дрожжей. Природа 494 234–237. 10.1038/nature11867 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Бойкот А. Е., Дайвер К., Гарстанг С. Л., Тернер Ф. М. (1931). Наследование левосторонней формы у limnæa peregra (Mollusca, Pulmonata). Филос. Транс. Р. Соц. Лонд. сер. Б 219 51–131. 10.1098/rstb.1931.0002 [CrossRef] [Google Scholar]
- Браво Дж. А., Форсайт П., Чу М. В., Эскаравэдж Э., Савиньяк Х. М. , Динан Т. Г. и др. (2011). Прием внутрь штамма Lactobacillus регулирует эмоциональное поведение и экспрессию центрального рецептора ГАМК у мышей через блуждающий нерв. Проц. Натл. акад. науч. США 108
16050–16055. 10.1073/pnas.1102999108 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Bricelj V.M., Connell L., Konoki K., Macquarrie S.P., Scheuer T., Catterall W.A., et al. (2005). Мутация натриевого канала, приводящая к устойчивости моллюсков к сакситоксину, увеличивает риск ПНП. Природа 434 763–767. 10.1038/nature03415 [PubMed] [CrossRef] [Google Scholar]
- Кэрролл С. Б., Гренье Дж., Уэтерби С. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных . Молден, Массачусетс: John Wiley & Sons. [Google Scholar]
- Кассирер Э. (1910). Вещество и функция 2010 Эдн Париж: Набу Пресс. [Google Scholar]
- Коэн Э. (2012). От клеток к цивилизациям: принципы изменения, формирующие жизнь . Принстон, Нью-Джерси: Издательство Принстонского университета;
10.1515/9781400841653 [CrossRef] [Google Scholar]
- Colosimo P.F., Hosemann K.E., Balabhadra S., Villarreal G., Jr., Dickson M., Grimwood J., et al. (2005). Широко распространенная параллельная эволюция у колюшек путем повторной фиксации аллелей эктодисплазина. Наука 307 1928–1933 гг. 10.1126/science.1107239 [PubMed] [CrossRef] [Google Scholar]
- Докинз Р. (1982). Расширенный фенотип: длинная досягаемость гена . Оксфорд: Издательство Оксфордского университета. [Google Scholar]
- Динс А. Р., Льюис С. Э., Хуала Э., Анзальдо С. С., Эшбернер М., Балхофф Дж. П. и др. (2015). Находим свой путь через фенотипы. PLoS Биол. 13:e1002033 10.1371/journal.pbio.1002033 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Данбар Х.Е., Уилсон А.К.С., Фергюсон Н.Р., Моран Н.А. (2007). Термическая устойчивость тли определяется точечной мутацией в бактериальных симбионтах. PLoS Биол. 5:e96
10.1371/journal.pbio.0050096 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Engelman C.D., Baurley J.W., Chiu Y.-F., Joubert B.R., Lewinger J.P., Maenner M.J., et al. (2009). Обнаружение взаимодействий генов и окружающей среды в данных полногеномной ассоциации. Жен. Эпидемиол. 33(Приложение 1) С68–С73. 10.1002/gepi.20475 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Фальк Р. (1984). Ген в поисках идентичности. Гул. Жене. 68 195–204. 10.1007/BF00418388 [PubMed] [CrossRef] [Google Scholar]
- Feldman C.R., Brodie E.D., Jr., Brodie E.D., III, Pfrender M.E. (2012). Ограничение формирует конвергенцию в устойчивых к тетродотоксину натриевых каналах змей. Проц. Натл. акад. науч. США 109 4556–4561. 10.1073/pnas.1113468109 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Ffrench-Constant R.H., Daborn P.J., Le Goff G. (2004). Генетика и геномика устойчивости к инсектицидам. Тенденции Жене. 20
163–170. 10.1016/j.tig.2004.01.003 [PubMed] [CrossRef] [Google Scholar]
- Fischbach MA (2009). Антибиотики от микробов: сходятся, чтобы убить. Курс. мнение микробиол. 12 520–527. 10.1016/j.mib.2009.07.002 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Fisher RA (1930). Генетическая теория естественного отбора . Оксфорд: Кларендон; 10.5962/bhl.title.27468 [CrossRef] [Google Scholar]
- Фонтана В. (2002). Моделирование «эво-дево» с помощью РНК. Новости BioEssays Rev. Mol. Клетка. Дев. биол. 24 1164–1177 гг. 10.1002/bies.10190 [PubMed] [CrossRef] [Google Scholar]
- Frankel N., Erezyilmaz D.F., McGregor A.P., Wang S., Payre F., Stern D.L. (2011). Морфологическая эволюция, вызванная множеством малозаметных замен в регуляторной ДНК. Природа 474 598–603. 10.1038/nature10200 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Geffeney S.L., Fujimoto E. , Brodie E.D., III, Brodie E.D., Jr., Ruben P.C. (2005). Эволюционная диверсификация устойчивых к ТТХ натриевых каналов во взаимодействии хищник-жертва. Природа 434
759–763. 10.1038/nature03444 [PubMed] [CrossRef] [Google Scholar]
- Gerstein M.B., Bruce C., Rozowsky J.S., Zheng D., Du J., Korbel J.O., et al. (2007). Что такое ген после кодирования? История и обновленное определение. Рез. генома. 17 669–681. 10.1101/gr.6339607 [PubMed] [CrossRef] [Google Scholar]
- Гилберт С. Ф. (2000). «Классические гены и гены развития: различное использование генов в эволюционном синтезе», в Концепция гена в развитии и эволюции редакторы Бертон П., Фальк Р., Райнбергер Х.-Дж. (Кембридж, Нью-Йорк: Издательство Кембриджского университета;). [Google Scholar]
- Гилберт С. Ф., Эпель Д. (2009). Экологическая биология развития: интеграция эпигенетики, медицины и эволюции . Сандерленд, Массачусетс: Sinauer Associates. [Google Scholar]
- Гьювсланд А. Б., Вик Дж. О., Берд Д. А., Хантер П. Дж., Омхолт С. В. (2013). Преодоление разрыва между генотипом и фенотипом: что для этого нужно? J. Physiol. 591
2055–2066 гг. 10.1113/jphysiol.2012.248864 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Гомпель Н., Прюдом Б. (2009). Причины повторной генетической эволюции. Дев. биол. 332 36–47. 10.1016/j.ydbio.2009.04.040 [PubMed] [CrossRef] [Google Scholar]
- Graur D., Zheng Y., Azevedo R.B.R. (2015). Эволюционная классификация геномной функции. Геном Биол. Эвол. 7 642–645. 10.1093/gbe/evv021 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Griffiths P., Stotz K. (2013). Генетика и философия: введение . Кембридж: Издательство Кембриджского университета; 10.1017/CBO9780511744082 [CrossRef] [Google Scholar]
- Gujas B., Alonso-Blanco C., Hardtke C.S. (2012). Природные аллели потери функции Arabidopsis brx обеспечивают адаптацию корней к кислой почве. Курс. биол. 22
1962–1968 гг. 10.1016/j.cub.2012.08.026 [PubMed] [CrossRef] [Google Scholar]
- Hansen TF (2013). Почему эпистаз важен для отбора и адаптации. Эволюция 67 3501–3511. 10.1111/evo.12214 [PubMed] [CrossRef] [Google Scholar]
- Холлочер Х., Хэтчер Дж. Л., Дайресон Э. Г. (2000). Генетический анализ и анализ развития различий в пигментации брюшка у разных видов подгруппы Drosophila dunni . Эволюция. Междунар. Дж. Орг. Эвол. 54 2057–2071 гг. 10.1111/j.0014-3820.2000.tb01249.x [PubMed] [CrossRef] [Google Scholar]
- Хонг Р.Л., Витте Х., Соммер Р.Дж. (2008). Естественная вариация в привлечении феромонов насекомых Pristionchus pacificus включает протеинкиназу EGL-4. Проц. Натл. акад. науч. США 105 7779–7784. 10.1073/pnas.0708406105 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Houle D., Govindaraju D.R., Omholt S. (2010). Феномика: следующий вызов. Нац. Преподобный Жене. 11
855–866. 10.1038/nrg2897 [PubMed] [CrossRef] [Google Scholar]
- Johannsen W. (1911). Генотипическая концепция наследственности. утра. Нац. 45 129–159. 10.1086/279202 [CrossRef] [Google Scholar]
- Jones F.C., Grabherr M.G., Chan Y.F., Russell P., Mauceli E., Johnson J., et al. (2012). Геномные основы адаптивной эволюции трехиглой колюшки. Природа 484 55–61. 10.1038/nature10944 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Камберов Ю.Г., Ван С., Тан Дж., Гербо П., Уорк А., Тан Л. и др. (2013). Моделирование недавней эволюции человека у мышей путем экспрессии выбранного варианта EDAR. Сотовый 152 691–702. 10.1016/j.cell.2013.01.016 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Камменга Дж. Э., Дорошук А., Риксен Дж. А. Г., Хазендонк Э., Спиридон Л., Петреску А.-Й. , и другие. (2007). А Caenorhabditis elegans дикого типа не поддается правилу «температура-размер» из-за полиморфизма одного нуклеотида в tra-3. PLoS Genet 3:е34
10.1371/journal.pgen.0030034 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Keller E. F. (2010). Мираж пространства между природой и воспитанием . Дарем, Северная Каролина: Издательство Университета Дьюка; 10.1215/9780822392811 [CrossRef] [Google Scholar]
- Киршнер М., Герхарт Дж. (1998). Эволюционируемость. Проц. Натл. акад. науч. США 95 8420–8427. 10.1073/pnas.95.15.8420 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Кислухин Г., Кинг Э. Г., Уолтерс К. Н., Макдональд С. Дж., Лонг А. Д. (2013). Генетическая архитектура токсичности метотрексата аналогична Drosophila melanogaster и людям. 3 1301–1310 гг. 10.1534/g3.113.006619 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Landry CR, Rifkin S.A. (2012). Генотип-фенотипические карты системной биологии и количественной генетики: различные и взаимодополняющие. Доп. Эксп. Мед. биол. 751
371–398. 10.1007/978-1-4614-3567-9_17 [PubMed] [CrossRef] [Google Scholar]
- Langlade N.B., Feng X., Dransfield T., Copsey L., Hanna A.I., Thébaud C., et al. (2005). Эволюция через генетически контролируемое аллометрическое пространство. Проц. Натл. акад. науч. США 102 10221–10226. 10.1073/pnas.0504210102 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Lenser T., Theißen G. (2013). Молекулярные механизмы, вовлеченные в конвергентное одомашнивание сельскохозяйственных культур. Тенденции Растениевод. 18 704–714. 10.1016/j.tplants.2013.08.007 [PubMed] [CrossRef] [Google Scholar]
- Lettre G. (2011). Последние успехи в изучении генетики роста. Гул. Жене. 129 465–472. 10.1007/s00439-011-0969-x [PubMed] [CrossRef] [Google Scholar]
- Левонтин Р. (1974a). Генетическая основа эволюционных изменений (Колумбийская биологическая серия) . Колумбия: Издательство Колумбийского университета. [Google Scholar]
- Левонтин Р. К. (1974б). Дисперсионный анализ и анализ причин. утра. Дж. Хам. Жене. 26
400–411. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Левонтин Р. (1978). Приспособление. Науч. Являюсь. 293 212–228. 10.1038/scientificamerican0978-212 [PubMed] [CrossRef] [Google Scholar]
- Linnen C.R., Poh Y.-P., Peterson B.K., Barrett R.D.H., Larson J.G., Jensen J.D., et al. (2013). Адаптивная эволюция нескольких признаков посредством множественных мутаций в одном гене. Наука 339 1312–1316 гг. 10.1126/science.1233213 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Liu F., Wen B., Kayser M. (2013). Красочные полиморфизмы ДНК у человека. Семин. Сотовый Дев. биол. 24 562–575. 10.1016/j.semcdb.2013.03.013 [PubMed] [CrossRef] [Google Scholar]
- Luong H.T.T., Chaplin J., McRae A.F., Medland S.E., Willemsen G., Nyholt D.R., et al. (2011). Вариации BMPR1B, TGFRB1 и BMPR2 и контроль дизиготного двойникования. Двойной рез. Гум. Жене. Выключенный. Дж. Междунар. соц. Твин Стад. 14
408–416. 10.1375/twin.14.5.408 [PubMed] [CrossRef] [Google Scholar]
- Линч М., Уолш Б. (1998). Генетика и анализ количественных признаков . Сандерленд, Массачусетс: Sinauer Associates. [Google Scholar]
- Маккаллум Д. М., Кост А., Ишер Ф., Якобсен М. Д., Оддс Ф. К., Санглард Д. (2010). Генетическое исследование механизмов устойчивости к азолам у Candida albicans и их проверка на модели диссеминированной инфекции на мышах. Антимикроб. Агенты Чемотер. 54 1476–1483 гг. 10.1128/AAC.01645-1649 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Manske M., Miotto O., Campino S., Auburn S., Almagro-Garcia J., Maslen G., и другие. (2012). Анализ разнообразия Plasmodium falciparum при естественных инфекциях методом глубокого секвенирования. Природа 487 375–379. 10.1038/nature11174 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Martin A. , Orgogozo V. (2013a). Данные из: Локусы повторяющейся эволюции: Каталог генетических горячих точек фенотипической изменчивости .
Доступно по адресу: http://datadryad.org/resource/
10.5061/dryad.v66p0 [PubMed] [CrossRef] [Google Scholar]
- Мартин А., Оргогозо В. (2013b). Локусы повторной эволюции: каталог генетических очагов фенотипической изменчивости. Эволюция. Междунар. Дж. Оргн. Эвол. 67 1235–1250 гг. 10.1111/evo.12081 [PubMed] [CrossRef] [Google Scholar]
- Mayr E. (1963). Виды животных и эволюция . Гарвард: Издательство Гарвардского университета; 10.4159/harvard.9780674865327 [CrossRef] [Google Scholar]
- МакГэри К.Л., Парк Т.Дж., Вудс Дж.О., Ча Х.Дж., Уоллингфорд Дж.Б., Маркотт Э.М. (2010). Систематическое открытие неочевидных моделей болезней человека с помощью ортологических фенотипов. Проц. Натл. акад. науч. США 107 6544–6549. 10.1073/pnas.0 0107 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Мери Ф. , Белай А. Т., Со А. К.-К., Соколовский М. Б., Кавецкий Т. Дж. (2007). Природный полиморфизм, влияющий на обучение и память у дрозофилы . Проц. Натл. акад. науч. США 104
13051–13055. 10.1073/pnas.0702923104 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Moss L. (2003). Один, два (тоже?), много генов? Q. Rev. Biol. 78 57–67. 10.1086/367581 [CrossRef] [Google Scholar]
- Narod SA, Foulkes WD (2004). BRCA1 и BRCA2: 1994 г. и последующие годы. Нац. Преподобный Рак 4 665–676. 10.1038/nrc1431 [PubMed] [CrossRef] [Google Scholar]
- Натараджан С., Иногути Н., Вебер Р. Э., Фаго А., Морияма Х., Сторц Дж. Ф. (2013). Эпистаз среди адаптивных мутаций гемоглобина оленьих мышей. Наука 340 1324–1327 гг. 10.1126/science.1236862 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Noble D. (2006). Музыка жизни: биология за пределами генома .
Оксфорд: Издательство Оксфордского университета. [Google Scholar]
- Осборн К. А., Робишон А., Берджесс Э., Батланд С., Шоу Р. А., Култхард А. и др. (1997). Естественный полиморфизм поведения, обусловленный цГМФ-зависимой протеинкиназой Drosophila . Наука 277 834–836. 10.1126/science.277.5327.834 [PubMed] [CrossRef] [Google Scholar]
- Ояма С. (2000). Онтогенез информации: системы развития и эволюция 2-е изд. Кембридж, Нью-Йорк: Издательство Университета Дьюка; 10.1215/9780822380665 [CrossRef] [Google Scholar]
- Пааби А. Б., Рокман М. В. (2013). Многоликость плейотропии. Тенденции Жене. 29 66–73. 10.1016/j.tig.2012.10.010 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Палмер А. Р. (2004). Нарушение симметрии и эволюция развития. Наука 306 828–833. 10.1126/science.1103707 [PubMed] [CrossRef] [Google Scholar]
- Paterson A.H., Lin Y.R., Li Z., Schertz K.F., Doebley J.F., Pinson S.R., et al. (1995). Конвергентное одомашнивание злаковых культур путем независимых мутаций в соответствующих генетических локусах. Наука 269
1714–1718 гг. 10.1126/science.269.5231.1714 [PubMed] [CrossRef] [Google Scholar]
- Перуц М. Ф. (1983). Видовая адаптация в белковой молекуле. Мол. биол. Эвол. 1 1–28. [PubMed] [Академия Google]
- Phillips PC (2008 г.). Эпистаз – существенная роль взаимодействий генов в структуре и эволюции генетических систем. Нац. Преподобный Жене. 9 855–867. 10.1038/nrg2452 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Pokorná M., Kratochvíl L. (2009). Филогения механизмов определения пола у чешуйчатых рептилий: половые хромосомы — эволюционная ловушка? Зоол. Дж. Линн. соц. 156 168–183. 10.1111/j.1096-3642.2008.00481.x [CrossRef] [Google Scholar]
- Робинсон П. Н., Уэббер К. (2014). Онтологии фенотипов и межвидовой анализ для трансляционных исследований. PLoS Genet 10:e1004268 10.1371/journal.pgen.1004268 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Rockman MV (2012). Программа QTN и аллели, имеющие значение для эволюции: все золото не блестит. Эволюция. Междунар. Дж. Оргн. Эвол. 66
1–17. 10.1111/j.1558-5646.2011.01486.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Роджерс В. А., Саломоне Дж. Р., Тейси Д. Дж., Камино Э. М., Дэвис К. А., Ребейз М. и др. (2013). Рекуррентная модификация консервативного цис-регуляторного элемента лежит в основе разнообразия пигментации плодовых мушек. PLoS Genet 9:e1003740 10.1371/journal.pgen.1003740 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Romero-Herrera AE, Lehmann H., Joysey KA, Friday AE (1978). Об эволюции миоглобина. Филос. Транс. Р. Соц. Лонд. Б. биол. науч. 283 61–163. 10.1098/rstb.1978.0018 [PubMed] [CrossRef] [Google Scholar]
- Salazar-Ciudad I., Marín-Riera M. (2013). Адаптивная динамика по картам генотип-фенотип, основанным на развитии. Природа 497 361–364. 10.1038/nature12142 [PubMed] [CrossRef] [Google Scholar]
- Саркар С. (1999). От нормы реакции к норме адаптации: норма реакции, 1909–1960 гг. Биол. Филос. 14
235–252. 10.1023/A:10066648 [CrossRef] [Google Scholar]
- Швандер Т., Леймар О. (2011). Гены как лидеры и последователи эволюции. Тенденции экол. Эвол. 26 143–151. 10.1016/j.tree.2010.12.010 [PubMed] [CrossRef] [Google Scholar]
- Шварц С. (2000). «Дифференциальная концепция гена: прошлое и настоящее». Концепция гена в развитии и эволюции редакторы Бертон П. Дж., Фальк Р., Райнбергер Х. Дж. (Кембридж, Нью-Йорк: издательство Кембриджского университета;) 26–39. 10.1017/CBO9780511527296.004 [CrossRef] [Google Scholar]
- Ширанги Т. Р., Дюфур Х. Д., Уильямс Т. М., Кэрролл С. Б. (2009). Быстрая эволюция экспрессии фермента, продуцирующего половые феромоны, у Drosophila . PLoS Биол. 7:e1000168 10.1371/journal.pbio.1000168 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Simondon G. (1968). L’individuation à la Lumière des Notions de Forme et d’Information 2005-е изд. Гренобль: Миллон. [Google Scholar]
- Собер Э. (1988). Распределение причинной ответственности. Дж. Филос. 85 303–318. 10.2307/2026721 [CrossRef] [Академия Google]
- Steiner C.C., Weber JN, Hoekstra HE (2007). Адаптивная вариация у пляжных мышей, вызванная двумя взаимодействующими генами пигментации. PLoS Биол. 5:e219 10.1371/journal.pbio.0050219 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Stern DL (2000). Эволюционная биология развития и проблема изменчивости. Эволюция. Междунар. Дж. Оргн. Эвол. 54 1079–1091. 10.1111/j.0014-3820.2000.tb00544.x [PubMed] [CrossRef] [Google Scholar]
- Стерн Д. Л. (2013). Генетические причины конвергентной эволюции. Нац. Преподобный Жене. 14 751–764. 10.1038/nrg3483 [PubMed] [CrossRef] [Google Scholar]
- Стерн Д., Оргогозо В. (2008). Локусы эволюции: насколько предсказуема генетическая эволюция? Эволюция. Междунар. Дж. Оргн. Эвол. 62
2155–2177. 10.1111/j.1558-5646.2008.00450.x [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Stewart C.B., Schilling J.W., Wilson A.C. (1987). Адаптивная эволюция в желудочных лизоцимах ферментеров передней кишки. Природа 330 401–404. 10.1038/330401a0 [PubMed] [CrossRef] [Google Scholar]
- Стоц К. (2012). Убийство на экспрессе развития: кто убил природу/воспитание? биол. Филос. 27 919–929. 10.1007/s10539-012-9343-1 [CrossRef] [Google Scholar]
- Sturtevant AH (1932). Использование мозаики в изучении влияния генов на развитие. Проц. Шестой межд. конгр. Жене. Итака, штат Нью-Йорк, 1 304–307. [Google Scholar]
- Supple M., Papa R., Counterman B., McMillan W. O. (2014). Геномика адаптивного излучения: понимание континуума видообразования геликониуса. Доп. Эксп. Мед. Биол . 781 249–271. 10.1007/978-94-007-7347-9_13 [PubMed] [CrossRef] [Google Scholar]
- Таутц Д., Шмид К. Дж. (1998). От генов к индивидуумам: гены развития и формирование фенотипа. Филос. Транс. Р. Соц. Лонд. Б. биол. науч. 353
231–240. 10.1098/rstb.1998.0205 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Томас Д. (2010). Исследования ассоциации генов и окружающей среды: новые подходы. Нац. Преподобный Жене. 11 259–272. 10.1038/nrg2764 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Венкатеш Б., Лу С. К., Дандона Н., См. С. Л., Бреннер С., Сунг Т. В. (2005). Генетическая основа устойчивости к тетродотоксину у иглобрюхов. Курс. биол. 15 2069–2072 гг. 10.1016/j.cub.2005.10.068 [PubMed] [CrossRef] [Google Scholar]
- Visscher PM, Brown MA, McCarthy MI, Yang J. (2012). Пять лет открытия GWAS. утра. Дж. Хам. Жене. 90 7–24. 10.1016/j.ajhg.2011.11.029[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Waddington CH (1957). Стратегия генов 2014 Эдн Репринт Лондон: Рутледж. [Google Scholar]
- Вагнер Г. П. (1996). Гомологи, природные виды и эволюция модульности. утра. Зоол. 36
36–43. 10.1093/icb/36.1.36 [CrossRef] [Google Scholar]
- Wagner GP (2000). Концепция характера в эволюционной биологии . Сан-Диего, Калифорния: Academic Press. [Академия Google]
- Вагнер Г. П. (2014). Гомология, гены и эволюционные инновации . Принстон, Нью-Джерси: Издательство Принстонского университета; 10.1515/9781400851461 [CrossRef] [Google Scholar]
- Wang J., Wurm Y., Nipitwattanaphon M., Riba-Grognuz O., Huang Y.-C., Shoemaker D., et al. (2013). Y-подобная социальная хромосома вызывает альтернативную организацию колонии у огненных муравьев. Природа 493 664–668. 10.1038/nature11832 [PubMed] [CrossRef] [Google Scholar]
- Waters CK (2007). Причины, которые имеют значение. Дж. Филос. 104 551–579. [Google Scholar]
- Велтер Д., Макартур Дж., Моралес Дж., Бердетт Т., Холл П., Джанкинс Х. и др. (2014). Каталог NHGRI GWAS, курируемый ресурс ассоциаций SNP-признаков. Рез. нуклеиновых кислот. 42
Д1001–Д1006. 10.1093/nar/gkt1229 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- West-Eberhard M. J. (2003). Пластичность развития и эволюция . Оксфорд: Издательство Оксфордского университета. [Google Scholar]
- West-Eberhard M. J. (2005). Пластичность развития и происхождение видовых различий. Проц. Натл. акад. науч. США 102 (Приложение 1) 6543–6549. 10.1073/pnas.0501844102 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Wilkins A. (2014). «Набор генетических инструментов»: история жизни важной метафоры. Достижения в эволюционной биологии развития изд. Тодд Стрилман Дж. (Хобокен, Нью-Джерси: John Wiley & Sons;) 1–14. [Google Scholar]
- Уилкинс А.С. (2002). Эволюция путей развития . Сандерленд, Массачусетс: Sinauer Associates. [Академия Google]
- Сюй С., Дун Г.-С., Ху С.-С., Мяо Л., Чжан С.-Л., Чжан Д.-Л. и др. (2013). Генетическая основа белых тигров. Курс. биол. 23
1031–1035. 10.1016/j.cub.2013.04.054 [PubMed] [CrossRef] [Google Scholar]
- Zhang J., Wagner GP (2013). Об определении и измерении плейотропии. Тенденции Жене. 29 383–384. 10.1016/j.tig.2013.05.002 [PubMed] [CrossRef] [Google Scholar]
Статьи из Frontiers in Genetics предоставлены здесь с разрешения Frontiers Media SA
Экспрессия генов и концепция фенотипа
Обзор
. 2007 март; 38(1):238-54.
doi: 10.1016/j.shpsc.2006.12.014. Epub 2007, 12 февраля.
Охад Начтоми 1 , Айелет Шавит, Зохар Яхини
принадлежность
- 1 Факультет философии, Университет Бар-Илан, Рамат-Ган 52900, Израиль. [email protected]
- PMID: 17324816
- DOI: 10.1016/ж.шпсц.2006.12.014
Обзор
Ohad Nachtomy et al. Stud Hist Philos Biol Biomed Sci. 2007 9 марта0011
. 2007 март; 38(1):238-54.
doi: 10.1016/j.shpsc.2006.12.014. Epub 2007, 12 февраля.
Авторы
Охад Начтоми 1 , Айелет Шавит, Зохар Яхини
принадлежность
- 1 Факультет философии, Университет Бар-Илан, Рамат-Ган 52900, Израиль. [email protected]
- PMID: 17324816
- DOI: 10.1016/ж.шпсц.2006.12.014
Абстрактный
Если определение «генотипа» претерпело кардинальные изменения при переходе от классической генетики к молекулярной, то определение «фенотипа» долгое время оставалось в классических рамках. Кроме того, в то время как понятие генотипа привлекло значительное внимание философов биологии, понятие фенотипа — нет. Недавние разработки в технологии измерения уровней экспрессии генов позволили представить фенотипические признаки с точки зрения уровней экспрессии генов. Мы демонстрируем, что это не только стало возможным, но и стало реальной практикой. Это предполагает существенное изменение нашей концепции фенотипа: как и в случае с «генотипом», фенотипы теперь могут быть представлены в количественных и измеримых терминах на всестороннем молекулярном уровне. Мы обсуждаем, в каком смысле профили экспрессии генов можно рассматривать как фенотипические признаки и лучше ли эти признаки описывать как новую концепцию фенотипа или как расширение классической концепции. Мы выступаем за расширение классической концепции и призываем к исследованию типа вовлеченного расширения.
Похожие статьи
Генетическая геномика: сочетание генетики с анализом экспрессии генов.
Ли Дж., Бурмейстер М. Ли Дж. и др. Хум Мол Жене. 15 октября 2005 г .; 14 Спецификация № 2: R163-9. дои: 10.1093/hmg/ddi267. Хум Мол Жене. 2005. PMID: 16244315 Обзор.
Геномные нормы реакции: использование интегративной биологии для понимания молекулярных механизмов фенотипической пластичности.
Aubin-Horth N, Renn SC.
Обин-Хорт Н. и соавт.
Мол Экол. 2009 сен; 18 (18): 3763-80. doi: 10.1111/j.1365-294X.2009.04313.x. Epub 2009 1 сентября.
Мол Экол. 2009.
PMID: 19732339
Обзор.Поиск ассоциаций генотип/фенотип и сканирование фенотипа.
Джонс Р., Пембри М., Голдинг Дж., Херрик Д. Джонс Р. и соавт. Педиатр Перинат Эпидемиол. 2005 июл; 19(4): 264-75. doi: 10.1111/j.1365-3016.2005.00664.x. Педиатр Перинат Эпидемиол. 2005. PMID: 15958149 Обзор.
От феномена к фенотипу и от фенотипа к гену: генетика вперед и проблема сепсиса.
Beutler B, Du X, Hoebe K. Бейтлер Б. и др. J заразить Dis. 2003 г., 15 июня; 187 Приложение 2: S321-6. дои: 10.1086/374757. J заразить Dis. 2003. PMID: 12792846
Естественная изменчивость RPS2-опосредованной устойчивости среди образцов арабидопсиса: корреляция между профилями экспрессии генов и фенотипическими ответами.
Ван Поке Р.М., Сато М., Ленарц-Вятт Л., Вайсберг С., Катагири Ф. Ван Поке Р.М. и др. Растительная клетка. 2007 Декабрь; 19 (12): 4046-60. doi: 10.1105/tpc.107.053827. Epub 2007, 14 декабря. Растительная клетка. 2007. PMID: 18083910 Бесплатная статья ЧВК.
Посмотреть все похожие статьи
Цитируется
Наследование экспрессии генов на протяжении развития плода у перца чили.
Эското-Сандовал К., Очоа-Алехо Н., Мартинес О. Эското-Сандовал С. и др. Научный представитель 2021 22 ноября; 11 (1): 22647. doi: 10.1038/s41598-021-02151-z. Научный представитель 2021. PMID: 34811443 Бесплатная статья ЧВК.
Ловля вперед и назад: достижения в области феномики рыбок данио.
Фуэнтес Р., Летелье Дж., Таджер Б., Вальдивия Л.Е., Маллинз М.С. Фуэнтес Р. и соавт. Мех Дев. 2018 Декабрь; 154: 296-308. doi: 10.1016/j.mod.2018.08.007. Epub 2018 18 августа. Мех Дев. 2018. PMID: 30130581 Бесплатная статья ЧВК. Обзор.
PRECOG: инструмент для автоматизированного извлечения и визуализации компонентов пригодности в феноменологии микробного роста.
Фернандес-Рико Л., Курченко О., Закриссон М., Уоррингер Дж., Бломберг А. Фернандес-Рико Л. и соавт. Биоинформатика BMC. 2016 23 июня; 17:249. doi: 10.1186/s12859-016-1134-2. Биоинформатика BMC. 2016. PMID: 27334112 Бесплатная статья ЧВК.
Социальный паразитизм и молекулярная основа фенотипической эволюции.
Чини А.
, Паталано С., Сегондс-Пишон А., Басби Г.Б., Черво Р., Самнер С.
Чини А. и др.
Фронт Жене. 2015 18 фев; 6:32. дои: 10.3389/fgene.2015.00032. Электронная коллекция 2015.
Фронт Жене. 2015.
PMID: 25741361
Бесплатная статья ЧВК.Еда как воздействие: эпигенетика питания и новый метаболизм.
Ландекер Х. Ландекер Х. Биосообщества. 2011 июнь;6(2):167-194. doi: 10.1057/biosoc.2011.1. Epub 2011 7 марта. Биосообщества. 2011. PMID: 23227106 Бесплатная статья ЧВК.
Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Генотип – определение и примеры
Генотип
сущ. , множественное число: генотипы
[ˈdʒiːn.oʊ.taɪp]
Определение: генетическая структура, кодирующая признак
Содержание
Определение генотипа
Генотип — это генетическая структура отдельной клетки или организма, которая определяет или вносит вклад в его фенотип. Противоположные термины генотип и фенотип используются для определения характеристик или признаков организма. Генотип определяет аллели, относящиеся к одному признаку (например, Аа) или к ряду признаков (например, Аа Вb cc). Термин может быть расширен для обозначения всего набора генов организма (или таксона). На этом всестороннем уровне он несколько совпадает с термином геном, который относится ко всей ДНК в организме. Однако не вся ДНК состоит из генов. Чтобы полностью описать генотип организма, потребуется набор символов, представляющих аллели для каждого из примерно 20 000 известных генов в геноме человека.
Какой интересный способ учиться! Приходите и присоединяйтесь к нам в нашей дискуссии на форуме: Что является ключом к признанию кодоминирования?
Фенотип организма описывает физические или физиологические особенности организма и является следствием экспрессии генотипов. Однако генотип — не единственный фактор, определяющий фенотип индивидуума. Различные факторы окружающей среды также играют роль, и они влияют на экспрессию генов, определяя отличительный фенотип организма.
Гены, генотипы и аллели
У большинства эукариот гены представлены парами, называемыми аллелями. Парные аллели занимают одно и то же место на хромосоме (называемое локусом). Они также будут контролировать ту же черту. Термин аллель относится к варианту определенного гена. Поскольку большинство эукариот размножаются половым путем, каждый родитель вносит один из двух аллелей данного гена. Поскольку каждый родитель является продуктом разных линий организмов, случайные ошибки репликации и взаимодействие окружающей среды с ДНК привели к различным модификациям генов каждого родителя. Это различие лежит в основе термина « аллель » ( алло = другое), что подразумевает инаковость или различие.
Если признак следует простому менделевскому типу наследования, то один из аллелей будет выражен, а другой нет. Аллель, который экспрессируется, считается доминантным аллелем, тогда как аллель, который не экспрессируется, описывается как рецессивный. В случаях, когда доминантный аллель отсутствует, будет происходить экспрессия рецессивного аллеля.
Если доминантный аллель обозначен как «А», а рецессивный аллель — «а», возможны три различных генотипа: «АА», «аа» и «Аа». Термин « гомозиготный » используется для описания пар «АА» и «аа», поскольку аллели в паре одинаковы, то есть оба доминантны или оба рецессивны. Напротив, термин « гетерозиготный » используется для описания аллельной пары «Аа». Доминантный признак будет выражен в фенотипах особей с генотипами АА или Аа, а рецессивный фенотип будет выражен у особей с генотипом аа.
Не все признаки определяются одним геном; многие гены имеют несколько аллельных вариантов, и не все пары генов имеют простое доминантно-рецессивное отношение. Кодоминирование, неполное доминирование и полигенное наследование являются примерами неменделевского типа наследования. У людей многие из наблюдаемых признаков не являются менделевскими. Рост и цвет кожи, например, обусловлены взаимодействием не только пары аллелей, но и генов в нескольких разных локусах.
Изучайте кодоминирование и неполное доминирование вместе с нами! Присоединяйтесь к нашему форуму: Что является ключом к признанию кодоминирования? Поделитесь своими мыслями!
Прогнозирование простых генотипов: квадраты Пеннета
В простых генотипах указываются аллели только одного или нескольких признаков. Например, генотип гена, определяющего окраску цветка гороха, представлен двумя аллелями В и В 9.0042 . «В» — доминантный аллель, тогда как «b» — рецессивный аллель. Доминантный аллель кодирует признак пурпурного цветка, тогда как рецессивный аллель кодирует признак белого цветка. См. рисунок диаграммы ниже.
Квадрат Пеннета, показывающий взаимосвязь между фенотипом и генотипом Эта диаграмма является примером квадрата Пеннета, впервые представленного Реджинальдом Пеннеттом после повторного открытия работы Менделя. Квадрат Пеннета является важным инструментом в генетике для предсказания моделей и соотношений наследования. Этот квадрат изображает как генотип, так и фенотип потомства двух гетерозиготных (Bb) родителей, оба с фиолетовыми цветками. На диаграмме представлены аллели, присутствующие в родительских гаметах и в потомстве, полученном в результате различных комбинаций этих гамет. Каждый родитель производит два вида гамет ( В и В ), которые объединяются во все возможные пары. Из четырех потомков два гомозиготны, один пурпурный (BB) и один белый (bb), , и два гетерозиготны (Bb), , оба экспрессируют доминантный признак пурпурный. Это дает нам фенотипическое соотношение доминантных и рецессивных фенотипов 3:1, описанное Менделем. Это также дает нам соотношение генотипов 1:2:1 9генотипы 0041 BB, Bb, и bb .
Отзыв: Мэри Энн Кларк, доктор философии
Есть вопросы по генотипу? Присоединяйтесь к нашему форуму: Что является ключом к признанию кодоминирования? Наше сообщество может помочь!
Пройди тест по биологии генотипа!
Викторина
Выберите лучший ответ.
1. Набор генов организма
Genotype
Phenotype
2. Identifies the alleles related to a single trait
Genotype
Phenotype
3. Physical and physiological features
Genotype
Phenotype
4. Homozygous recessive allele
Aa
AB
аа
5. Инструмент, используемый для прогнозирования моделей и соотношений наследования
Соотношение фенотипов
Соотношение генотипов
Квадрат Пеннета
Отправьте ваши результаты (необязательно)
Ваше имя
Отправить по электронной почте
Далее
Отличие генотипа/фенотипа (Стэнфордская энциклопедия философии/Зимнее издание 2011 г.)
1 90 Citation Друзья Предварительный просмотр PDF | Инфо Поиск | Библиография PhilPapers
Впервые опубликовано в пятницу, 23 января 2004 г . ; основная редакция Вт 26 апреля 2011 г.
Различие между фенотипом и генотипом имеет основополагающее значение для понимание наследственности и развития организмов. генотип организма – это класс, к которому относится этот организм. принадлежит, как определено описанием фактического физического материал, состоящий из ДНК, которая была передана организму его родителями при зачатии организма. Для организмов, размножающихся половым путем этот физический материал состоит из ДНК, внесшей свой вклад в оплодотворенная яйцеклетка сперматозоидом и яйцеклеткой двух ее родителей. Для бесполого размножающихся организмов, например бактерий, наследственный материал прямая копия ДНК своего родителя. фенотип организм — это класс, к которому принадлежит этот организм, определяемый описание физических и поведенческих характеристик организма, например, его размер и форма, его метаболическая активность и характер его движения.
Необходимо различать дескрипторы организма, его
генотип и фенотип, от материальных объектов, которые
описано. Генотип — это дескриптор генома , который
представляет собой набор физических молекул ДНК, унаследованных от
родители. Фенотип – это дескриптор феноме ,
проявляются физические свойства организма, его физиология,
Морфология и поведение.
Понятия фенотипа и генотипа также требуют разграничения. между типами и токенами . Как слова «генотип» и «фенотип» предполагают, что это типы, наборы, членами которых являются любой данный организм и его геном, множества, определяемые их физическим описанием. Любой индивидуальный организм и его геном являются членами этих наборов, маркерами этих типов.
- 1. Наследственность и развитие
- 2. Частичный генотип и частичный фенотип
- 3. Сопоставление генотипа с фенотипом
- 3.1 ДНК-белковые связи
- 3.2 Отношения между генами
- 3.3 Гены и окружающая среда
- 3.4 Стохастические эффекты
- 4. Заключение
- Библиография
- Академические инструменты
- Другие интернет-ресурсы
- Связанные записи
Различие между генотипом и фенотипом проводится
необходимо разделением причинных путей, ведущих к одному
стороны к передаче информации об организмах между последовательными
поколений, а с другой — к росту и развитию
организма в течение одного поколения от зачатия до смерти. Механизм
наследственность такова, что причинный путь наследования идет от
геномы в одном поколении к геномам в следующем без какого-либо влияния
на геном событий, происходящих в развитии
явления в течение жизни организма. Механизм
развитие феномена в пределах одного поколения от генома таково
что результат развития прекращается со смертью
организм. В то время как геном является элементом причинного пути, ведущего
от первой стадии жизни организма до конечной
индивидуально, нет обратного влияния феномена
развитого организма по геному, передаваемому между поколениями. Это может
представить схематически как:
| поколение 1 | поколение 2 | поколение № | ||
|---|---|---|---|---|
| наследственность | наследственность | наследственность | ||
| ↓ | ↓ | ↓ | ||
| развитие генома | → | разработка генома | → | разработка генома |
| ↓ | ↓ | ↓ | ||
| феноме | феноме | феноме |
Различие между генотипом и фенотипом было введено
Вильгельм Йоханнсен в 1908 году вследствие осознания того, что
наследственные пути и пути развития были причинно разделены. Этот
заявление уже было открыто сделано Августом Вейсманом в конце
XIX века, который различал зародышевая плазма организма, ткань, образующая гаметы для
производят следующее поколение, и соматоплазма , ткани
остальной части тела. По Вейсману соматоплазма
развивалась и находилась под влиянием окружающей среды, тогда как зародышевая плазма
сегрегирован в начале развития и не подвержен
воздействия окружающей среды. Таким образом, не могло быть наследования
приобретенные характеристики. Различие Йоханнсена между генотипом
и фенотип, однако, был индуцирован не вейсманизмом, а
новое открытие в 1900 работ Менделя о наследовании в саду
горох.
Важнейшей особенностью результата Менделя был результат, полученный им в
первое и второе поколения скрещивания растений гороха с
четкие фенотипические различия. При чистом разведении красноцветковых
сорт был скрещен с чисто селекционной белоцветковой формой, все
потомство в первом поколении было красноцветковым. Когда, однако,
эти гибриды с красными цветками были скрещены друг с другом, оба
в потомстве появились красноцветковые и белоцветковые растения. В
для того, чтобы объяснить это необычайное повторное появление белоцветков.
растения во втором поколении, несмотря на то, что первое
скрещивание поколений дало растения только с красными цветками, Мендель
различать внутреннее состояние растений и их
внешний вид. Он постулировал наличие внутренней дискретной
элементы, «факторы», внесенные родителями
потомству. Хотя эти факторы каким-то образом взаимодействовали,
производят внешний вид растений, они физически не
смешиваются или загрязняют друг друга, но сохраняют свою дискретность
индивидуальность. Таким образом, чисто выведенное белоцветковое растение имело два белых цветка.
факторы, один из которых внес материнский родитель, а другой — его
отцовский родитель, в то время как растения с красными цветками имели два красных фактора.
гибрид между этими двумя чисто выведенными разновидностями будет иметь один белый
фактор и один красный фактор. Красные цветы этого гибрида были
следствие «доминирования» красных факторов над белыми
факторы их причинно-следственной связи в формировании окраски цветка. Что
преобладание в физиологическом действии, однако, никоим образом не повлияло на
природа самих факторов, вновь разделившихся,
незараженные, когда гибридные растения продуцировали пыльцу и семязачатки. В качестве
как следствие, при скрещивании двух гибридных растений некоторые из
потомство получило бы белый фактор как от пыльцы, так и от
семязачатки и будут иметь белые цветки.
Эта схема менделевского объяснения делает четкое различие
между тем, что мы сейчас называем «геномом», и
«феномен». Менделевские факторы являются частью генома.
а описание их состояния есть генотипическое описание.
Внешний вид растения дает описание его
фенотип. Самое главное, формальное описание свойств
причинно-следственные связи, заключенные в законах Менделя, проводят четкое разделение
между наследственными и эволюционными путями. Менделя Второй
Закон, Закон Сегрегации, утверждает, что факторы,
вместе, когда яйцеклетка оплодотворена, снова разделятся, не затронутые
их смесь в развитом организме и не подвержена влиянию их
физиологическое взаимодействие, которое производит фенотип взрослого. Третий закон Менделя, закон независимого ассортимента, утверждает, что
когда есть пары факторов для разных характеристик, скажем,
факторная пара для цвета цветка и факторная пара для высоты растения, что
сегрегация факторов окраски цветка при образовании гамет
причинно не зависит от расщепления пары факторов для растений
высота. Тогда Второй и Третий законы являются описаниями
закономерности наследования. Первый закон Менделя, закон доминирования,
напротив, есть закон развития, утверждающий, что когда два
факторы факторной пары в отдельном организме различаются по своему
физиологический эффект, в конечном результате одно будет доминировать над другим
развития.
В первой половине двадцатого века прогресс был очень незначительным.
сделано при выявлении физической основы факторов Менделя. Главный
заранее была демонстрация того, что различные факторы, теперь переименованные
«гены» располагались линейно вдоль тел в ядре
клеток, хромосом. Поведение хромосом в
процесс образования гамет был тем, что предсказывали формальные
наследственные свойства генов. Изменения в наблюдаемом
физическая форма определенных мест на хромосомах может быть связана
со специфическими изменениями фенотипа и наследственными изменениями в
Фенотип может быть получен путем бомбардировки организмов высокой энергией.
радиационное ионизирующее излучение. Но гены остались абстрактными сущностями
существование которых как элементы наследственности и причины
развитие полностью зависело от выводов из фенотипов
организмы, участвующие в различных селекционных экспериментах. То есть,
генотипы должны были быть выведены из их фенотипических эффектов.
развитие молекулярной биологии началось с окончательного
идентификация ДНК как материальной основы генов в конце
1940-х и начале 1950-х годов. Затем последовало выяснение
химическая и физическая структура ДНК, молекулярный механизм
его воспроизведение в наследственности и подробное молекулярное описание
способ, которым клетки преобразовывали информацию в ДНК генов
в молекулы физиологической и эволюционной функции. Все
этих молекулярных деталей подтвердили причинно-следственную независимость
наследственное поведение генома от его развития
функции. ДНК генома состоит из длинных нитей, состоящих из
последовательности нуклеотидов, которых всего четыре вида.
различия между генами – это различия в количестве и особенностях
порядок четырех типов нуклеотидов в генной цепочке. ДНК
реплицируется клетками путем прямого копирования цепочек ДНК в
молекул ДНК потомства. С другой стороны, чтение генотипа
информацию клетками и использование этой информации для производства
молекулы, лежащие в основе развития характеристик
фенотип осуществляется по другому пути. ДНК различных
гены сначала транскрибируются в родственную молекулу, РНК, и
разные молекулы РНК, транскрибированные с разных генов, несут
информацию, определяющую химическую структуру различных
белков, из которых строятся клетки. Информация о том, какие гены
транскрибироваться в каких клетках, в какие периоды развития и в
какое количество также содержится в участках ДНК, называемых контролирующими
или регулирующие элементы. Это транскрипция геномной ДНК
в особую отдельную молекулу РНК, которая, в свою очередь, несет
генотипическую информацию в метаболический аппарат клетки, т. критический элемент в разделении наследственного и
эволюционные функции генома. Это механизм, который
позволяет геному быть причиной фенотипа, но который, в
в то же время изолирует геном от обратного влияния
феномен, препятствующий наследованию приобретенных признаков.
Реальные организмы характеризуются большой изменчивостью от
еще один. Как правило, отдельные представители любого вида отличаются очень
большое количество нуклеотидов, составляющих их ДНК. У людей есть
составляют в среднем 3 миллиона нуклеотидных различий между любыми двумя
людей, взятых наугад. Даже очень близкие люди имеют
множество генетических различий. За исключением близнецов или отдельных
клонированы от одного и того же родителя, нет двух одинаковых организмов
геномы. Кроме того, существует некоторая двусмысленность в назначении
индивида к генотипу, потому что многие мутации происходят в клетках во время
процесс роста и развития, так что все клетки в
тела не содержат идентичных геномов. Даже бесполые одноклеточные
организмы, подобные бактериям, которые размножаются делением
родительские клетки различаются по своим геномам, потому что мутации ДНК
достаточно распространены, чтобы по крайней мере один из нуклеотидов,
составляют их ДНК, претерпят спонтанные изменения во время
деление клеток. Таким образом, генотипы — это классы только с одним
член. Более того, даже клонированные особи или однояйцевые близнецы,
хотя и идентичны по генотипу, будут отличаться друг от друга по
фенотип из-за различий в их развитии
среды. Таким образом, фенотипы также являются классами только с одним
член. Если понимать буквально, различие между генетическими и фенотипическими
типы и токены, хотя и являются логически правильными, по-видимому, не имеют никакого значения.
практический импорт.
На практике генотипические и фенотипические описания не являются полными
но частичный, ограниченный некоторым подмножеством характеристик
организм, который считается релевантным для конкретного объяснения или
экспериментальная цель. При разграничении частичных фенотипических и
генотипических описаний необходимо принять два решения. Во-первых, особое
аспект всего фенотипа выбирается для описания, скажем, скорость
производства пигмента меланина в результате биохимической обработки
небольшие молекулы с помощью ферментов, что затем диктует, что гены, которые
код ферментных белков, участвующих в реакции, является частью
интересующий частичный генотип. Во-вторых, необходимо принять решение о
какой набор фенотипов и генотипов следует рассматривать как
неразличимы и поэтому должны быть включены в определения
частичные генотипические и фенотипические классы. В случае меланина
пигментации наблюдается сплошное распространение признака от очень
от светлого до очень темного цвета из-за изменений окружающей среды и
из-за небольших различий от человека к человеку в
фактические показатели ферментативной активности. Какой диапазон отложения меланина
будут считаться принадлежащими к одному и тому же фенотипическому классу?
возникает необходимость установления границ для фенотипических классов
потому что, хотя интенсивность пигмента является непрерывной переменной,
генотипы по своей природе являются дискретными классами. Таким образом, проблема
должны быть рассмотрены фенотипические границы класса.
Также неясно, какие наборы генотипов следует включать
внутри определенного частичного генотипического класса по трем причинам. Маленький
вариации фенотипа возникают отчасти из-за взаимосвязей
метаболические пути в организме настолько сложны, что вариации
белки, которые не являются непосредственно частью пути производства меланина
тем не менее может влиять на скорость образования меланина. Но
эти изменения в таких периферических белках, в свою очередь, являются результатом
генетических вариаций в генах, которые их кодируют. Если
гены, кодирующие эти белки, должны быть включены в определение
частичный генотип, и в этом случае количество генотипических классов в
частичный генотип становится очень большим? Но проблема рекурсивная,
потому что белки, кодируемые «вторичными» генами
в свою очередь, на темпы их деятельности влияют еще и другие
белки и так далее, пока все гены в геноме не будут включены
в «частичном» генотипическом описании. Во-вторых,
связь между последовательностью ДНК и содержанием аминокислот в белках
много к одному. Код ДНК избыточен, поэтому триплеты,
отличаются только своим третьим положением, не влияют на то, какая аминокислота
указано, поэтому с этой точки зрения все избыточные варианты
включены в один и тот же частичный генотипический класс. С другой стороны, такие
изменение третьей позиции может повлиять на скорость чтения ячейки
код ДНК, и поэтому такая вариация действительно может эффективно очертить
разные генотипы. Наконец, существуют вариации в ДНК
геном вне кодирующих областей генов, которые не влияют на
химическую структуру белков-ферментов, но влияют на скорость их
синтез. Это вариации в ДНК так называемых
«контролирующие элементы», которые являются частью каждого гена и
которые влияют на скорость, с которой клетка будет считывать ген при создании
белки.
Проблема того, какие части генома и фенома должны быть
включены в частичное генотипическое и фенотипическое описание
организм в частных случаях является одним из самых проблемных в
биология. Хотя несомненно верно, что каждая часть генома
причинно связано с явлением каким-либо путем, это просто
невозможно учесть все пути связи. Иногда
биологи выбирают частичный генотип и частичный фенотип из-за
практические ограничения на то, сколько можно сделать экспериментально и
просто притворитесь для удобства, что остальные
организм действительно постоянен. Иногда они явно признают
гетерогенность остального генома и фенома, но утверждают, что это
фактически причинно не имеет отношения к явлениям,
расследования, а иногда они явно признают причинную
релевантность остальной части генома и фенома, но относиться к ней как к
фоновый экспериментальный «шум», который можно усреднить
в достаточно большом наборе наблюдений.
Утверждение, что остальные геномные и феномические эффекты могут быть
игнорируется или усредняется, зависит от предположения, что эти пути
причинно ортогональны путям, анализируемым в
наблюдения. То есть разные частичные генотипы и фенотипы
которые являются предметом анализа и эксперимента не являются
по-разному зависит от вариаций в остальной части генома и
феномен. Заявление о достоверности этого предположения является частью
приверженность редукционизму аналитической биологии, что организмы могут
понять, разрезав их на интуитивно разумные маленькие
частей, отдельного изучения которых будет достаточно, чтобы
понимание целого. Нет никаких сомнений в том, что организмы
совокупность эффективных подсистем, внутри которых причинные взаимодействия
сильны и между которыми они слабы. Сложность в том, что
границы этих подсистем не фиксируются, а меняются от функции
функционировать, поэтому они должны быть определены из наблюдений на ad hoc
основе от случая к случаю. Глаза и ноги развиваются
независимый. Существует генетическая изменчивость, влияющая на размер и форму
одного без какого-либо влияния на размер и форму другого, и
травмы развития могут сделать животное слепым, не делая его
хромой. Однако движение по неровной поверхности зависит от
координация сокращения мышц ног с предиктивной
визуальные подсказки, так что проблема передвижения заставляет этих двух
независимые от развития подсистемы, входящие в состав одной и той же функциональной
Ед. изм.
Если бы механизмы развития были таковы, что каждое изменение в
генотип приводил к другому фенотипу и каждому другому
фенотип был следствием различия в генотипе, исследование
происхождения органических вариаций было бы значительно упрощено. Данный
знание фенотипа, лежащий в основе причинный генотип может быть
однозначно и наоборот. Проблема понимания
тогда явная изменчивость среди организмов сведется к
обеспечивая механическую историю цепи биохимических реакций,
начиная с прочтения генома клеткой и заканчивая
конечном состоянии, подобно тому, как производство автомобиля может быть
полностью реконструированный по чертежам, описание
используемых материалов, производственного оборудования и порядка, в котором
материалы проходят через это оборудование. Однако фактический
соответствие между генотипом и фенотипом — это отношение многие-многие
в котором любой данный генотип соответствует множеству различных фенотипов
и существуют различные генотипы, соответствующие данному
фенотип. Современное состояние изучения развития организмов
игнорирует эту связь «многие-многие» и строится по модели
автосборочный завод. это не то развитие
биологи не знают о много-много отношениях между генотипом
и фенотип. Скорее, прагматические соображения диктуют, что
понимание механизмов развития лучше всего достигается
сосредоточив внимание сначала на тех результатах развития, которые
однозначная связь между генотипом и фенотипом, оставляя для
будущее вопросы, поставленные отношением многие-многие. Непреднамеренный
побочным продуктом этого стратегического решения является то, что язык, используемый для
описывать проблематику и результаты исследования, создавать и
подкрепляют слишком простой взгляд на взаимосвязь между генами и
персонажи.
Отображение многие-многие между генотипом и фенотипом возникает из четырех
источники: (1) связь между последовательностью ДНК и химическим
структура белков; (2) отношения между продуктами
транскрипция и трансляция информации, закодированной в геноме;
(3) зависимость развития и физиологии как от генотипа
организма и временная последовательность сред, в которых
организм развивается и функционирует; (4) стохастические вариации
Молекулярные процессы внутри клеток.
3.1 ДНК-белковые связи
Белок состоит из цепочки аминокислот, каждая из которых кодируется триплетом нуклеиновых кислот в цепочке ДНК составляющих ген. Чтобы белок имел физиологическую активность идентичность многих из этих аминокислот имеет важное значение. Таким образом, изменение любая часть гена, вызывающая замену любого из этих аминокислоты будут препятствовать физиологической активности белка. Это невозможно сказать по фенотипу, отсутствие физиологическая активность белка, какое изменение генотипа имеет произошел. Это наиболее распространенная форма отображения «многие к одному». генотип на фенотип.
3.2 Отношения между генами
Наблюдения Менделя представляют собой классический пример двусмысленности в
соотношение между генотипом и фенотипом. Он заметил, что растения
которые несли один член пары генов, определяющей красные цветы, и один
член, указывающий, что белые цветы неотличимы от растений
несущие две копии красной формы гена. Он наблюдал подобные
доминирование одной формы гена и рецессивность альтернативного гена
форму в других персонажах, а также приводит его к обобщению
явление как закон, Закон Доминирования. При последующем исследовании
показал, что доминирование одной копии гена над другой далеко
от универсального, достаточно распространено, что большая часть
генетическая изменчивость, присутствующая в популяциях организмов, скрыта в
уровне фенотипа и требует специальных экспериментальных методов для
раскрыть это. Это особенно верно, когда одна копия гена
дефектный, так что вырабатывается белок с активностью ниже нормальной
из этой копии, в то время как альтернативная нормальная копия кодирует белок, который
физиологически достаточно активен, чтобы произвести нормальный фенотип
(Фишер 1931; Холдейн 1939; Райт 1934).
Вторая форма взаимодействия, которая чрезвычайно распространена, это то, что происходит
между продуктами, считываемыми с разных генов в геноме. Если продукты
чтения различных генов необходимы для получения физиологического
эффект, то изменения в любом из генов блокируют эффект. Такой
взаимодействия происходят, когда физиологический эффект является результатом цепочки
химические этапы, каждый из которых опосредуется продуктом другого гена. За
Например, окраска шерсти у млекопитающих является результатом действия продуктов
три разных гена. Один определяет распределение пигмента в волосах,
а другой определяет, является ли цвет пигмента черным или коричневым.
Различные комбинации разных генотипов этих генов соответствуют
разные цвета шерсти. Однако существует третий ген, кодирующий фермент
это необходимо для выражения любого цвета. Если этот ген
дефектная шерсть будет белой, независимо от генотипа другого
гены (Райт 1925).
Третий источник отношения «многие к одному» между генотипом и
фенотип — феномен буферизации развития, механизм которого
плохо понимается. Существует множество фенотипических признаков организмов.
которые не показывают различий между особями, принадлежащими к одному и тому же
виды или даже постоянны среди многих родственных видов. Например
все особи всех видов плодовых мушек рода Drosophila имеют
ровно три простых световых рецептора, глазков , расположенных в
симметричный треугольник на средней линии макушки головы. Самое простое предположение состоит в том, что для этого нет генетической изменчивости.
признак и что его развитие устойчиво к нормальным условиям окружающей среды.
нарушение. Если развитие мухи достаточно нарушено,
однако наблюдаются некоторые мухи с двумя или меньшим количеством глазков. Если те
с менее чем тремя глазками используются в качестве родителей для следующих
поколение они произвели больше аномальных мух, чем родительское
поколение. Когда идет процесс селекции от аномальных мух
продолжается на протяжении многих поколений, вырабатывается линия мух,
постоянно имеет два глазка, даже при отсутствии какого-либо внешнего
нарушение развития и эти глазки могут быть симметричными или
расположены асимметрично (Мейнард Смит и Сондхи 1960). Успех
такого селекционного эксперимента доказывает, что существовал генотипический
вариация количества и расположения глазков в оригинале
популяция нормальных мух, но все разные генотипы
сопоставляются с одним и тем же фенотипом. Это феномен канализация развития , в которой имеется буферизация
развития против возмущающих сил. Существует генетическая изменчивость
среди особей по генам, влияющим на число глазков, но
эффекты развития этой вариации предотвращаются системой
буферизация (Уоддингтон 1953, 1957). Если достаточно большой
возмущения, буферность развития
преодолены, и выявлена генетическая изменчивость числа глазков. Это
тогда можно отобрать генотипы, настолько экстремальные по своим
влияние на развитие, что они находятся за пределами буферной способности
нормально развивающейся системы и производить необычные фенотипы даже при
нормальные обстоятельства. Эксперименты с различными другими постоянными функциями
различных животных показали, что канализация развития является
общий признак, так что фенотипическое единообразие нельзя рассматривать как
демонстрация соответствующего генотипического единообразия (Rendel 1967; ДеВиссер и др. . 1993).
3.3 Гены и окружающая среда
Полная последовательность ДНК организма не содержит
информация, необходимая для уточнения организма. Результат
процессы развития зависят как от генотипа, так и от
временная последовательность условий, в которых развивается организм.
Более того, отображение разных генотипов в фенотипы в одном
окружение часто бывает совершенно непредсказуемым по их отображению в
другая среда. Классическая демонстрация сложности
это картирование является экспериментом на клонах растения Ахиллея (Клаузен, Кек и Хизи, 1958). Отдельные незрелые растения были
собраны из природы, и из каждого растения было получено три клона
простым методом разрезания их на три части. Один кусок
каждое растение было выращено на небольшой высоте в горах Сьерра, одно на
средней высоты и один на большой высоте. Результат роста
на трех высотах заключалась в том, что относительные высоты различных
растения были непредсказуемы из одной среды в другую. Например,
генотип, который рос самым высоким на небольшой высоте, был самым низким на
средней высоты и второй по высоте на большой высоте. Более того,
где, поскольку этот генотип цвел на низких и больших высотах, он не смог
цветут на средней высоте, в то время как другие генотипы цветут на этой высоте. высоте, но не на большой высоте. На самом деле не было
корреляции между растениями в их росте в разных
среды. Множество экспериментов на самых разных организмах, где
было возможно произвести несколько особей одного и того же генотипа
показывают тот же результат (Левонтин и Госс, 2004).
Если нанести на график фенотип организма данного генотипа
по отношению к переменной окружения получаемая функция
называется нормой реакции генотипа
(Шмальгаузен, 1949). Это функция отображения окружающей среды в
фенотип для этого генотипа. Общеизвестно, что нормы
реакции разных генотипов представляют собой кривые неправильной формы, которые
пересекаются друг с другом. Таким образом, невозможно предсказать фенотипы
различных генотипов в новых условиях. Есть, конечно, некоторые
генотипы, которые настолько дефектны, что не выживут ни в
окружающая среда. Но они не типичны для естественной изменчивости.
среди организмов. Можно принять за общее правило, что результат
развитие любого генотипа является уникальным следствием взаимодействия
между геномом и окружающей средой.
3.4 Стохастические эффекты
Даже полная спецификация как генотипа, так и временного
порядок среды развития недостаточен для предсказания
фенотип. Если левая и правая стороны двустороннего
исследуют «симметричный» организм, он будет обнаружен
что, в общем, что он несимметричен, но что направление и
степень асимметрии варьируется от человека к человеку без
средняя разница между сторонами. Так, мухи имеют небольшие сенсорные
щетинки с левой и правой стороны. У одного человека будет, скажем,
, шесть щетинок справа и восемь слева, а еще одна
пять справа и семь слева. В среднем по многим
лиц число одинаковое с обеих сторон, но есть
колеблющаяся асимметрия от мухи к мухе. Люди, не имеют того же
отпечатки пальцев на левой и правой руках и различия в
картина может быть настолько велика, что никакого сходства не может быть обнаружено вообще. Пока что
гены левой и правой стороны одинаковые и не обычные
значение среды позволит, чтобы левая и правая руки
плод в утробе матери имеет разное развитие
среды.
Другим важным явлением случайной изменчивости является асинхронность клеточные деления. Одна бактериальная клетка, инокулированная в большую колбу постоянно перемешиваемой среды разделится на две клетки примерно через час. Каждая из этих двух клеток разделится примерно через час, но не одновременно. Их дочерние клетки снова будут делиться, но каждая на несколько минут раньше или позже остальных и так далее, пока не популяция клеток постоянно растет во времени без синхронизация деления. Тем не менее, на ранних стадиях развития культуры не хватило поколений, чтобы накапливают мутации, поэтому клетки генетически идентичны, а существует любая возможность различных сред в постоянно перемешиваемая культура. Такая же асинхронность делений возникает на всех стадиях при делении клеток оплодотворенной яйцеклетки у эмбрионов.
Источником этих асимметрий и асинхроний является очень низкая
количество копий биологически важных больших молекул в каждом
клетка. Закон действия масс в химии, который основан на
статистическое усреднение по очень большому количеству молекул не
применяются, когда есть только три одной молекулы и семь другой. Но
именно так обстоит дело с молекулярными числами внутри
клетки. Каждый вид молекул находится в небольшом количестве, и они
распределяется по пространству внутри клетки. Чтобы произошла реакция
между молекулами они должны быть в непосредственной близости и каждая молекула в
реакция должна быть в правильном колебательном состоянии для
взаимодействие. Вибрационные состояния, в свою очередь, колеблются для каждого
молекулы, в конечном счете, как следствие квантовой неопределенности. Как
следствие стохастического изменения числа, пространственного положения
и реакционной способности каждого вида молекул, существует значительный случайный
изменение от клетки к клетке времени клеточного деления и его
исход. Если есть семь молекул определенного типа, присутствующих в
время клеточного деления, одна дочерняя клетка может получить три копии и
один четыре копии, так что потребуется разное количество времени для
эти клетки, чтобы синтезировать достаточное количество копий молекул для следующего
дивизия (Госс и Пеккуд 1998; Левонтин и Госс, 2004 г. ; МакАдамс и
Аркин 1997).
Обширные измерения флуктуирующей асимметрии и асинхронии показали, что эти стохастические эффекты являются важными источниками фенотипической изменчивости.
Сложная случайность отношений между генотипом и
фенотипы возникают из природы организмов как физических систем. Они
отличаются от физических систем, которые были объектами изучения
большинства физики и химии в двух отношениях. В отличие от атомов или
планеты они промежуточные по размеру и внутренне функционально
неоднородный. Как следствие, они являются связующим звеном очень большого
число слабо детерминирующих взаимодействующих причинно-следственных цепочек и подверженных
эффект случайного шума на всех уровнях. Последствие для
понимание строения и функций живых организмов, в том числе
их индивидуальное и социальное поведение состоит в том, что не существует какой-то мелкой
набор универсалий, таких как законы Ньютона. Даже в законах Менделя много
исключения и Биогенетический Закон всего живого из жизни не всегда может
были правдой, иначе организмов не было бы. Как верно для жизни
систем в целом отношения между генотипом и фенотипом
контингент, варьирующийся от случая к случаю.
- Аллен Г., 1979 г., «Натуралисты и экспериментаторы: генотип и фенотип», Исследования по истории биологии , 3: 179–210.
- Черчилль, Ф.Б., 1974, «Уильям Йоханнсен и генотип Понятие», Журнал истории биологии , 7: 5–30.
- Клаузен, К., Кек, Д. и В. Хизи, 1958 г., «Экспериментальные Исследования в области природы видов, III. Относящийся к окружающей среде Отклики климатических рас Achillea », Carnegie Публикация 9 Вашингтонского института0042 581: 1–129.
- Де Виссер, Дж., Хермиссон, Дж., Вагнер, Г., Мейерс, Л., Багьери-Чайчиан Х., Бланшар Дж., Чао Л., Шеверуд Дж., Елена, С., Фонтана В., Гибсон Г., Хансен Т., Кракауэр Д., Левонтин Р., Офриа К., Райс С., фон Дассов Г., Вагнер А. и Уитлок М., 1993, «Перспектива: эволюция и обнаружение генетических Надежность», Evolution , 57: 1959–1972.
- Фальк, Р. , 2001, «Может ли норма реакции спасти гены?»
Концепция», в книге «Размышления об эволюции: историческая,
Философские и политические взгляды (Том 2), Р.
С. Сингх и др. . (ред.), Кембридж: Кембриджский университет
Пресса, 119–40.
- Фишер Р., 1931 г., «Эволюция доминирования». Биологические обзоры , 6: 345–368.
- Госс, П. и Дж. Пекку, 1998 г., «Количественное моделирование Стохастические системы в молекулярной биологии с использованием стохастического анализа Петри Сети», Proceedings of the National Academy of Sciences, США , 95: 6750–6755.
- Гриффитс, ЧП, 2006 г., «Гены в постгеномной Эра», Теоретическая медицина и биоэтика , 27 (6): 499–521.
- Холдейн, JBS, 1939, «Теория эволюции Господство», Journal of Genetics , 37: 365–374.
- Каплан, Дж. М., 2008 г., «Фенотипическая пластичность и реакция
Нормы», A Companion to the Philosophy of Biology ,
С. Саркар и А. Плутински (ред. ), Малден, Массачусетс: Блэквелл,
205–22.
- Левонтин Р. и Госс П., 2004 г., «Канализация развития, Стохастичность и устойчивость», в Надежная конструкция: репертуар биологических, экологических и инженерных тематических исследований , Э. Джен (редактор), Нью-Йорк: Oxford University Press
- Мейнард Смит, Дж. и К. Сондхи, 1960, «Генетика Паттерн», Генетика , 45: 1039–1050.
- МакАдамс, Х. и А. Аркин, 1997 г., «Стохастические механизмы в генетическом анализе». Expression», Proceedings of the National Academy of Sciences, США , 94: 814–819.
- Nachtomy, O., et al. ., 2007, «Экспрессия генов и концепция фенотипа» Исследования по истории и философии Биологические и биомедицинские науки , 38 (1): 238–54.
- Rendel, J., 1967. Canalization and Gene Control , London: Академик, Логос Пресс.
- Роберт, Дж. С., 2004, Эмбриология, эпигенез и эволюция: взятие
Development Seriously , Кембридж: Издательство Кембриджского университета.
- Ролл-Хансен, Нильс, 2009 г., «Источники генотипа Вильгельма Йохансена». Theory», Journal of the History of Biology , 42 (3): 457–93.
- Шмальхаузен, И., 1949, Факторы эволюции , Филадельфия: Блэкистон.
- Waddington, C., 1953, «Генетическая ассимиляция приобретенного Персонаж», Эволюция , 7: 118–126.
- Waddington, C., 1957, Стратегия генов , Лондон: Аллен и Анвин.
- Райт, С., 1934, «Физиологические и эволюционные теории Господство», American Naturalist , 68: 25–53.
- Райт, С., 1925, «Факторы серии альбиносов Морские свинки и их влияние на черный и желтый пигмент». Генетика , 10: 223–260.
Как цитировать эту запись. Предварительный просмотр PDF-версии этой записи на Друзья общества SEP. Найдите тему этой записи в Проекте онтологии философии Индианы (ИнФО). Расширенная библиография для этой записи в PhilPapers со ссылками на его базу данных.
- Могут ли нелинейные эпигенетические взаимодействия скрыть причинно-следственные связи между генотипом и фенотипом? Андреас Вагнер (Йельский университет) в Нелинейность
биология: философия | характер/черта | Дарвинизм | биология развития | эволюция | ген | генетика: и геномика | информация: биологическая | врожденное/приобретенное отличие | молекулярная биология | естественный отбор | естественный отбор: единицы и уровни | типы и токены
Генотип и фенотип: определение, таблица и пример
На этой планете есть только один из вас – ваша ДНК не похожа ни на одну другую; это уникально. Даже генетически идентичные близнецы отличаются по внешнему виду и поведению.
Но что нас отличает? Здесь в игру вступают понятия генотипа и фенотипа. Мы быстро рассмотрим их, чтобы прояснить любые различия между ними, прежде чем углубляться в примеры и их значение в психологии.
Генотип является генетической основой организма. Он состоит из всей генетической информации, определяющей характеристики организмов.
Фенотип представляет собой наблюдаемую экспрессию этих генов. Окружающая среда влияет на фенотип.
Отличный способ запомнить это — думать о генотипе как о внутренней генетической структуре, уникальной для каждого человека, а о фенотипе — как о физическом, наблюдаемом выражении этой информации.
Примером генотипа может служить состав кожи человека и количество вырабатываемого им меланина (пигмента кожи). Примером фенотипа может быть потемнение кожи из-за УФ-излучения.
Примеры генотипа и фенотипа, оригиналы StudySmarter (изображения из Flaticon и Canva)
Генотип
Генотип относится к генетической основе организма.
ДНК, с которой мы рождаемся, является примером генотипа.
Генотип имеет решающее значение для развития фенотипа, но факторы окружающей среды, которые определяют фенотип, часто могут привести к тому, что даже генетически идентичные люди (например, однояйцевые близнецы) будут иметь значительные различия.
Исследования близнецов позволяют нам определить, насколько генотип влияет на поведение человека. В этих исследованиях рассматриваются группы близнецов и их поведение. Поскольку близнецы очень похожи генетически (100% генетическое совпадение для монозиготных близнецов и 50% для дизиготных близнецов), результаты этих исследований позволяют нам измерять и оценивать генетическую основу поведения.
Coccaro (1997) является примером этих исследований близнецов . Coccaro исследовал преступность групп монозиготных близнецов и некоторых дизиготных близнецов. Монозиготные близнецы имели такие же показатели преступности, как и дизиготные близнецы. Это открытие предполагает генетический компонент поведения, поскольку более генетически схожие близнецы демонстрировали более значительное сходство в поведении.
Также существуют исследования конкретных генов. Туарт и др. (2014) обнаружили, что из 97 участников (все мужчины), которые подвергали жестокому физическому насилию своих романтических партнеров, мужчины, у которых оказалась дисфункциональная версия гена , известная как MAOA , совершали самые ужасные, самые жестокие и садистские действия. . Эти данные свидетельствуют о том, что гены могут иметь прямое влияние на агрессивное поведение.
Как показывают данные, генетическая предрасположенность играет роль в психологическом развитии и поведении человека. Однако мы не можем говорить о генотипе, не обсуждая фенотип и разницу между ними.
Туарт и др. (2014): Прямое влияние гена MAOA на агрессивное поведение, StudySmarter Originals (изображения из Flaticon и Canva)
Фенотип
Фенотип — это наблюдаемое, измеримое выражение генов организма, на которое влияет окружающая среда.
Примером фенотипа может быть ваш высота , потому что факторы окружающей среды, такие как диета, влияют на рост.
В психологии примером фенотипа может быть то, как факторы окружающей среды, такие как семейная жизнь в детстве, могут влиять на то, как люди развиваются и ведут себя во взрослой жизни. Например, Caspi et al. (2002) обнаружили, что участники исследования, которые демонстрировали более агрессивное поведение, не только имели дисфункциональный ген MAOA, но и пережили жестокое детство . Таким образом, генотип дисфункционального гена МАОА мог быть не единственной причиной агрессивного поведения, а скорее экспрессией этого гена при воздействии агрессивных ситуаций.
Почему важно различать генотип и фенотип?
Нам необходимо различать генотип и фенотип, чтобы понять, в какой степени поведение человека определяется внутренними генетическими факторами (генотип) или влиянием окружающей среды и ситуациями, в которых возникают наблюдаемые черты и поведение (фенотип). Обычно это смесь генетических факторов (генотип) и факторов окружающей среды, которые влияют на экспрессию этих генов (фенотип), что приводит к изменениям в поведении.
Как различие между генотипом и фенотипом может помочь в решении проблем с психическим здоровьем?
Понимание различий между генотипом и фенотипом может также помочь нам понять унаследованных признаков .
Члены семьи могут иметь генетическую предрасположенность к развитию депрессии или другой проблемы с психическим здоровьем, но они могут избежать развития симптомов с помощью надлежащего лечения или условий.
Здесь важно различать генотип и фенотип, потому что:
- Некоторые люди рождаются с генетической предрасположенностью к развитию проблем с психическим здоровьем.
- Некоторые развивают их как продукт своего окружения.
- Комбинация обоих.
Их лечение может быть адаптировано к их обстоятельствам.
Различие между генотипом и фенотипом может помочь врачам более эффективно использовать свои ресурсы, когда речь идет о психическом здоровье. Пациент с семейной историей проблем с психическим здоровьем может с большей вероятностью страдать от химического дисбаланса в головном мозге, который лучше поддается медикаментозному лечению, чем терапии.
И наоборот, пациент с неизвестным семейным анамнезом проблем с психическим здоровьем, чьи проблемы с психическим здоровьем являются результатом их окружения, может помочь врачам определить, какие элементы их окружения повлияли на них и как. Этот подход может помочь с такими терапевтическими методами, как Когнитивно-поведенческая терапия (КПТ) и Диалектическая поведенческая терапия (ДПТ) .
Генотип и фенотип – Ключевые выводы
- Генотип – это генетическая структура организма.
- Гены могут влиять на поведение, например, ген MAOA.
- Фенотип – это видимый результат взаимодействия между окружающей средой и генетической структурой.
- Важно различать генотип и фенотип, чтобы понять происхождение поведения.
- Понимание различий между генотипом и фенотипом помогает лечить проблемы с психическим здоровьем.
Различия генотип-фенотип — обзор
ScienceDirectРегистрацияВход
PlusAdd to Mendeley T. Sandle, in Encyclopedia of Food Microbiology (Second Edition), 2014
Введение
Методы идентификации можно разделить на две группы: фенотипические и генотипические. Различие генотип-фенотип проводится в генетике. «Генотип» — это полная наследственная информация организма, даже если она не выражена. «Фенотип» — это реально наблюдаемые свойства организма, такие как морфология, развитие или поведение (Sutton and Cundell, 2004).
Фенотипические методы являются наиболее распространенными из-за их относительно низкой стоимости для многих лабораторий. Однако следует признать, что проявления микробного фенотипа, то есть размер и форма клеток, спорообразование, клеточный состав, антигенность, биохимическая активность, чувствительность к противомикробным агентам и т. д., часто зависят от среды и условий роста, которые был использован.
Эти условия будут включать такие переменные, как температура, pH, окислительно-восстановительный потенциал и осмоляльность, а также, возможно, менее известные переменные, такие как истощение питательных веществ, доступность витаминов и минералов, цикл роста, активность воды в твердых средах, статическая или вращающаяся жидкая культура и культура твердых сред по сравнению с жидкими, а также плотность колоний на чашке. Поэтому требуется определенная осторожность при интерпретации результатов микробиологической идентификации и анализе тенденций данных.
Еще одним ограничением фенотипических методов является размер и тип базы данных фенетической классификации. Что касается типа базы данных, многие базы данных ориентированы на клинические приложения и не обязательно хорошо подходят для промышленного применения. С точки зрения размера базы данных ограничены из-за относительно небольшого числа охарактеризованных микроорганизмов (Stager and Davis, 1992).
Классическая схема идентификации бактерий биохимическими методами зависит от того, может ли чистая культура интересующего микроорганизма расти в чашке с агаром, скошенном агаре, бульоне, бумажной полоске или другом вспомогательном материале, содержащем специализированные стимуляторы роста или ингибиторы в присутствии ферментируемого или разлагаемого соединения, что приводит к изменению цвета среды, образованию газа, образованию флуоресцентного соединения и другим проявлениям метаболической активности. Если поведение известных культур в этих средах известно, неизвестная культура может быть сопоставлена с этими характеристиками, и на основе наиболее близкого совпадения с базой данных аналитик может идентифицировать неизвестную культуру. Этот процесс утомителен, отнимает много времени и требует много труда, материалов, времени и энергии для проведения испытаний. Кроме того, умение аналитика интерпретировать реакции и прийти к правильному суждению делает этот процесс субъективным и часто ненадежным (Kalamaki et al., 19).97).
Генотипические методы не зависят от среды выделения или характеристик роста микроорганизма. Генотипические методы значительно расширили базы данных различных видов микроорганизмов. До появления генотипических методов микробиологи предполагали, что существует ряд таксонов, которые невозможно культивировать (так называемые жизнеспособные, но не культивируемые штаммы). Генотипические методы открыли совершенно новый набор видов и подвидов, а также переклассифицировали виды и родственные виды (таким образом, таксоны, которые часто сходно группируются фенотипическими методами, на самом деле являются полифилетическими группами, т. е. содержат организмы с разной эволюционной историей, которые гомологически несходные организмы, объединенные в группы).
Еще одним преимуществом генотипических методов является их точность и более быстрое получение результатов (поскольку микроорганизмы не нужно выращивать на питательных средах). Однако они относительно дороги.
Для полной идентификации можно провести множество тестов, как показано в таблице 1.
Таблица 1. Информация, необходимая для идентификации патогенов пищевого происхождения (Fung, 1995).
| Фенотипические признаки |
| Макроскопическая морфология на агаровых пластинах |
| Морфология при микроскопическом увеличении |
| Грамовая реакция (положительный, отрицательный или вариабельный) и специальные пленки |
| ). производство биолюминесценции, хемилюминесценции и флуоресцентных соединений |
| Требования к питанию и факторам роста |
| Temperature and pH requirements and tolerance |
| Fermentation products, metabolites, and toxin production |
| Antibiotic sensitivity pattern (antibiogram) |
| Gas requirements and tolerance |
| Cell wall, cell membrane, and cellular компоненты |
| Константа скорости роста и время генерации |
| Подвижность и спорообразование |
| Устойчивость к органическим красителям и специальным соединениям |
| Impedance, conductance, and capacitance characteristics |
| Genotypic characteristics |
| Genetic profile: DNA/RNA sequences and fingerprinting |
| Extracellular and intracellular products |
| Information relating to the microorganism |
| Патогенность для животных и человека |
| Серология и фаговое типирование |
| Экологическая ниша и способность к выживанию |
| Реакция на электромагнитные поля, свет, звук и излучение |
Посмотреть главуКнига покупок /science/article/pii/B9780123847300000367
Rebecca J. Многие из проблем, возникающих в связи с текущим взрывом данных о биологических последовательностях, требуют огромных вычислительных ресурсов для точного решения. Однако эти ресурсы не всегда легко доступны, и их, конечно же, не найти во многих лабораториях молекулярной биологии. Тем не менее, эти проблемы по-прежнему важны для решения молекулярных биологов. Следовательно, постоянно изучаются альтернативные вычислительные подходы. Эволюционные алгоритмы являются одним из возможных инструментов для решения таких проблем. Эти алгоритмы подходят к вычислительным задачам, используя методы выживания наиболее приспособленных и естественного отбора для выработки решений для конкретных экземпляров проблемы. Эти подходы привлекательны для решения сложных вычислительных задач и вычислительных задач, которые еще недостаточно изучены. Однако следует проявлять осторожность, поскольку эти подходы требуют понимания того, как различные компоненты алгоритмов взаимодействуют с проблемой и как эти взаимодействия влияют на эффективность эволюционного подхода. Ученые-компьютерщики традиционно классифицируют проблемы на основе их вычислительной сложности. Многие из интересных проблем, возникающих при анализе данных о биологических последовательностях и в других местах, в лучшем случае относятся к классу задач, называемых NP-трудными (описание NP-полноты и NP-трудности см. в главе 2). Как правило, задачи являются NP-трудными, потому что количество возможных альтернатив, которые необходимо рассмотреть, растет экспоненциально с размером входных данных. Например, попарное выравнивание последовательностей не является NP-трудным. Сложность этой задачи растет пропорционально произведению длины двух строк. Проблема, рассматриваемая в этой статье, проблема сборки последовательности для дробового секвенирования ДНК, является NP-полной. Рост сложности происходит экспоненциально по количеству фрагментов, потому что решения проблемы сборки — это упорядочение множества фрагментов. Существует экспоненциальное количество возможных порядков, требующих экспоненциального количества времени, чтобы определить, какой порядок является лучшим. Многие различные алгоритмы аппроксимации использовались в NP-сложных задачах, включая жадные алгоритмы, эвристические алгоритмы, методы Монте-Карло, моделируемый отжиг (вдохновленный физическим процессом отжига) и эволюционные подходы [2−4]. В этой главе рассматриваются эволюционные подходы, каждый из которых так или иначе вдохновлен процессами естественной эволюции. Многие задачи анализа последовательности являются задачами оптимизации; мы хотим найти лучший ответ на конкретный вопрос. Формулировка задачи оптимизации состоит из двух основных компонентов: пространства поиска и фитнес-функции. Аналогично различию генотип-фенотип, известному в генетическом сообществе, конкретное решение проблемы отличается от представления этого решения. Эти различия иногда бессмысленны. В частности, если существует однозначное соответствие между решениями и их кодированием в качестве решений, различия становятся менее значимыми. Во многих практических задачах существуют конкурирующие схемы представления задачи. Кроме того, может быть несколько представлений для одного и того же решения в рамках конкретной схемы представления. Конкретный выбор представления определяет пространство поиска проблемы. Компонент фитнес-функции управляет процессом оптимизации. Функция косвенно указывает правильный ответ или ответы. Выбор представления влияет на сложность поиска ответа для конкретной фитнес-функции, поскольку один и тот же выбор представления может быть хорошим или плохим в зависимости от фитнес-функции. Таким образом, при рассмотрении проблемы функция пригодности и тип используемого метода оптимизации влияют на выбор схемы представления. После описания проблемы в рамках данной оптимизации существует несколько возможных подходов, которые можно использовать. Полная характеристика всех алгоритмов оптимизации выходит далеко за рамки этой главы. Такого рода проблемам посвящено множество исследований в области исследования операций. Некоторые общие подходы к задачам оптимизации включают жадные алгоритмы [4], симулированный отжиг [5,6], табу-поиск [7,8], лагранжеву релаксацию [9].,10], искусственные нейронные сети [11,12] и обсуждаемые здесь эволюционные подходы. Ривз [2] содержит обзор некоторых из этих методов. В этой главе рассматриваются различные эволюционные подходы к задачам оптимизации. Затем мы представляем наши результаты по применению одного из этих подходов, генетических алгоритмов, к проблеме сборки последовательности ДНК. В разделе 2 представлены различные эволюционные подходы, применявшиеся к задачам оптимизации. В этом разделе основное внимание уделяется генетическим алгоритмам, поскольку здесь применяется именно этот метод. Раздел 3 описывает проблему сборки фрагментов ДНК в том виде, в каком мы ее охарактеризовали. Раздел 4 описывает процесс разработки генетического алгоритма, который мы использовали для задачи сборки фрагментов. В разделе 5 представлены некоторые модификации генетического алгоритма, которые могут решить определенные проблемы с выбранным конкретным генетическим алгоритмом. В разделе 6 мы описываем направление наших исследований в отношении генетических алгоритмов и анализа последовательностей ДНК. Просмотреть главуКнига покупок Прочитать главу полностью URL: https://www. G.E. McClearn, в International Encyclopedia of the Social & Behavioral Sciences, 2001 родители были смешаны, и влияние окружающей среды изменило смесь. Гениальный вклад Менделя состоял в том, что он постулировал существование дискретных элементов (позже названных «генами»). Дальнейшие постулаты, в сокращенной форме, заключались в том, что организм обладает многими такими генами, и каждый встречается в виде пары, причем организм получает по одному элементу от каждого родителя. Существуют различные формы генов (позже они будут названы «аллелями»). Точно так же, как индивидуум получил по одному аллелю для каждого гена от каждого родителя, он передает своему потомству только один аллель из каждой пары. Аллели не смешиваются и не модифицируются при ассоциации со своими партнерскими аллелями или с аллелями других генов, а передаются из организма в том же функциональном состоянии, в котором они были внесены в организм. В менделевской генетике проводится важное различие между фенотипом, измеряемым признаком и соответствующими генами или генотипом, и сутью теории является набор правил, определяющих отношения, которые могут возникнуть между генотипом и фенотипом. Менделевская генетика имеет дело с категориями. В идеальном случае всех людей можно однозначно отнести к одной из двух или трех категорий. Особая причина, по которой различие между генотипом и фенотипом важно, заключается в том, что отношения между ними не всегда аддитивны. Знание фенотипа не всегда однозначно информативно о генотипах. Например, в одной классической менделевской ситуации лица, обладающие двумя копиями определенного аллеля гена, относятся к категории А, обладающие двумя копиями другого аллеля того же гена относятся к категории В, а обладающие одной копией каждого гена также относятся к категории А. принадлежат к категории B. Другими словами, одна копия аллеля «для» категории B так же эффективна, как и две. Даже когда присутствует доминирование, в отношениях, конечно, присутствует дополнительный компонент. Таким образом, фенотипическую изменчивость можно рассматривать как за счет аддитивного компонента, так и за счет доминантных отклонений. Другая важная причина, по которой различие между генотипом и фенотипом имеет решающее значение, заключается в том, что гены не могут быть единственными детерминантами фенотипического исхода. Основной принцип множественных влияний, лежащий в основе количественной генетики, иллюстрируется на рис. Рисунок 1. Изображение менделевской частичной доминантной ситуации с влиянием окружающей среды, ответственным за фенотипическую изменчивость внутри генотипа. Источником генотипического разнообразия является, конечно же, наличие разных аллелей. Эти альтернативные формы гена возникают в результате мутационных событий, и их окончательная судьба в популяции зависит, среди прочего, от их влияния на репродуктивную «приспособленность» их обладателей (см. Falconer and Mackay 19).96, Хартл и Кларк, 1997). В любой популяции относительные частоты альтернативных аллелей (их может быть гораздо больше, чем два, но это обсуждение будет ограничено простым случаем двух) обычно далеко не равны. Таким образом, разные генотипы также не будут одинаково часты. Если p представляет собой частоту наиболее распространенного аллеля (скажем, A1), а q представляет наименее частый аллель (A2), и если спаривание происходит случайно, то относительные частоты генотипов A1A1, A1A2 и A2A2 будут p 2 , 2pq и q 2 соответственно. Таким образом, разные значения p и q, очевидно, приведут к разным частотам генотипов и, следовательно, к разным средним значениям и к разным дисперсиям. Просмотреть главуКнига покупки Прочитать главу полностью URL: https://www.sciencedirect.com/science/article/pii/B0080430767033751 Robert L. Chevalier, in Cellology 199 201 Binars in Cellology, 199 201 В то время как Современный синтез включал генетику в эволюционную теорию, эмбриология была практически исключена до конца 20-го века. Частично это могло быть результатом различения генотипа и фенотипа Йоханнсена, которое было преодолено с появлением эволюционной биологии развития (эво-дево). Британский эмбриолог Гэвин де Бир был исключением из этого разделения: в его книге Эмбриология и эволюция (1930) (расширено до Embryos and Ancestors в 1940 г.) он был одним из первых, кто постулировал роль отбора процессов развития в эволюции [53,54]. Раскол между биологией развития и эволюцией последовал за переходом от описательной эмбриологии в 19 веке к экспериментальной эмбриологии в 20 веке. Основа эво-дево была приписана Стивену Джею Гулду, биологу-эволюционисту из Гарварда, чья книга «Онтогения и филогения » (1977) защищала противоречивую концепцию де Бира о гетерохронии (эволюционных сдвигах во времени развития) как движущей силе эволюции. 55]. Открытие в 1983 г. генов гомеобокса привело к распознаванию факторов транскрипции, ответственных за морфогенетические программы, общие для животных, растений, грибов и одноклеточных эукариот. Такие «глубокие гомологии» между типами открыли множество новых направлений исследований в области развития почек, включая включение более широкого круга моделей животных и выяснение истории эволюции [56]. Ярким примером значимости глубокой гомологии для почечных заболеваний является эволюционная консервация генов цилиопатии, которая была показана для типов с общим происхождением, насчитывающим миллиард лет назад [57]. Просмотреть статью Прочтите полную статью URL: https://www.sciendirect.com/science/article/pii/s1084952117304305 Liane Gabora, В «Обзорах« Врачи жизни ». и будущие направления Учитывая нынешний ускоренный темп культурных изменений и их преобразующее воздействие на нас самих и нашу планету, становится необходимым получить четкое представление о том, как развивается культура. Поскольку культурная эволюция, как и биологическая эволюция, приводит к возникновению кумулятивной, открытой, адаптивной новизны, представляется целесообразным опираться на теорию биологической эволюции для объяснения культурной эволюции. Распространение взгляда Докинза на естественный отбор, взгляда, который не отражает основную алгоритмическую структуру процесса, привел к попыткам представить культурную эволюцию в дарвинистских терминах. Исследуя вопрос о том, являются ли элементы культуры репликаторами, мы увидели, что понятие репликаторов Докинза охватывает определенные аспекты самовоспроизведения, но игнорирует некоторые из наиболее фундаментальных принципов естественного отбора. К ним относятся: (1) код, который используется и как инструкции по самосборке, и как самоописание, (2) различие генотипа/фенотипа и (3) отсутствие передачи приобретенных характеристик, где (2) и (3) ) являются следствием (1). Предположение, что понятие репликатора отражает основные элементы естественного отбора, ошибочно привело к поддержке идеи о том, что культура развивается посредством селекционного процесса. Естественный отбор объясняет изменение частоты унаследованных признаков (признаков, передаваемых от одного поколения к другому посредством кода самосборки, такого как генетический код), а не приобретенных признаков (признаков, полученных между событиями передачи). Более поздние попытки представить культуру в эволюционных терминах, основанные на теориях происхождения жизни, основанных на общинном обмене, предполагают, что недарвинистские эпигенетические аспекты биологической эволюции могут быть полезны при разработке эволюционной основы культуры. . Это направление согласуется с возникающими усилиями (такими, как можно найти в [17]) по преодолению разрыва между эволюционными подходами к антропологии и археологии, с одной стороны, и интерпретативными и социальными конструкционистскими подходами, с другой. Исследования теории общественного обмена в рамках культурной эволюции сосредоточены на вычислительных моделях, которые включают эмпирические данные о том, как люди творчески адаптируют идеи к новым ситуациям. Parsons, Тел.: 1+407-823-5299 факс: 1+407-823-5419 , New Comprehensive Biochemistry, 9 010198 9 1 Введение
Таким образом, для задачи с 50 фрагментами оптимальному алгоритму потребовалось бы более 3570 лет, чтобы найти решение. Поскольку типовой проект сборки ружья может содержать более 10000 фрагментов [1], ясно, что точное решение невозможно. Вместо этого мы должны использовать приближенный алгоритм, который даст нам решение, которое почти всегда будет достаточно хорошим. 1.1 Проблемы оптимизации
Пространство поиска для задачи оптимизации — это просто набор всех возможных решений или ответов на вопрос. Функция пригодности — это мера, используемая для определения того, насколько хорош тот или иной ответ; поэтому функция пригодности определяет, какой ответ является лучшим. Таким образом, задача оптимизации становится задачей поиска в пространстве всех возможных ответов и нахождения наилучшего из них. sciencedirect.com/science/article/pii/S0167730608604664 1 Менделевская генетика
Точно так же факторы окружающей среды не изменяют природу элементов. Эти отношения описываются как доминирование; аллель, для которой одна копия столь же эффективна, как и две, является доминантным аллелем, а другой называется рецессивным аллелем. Два генотипа с одинаковыми аллелями называются гомозиготами, а генотип с одним аллелем — гетерозиготой. Возможны и другие отношения: в некоторых ситуациях связь между аллельной «дозой» и фенотипом является строго аддитивной, а для других фенотипов обнаруживаются промежуточные степени доминирования. 1. Фенотипические значения показаны для ряда особей каждого генотипа для одного гена с частично доминирующим функциональным отношением к фенотипу. Предполагается, что изменчивость внутри генотипа связана с обстоятельствами окружающей среды, которые уникальным образом повлияли на каждого человека. Проекция этих фенотипических значений на правую ось Y показывает распределение, изменчивость которого объясняется как генетическими различиями (с некоторым генетическим влиянием из-за аддитивного эффекта аллельной «замены» (представленной пунктирной линией регрессии ), а некоторые из-за доминирования (отклонения X, обозначающие средние значения каждой группы от линии регрессии), а также часть дисперсии из-за различий в окружающей среде между особями. Последствие, когда более одного гена могут влиять на фенотип будет рассмотрен ниже.0011 Фенотипические значения всех генотипов проецируются на единое распределение по правой оси Y 3.2 Эволюционная биология развития (evo-devo)
Первый был основан на изучении широкого круга видов по отношению к окружающей их среде, тогда как экспериментальная эмбриология стала ограничиваться несколькими модельными организмами в строго контролируемых условиях (рис. 1). Важно отметить, что отбор по признакам взрослых особей может управлять как вышестоящими, так и нижестоящими регуляторами генов Hox в развивающемся эмбрионе [58]. Остаются серьезные споры относительно относительного вклада мутаций в цис-регуляторных последовательностях плейотропных регуляторных локусов развития по сравнению со структурными мутациями [59,60]. Селекционная модель применима, когда передача приобретенных характеристик незначительна. Если процесс включает передачу приобретенных признаков и/или неслучайную изменчивость, эволюция во времени объясняется тем, что искажает или модифицирует изменчивость, а не естественным отбором9.0041, т.е. , это не связано со статистическим изменением частоты наследственных вариаций в поколениях из-за дифференциальной реакции на избирательное давление. Таким образом, попытки втиснуть культуру в дарвиновские рамки (даже те, которые на словах признают важность «богатой психологии») приводят к моделям, в которых «психология» сводится к источникам вариаций, которые возникают во время социальной передачи посредством таких процессов, как ошибка копирования. . Неспособность оценить дарвинистские подходы к культуре не проистекает из «тенденции мыслить категорически, а не количественно» (стр. 134), как Henrich et al. [61], и критики «не очень хорошо подготовлены для понимания математических моделей» (стр. 121). Критика этих моделей отражает подлинную ограниченность используемых методов и теоретического фундамента, на котором они основаны. Тем не менее дарвиновские подходы к культуре, такие как меметика и теория двойного наследования, внесли важный вклад в создание жизнеспособной научной основы для культуры, помогая определить, где именно нарушается аналогия между биологией и культурой.