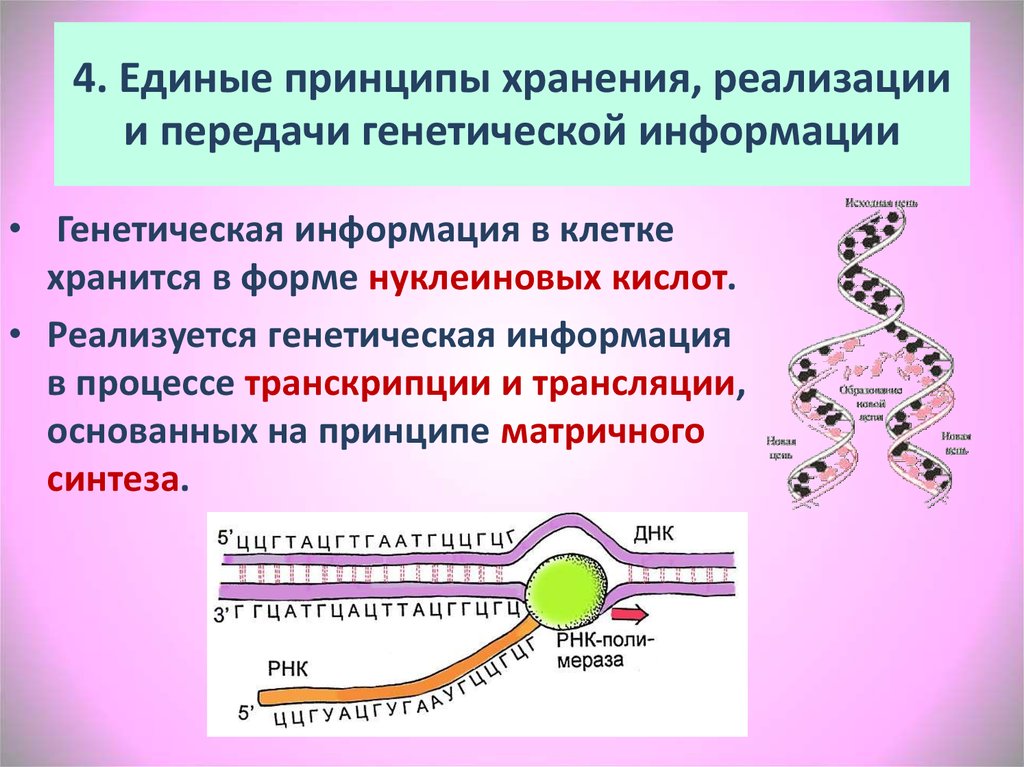
Генетический анализ: этапы развития, проблемы и перспективы
Генетическим анализом называют систему мероприятий, направленных на изучение механизмов генетической детерминации признаков. Среди них видное место отводится задачам формальной генетики, а именно формулированию и формализации моделей наследования и проверке генетических гипотез на конкретном эмпирическом материале. В мировой литературе этот раздел генетического анализа известен под названием segregation and linkage analysis. Именно о нем пойдет речь.
Как известно, генетика начиналась с генетического анализа, который был чрезвычайно популярен в первой половине века, являясь основным инструментом исследований. Затем популярность его падает, и новый подъем интереса к этому разделу генетики наблюдается лишь в последние годы. Как это ни парадоксально, но и потеря и новое приобретение популярности генетического анализа вызваны развитием молекулярно-генетических исследований. Причина первого понятна. Формальная генетика оперирует абстрактным понятием генов как неких дискретных факторов, линейно расположенных в группах сцепления и прямо или опосредованно влияющих на проявление признака. Формальная генетика не рассматривает проблем, связанных со структурой гена, с особенностями его функционирования, процессов, происходящих на пути от гена до признака. Именно поэтому в формальной генетике любое решение задачи имеет вероятностный, а не абсолютный характер. Результатом решения является формулировка модели наследования признака и указание степени ее правдоподобия. Молекулярная генетика, напротив, дает вполне конкретные ответы: она позволяет шаг за шагом проследить весь путь от гена до признака, выяснить, каким образом изменение нуклеотидного состава какого-либо участка ДНК приводит к полиморфизму признака. Очевидно, что знания, полученные методами молекулярной генетики, более однозначны и, следовательно, более ценны, чем результаты генетического анализа. Именно поэтому в 1960–1970-е годы постепенно формируется и закрепляется представление о том, что генетический анализ изжил себя и его следует рассматривать как один из архаизмов генетики.
Формальная генетика не рассматривает проблем, связанных со структурой гена, с особенностями его функционирования, процессов, происходящих на пути от гена до признака. Именно поэтому в формальной генетике любое решение задачи имеет вероятностный, а не абсолютный характер. Результатом решения является формулировка модели наследования признака и указание степени ее правдоподобия. Молекулярная генетика, напротив, дает вполне конкретные ответы: она позволяет шаг за шагом проследить весь путь от гена до признака, выяснить, каким образом изменение нуклеотидного состава какого-либо участка ДНК приводит к полиморфизму признака. Очевидно, что знания, полученные методами молекулярной генетики, более однозначны и, следовательно, более ценны, чем результаты генетического анализа. Именно поэтому в 1960–1970-е годы постепенно формируется и закрепляется представление о том, что генетический анализ изжил себя и его следует рассматривать как один из архаизмов генетики.
Это утверждение было бы справедливо, если бы не существовало широкого класса признаков, называемых сейчас комплексными (complex traits). Эти признаки проявляются на уровне целого организма, в их контроле задействовано большое число генетических и средовых факторов, часто взаимодействующих друг с другом. К комплексным признакам относится подавляющее большинство количественных признаков, характеризующих хозяйственно ценные свойства растений и животных, сюда относят многие распространенные наследственные болезни человека, такие, как гипертония, диабет, ишемическая болезнь сердца, некоторые формы онкологических заболеваний, многие психические расстройства. Очевидно, что, пользуясь только методами молекулярной генетики, нельзя разобраться в природе этих признаков. Прогресс может быть достигнут лишь при объединении усилий молекулярной и формальной генетики. Поскольку темпы развития генетического анализа существенно отставали от темпов развития молекулярной генетики, сейчас генетический анализ является лимитирующим звеном на пути прогресса генетики комплексных признаков. Именно поэтому в последние годы наблюдается заметное повышение интереса к формальной генетике: усиливается финансирование, организуются конференции, семинары и школы.
Эти признаки проявляются на уровне целого организма, в их контроле задействовано большое число генетических и средовых факторов, часто взаимодействующих друг с другом. К комплексным признакам относится подавляющее большинство количественных признаков, характеризующих хозяйственно ценные свойства растений и животных, сюда относят многие распространенные наследственные болезни человека, такие, как гипертония, диабет, ишемическая болезнь сердца, некоторые формы онкологических заболеваний, многие психические расстройства. Очевидно, что, пользуясь только методами молекулярной генетики, нельзя разобраться в природе этих признаков. Прогресс может быть достигнут лишь при объединении усилий молекулярной и формальной генетики. Поскольку темпы развития генетического анализа существенно отставали от темпов развития молекулярной генетики, сейчас генетический анализ является лимитирующим звеном на пути прогресса генетики комплексных признаков. Именно поэтому в последние годы наблюдается заметное повышение интереса к формальной генетике: усиливается финансирование, организуются конференции, семинары и школы. К сожалению, в нашей стране пока не ощущается больших перемен, и для многих генетиков термин «генетический анализ» ассоциируется с исследованиями, проводимыми в начале века. Чтобы продемонстрировать прогресс в этом направлении генетики, я попыталась проследить логику развития генетического анализа, эволюцию объектов исследования и предположений, в рамках которых велся анализ, а также указать те проблемы, которые необходимо решить, чтобы достичь прогресса в понимании природы комплексных признаков.
К сожалению, в нашей стране пока не ощущается больших перемен, и для многих генетиков термин «генетический анализ» ассоциируется с исследованиями, проводимыми в начале века. Чтобы продемонстрировать прогресс в этом направлении генетики, я попыталась проследить логику развития генетического анализа, эволюцию объектов исследования и предположений, в рамках которых велся анализ, а также указать те проблемы, которые необходимо решить, чтобы достичь прогресса в понимании природы комплексных признаков.
В истории развития генетического анализа можно выделить три основных этапа, отличающихся друг от друга объектом исследования, эмпирическим материалом, тестируемыми генетическими гипотезами и общими предположениями, в рамках которых ведется анализ. Объектом исследования этого раздела генетики является признак, который может быть формально представлен как бинарная, качественная или количественная характеристика. Первый этап генетического анализа называют обычно классическим генетическим анализом, он базируется на гибридологическом эксперименте и строится на двух предположениях: 1) исходные родительские линии гомозиготны по локусам, принимающим участие в контроле признака; 2)главное место в контроле признака отводится майоргенам, полигенная и средовая компоненты пренебрежимо малы.
Из первого предположения следует, что обе родительские формы и F1 являются генетически однородными группами, в которых все особи абсолютно идентичны по генам, принимающим участие в формировании изучаемого признака. Следовательно, потомки всех однотипных скрещиваний между представителями этих трех групп (P1, P2 и F1) могут быть объединены без какой-либо потери информации. Из второго предположения следует, что все особи с одинаковым генотипом фенотипически неразличимы. Следовательно, сложность формализованного представления признака ограничена числом генотипов. Именно поэтому объектом исследования на этом этапе являются бинарные признаки.
Таким образом, в рамках сделанных предположений эмпирический материал может быть представлен небольшим числом групп большой численности. Задача заключается в том, чтобы проверить, соответствуют ли эмпирические данные сформулированной модели наследования этого признака. На первом этапе модели наследования максимально просты – это рецессивная или доминантная модели. Единственным параметром интереса является сегрегационная частота, определенная как вероятность родителей с определенными генотипами произвести потомка с заданным фенотипом. Ожидаемая сегрегационная частота определяется типом скрещивания и моделью наследования. Генетические гипотезы проверяются одним из двух статистических методов: либо подсчитываются ожидаемые численности различных фенотипических классов и сравниваются с наблюдаемыми, либо оценивается сегрегационная частота и сравнивается с ожидаемой.
Единственным параметром интереса является сегрегационная частота, определенная как вероятность родителей с определенными генотипами произвести потомка с заданным фенотипом. Ожидаемая сегрегационная частота определяется типом скрещивания и моделью наследования. Генетические гипотезы проверяются одним из двух статистических методов: либо подсчитываются ожидаемые численности различных фенотипических классов и сравниваются с наблюдаемыми, либо оценивается сегрегационная частота и сравнивается с ожидаемой.
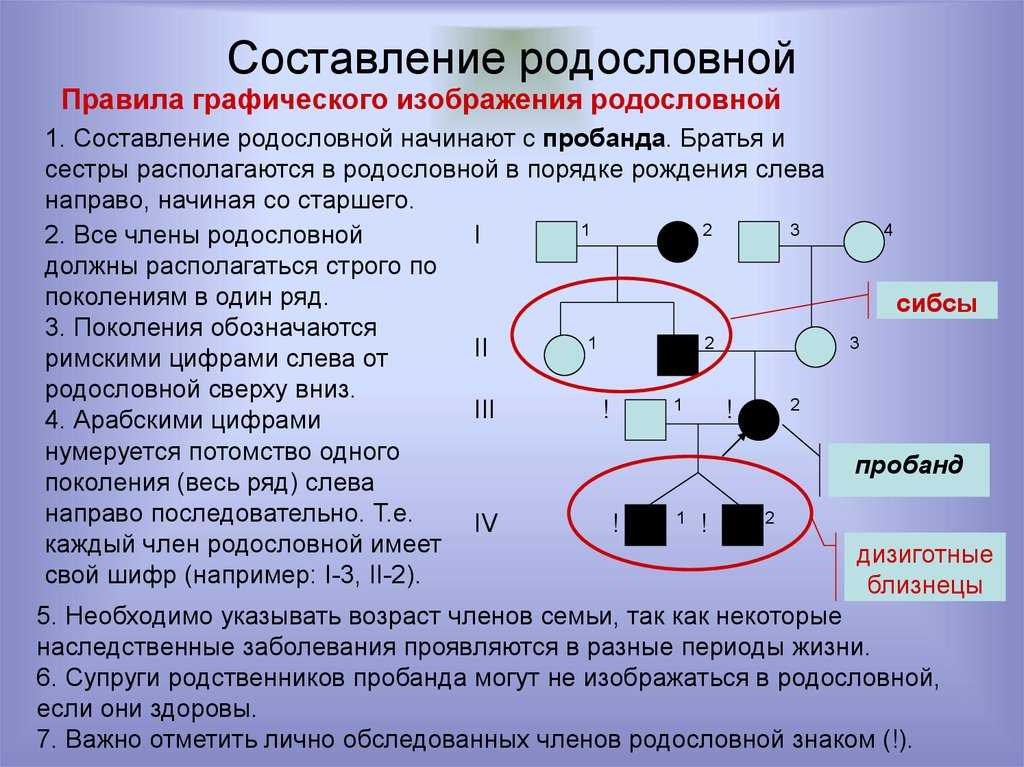
Эволюция метода на первом этапе заключается главным образом в усложнении моделей за счет увеличения числа локусов, принимающих участие в контроле признака, и за счет рассмотрения различных типов взаимодействия генов. Второй этап развития сегрегационного анализа характеризуется сменой объекта исследования, переходом на анализ признаков человека. У человека, как известно, существуют проблемы с получением чистых линий и принудительными скрещиваниями. Поэтому здесь нельзя использовать методы классического генетического анализа. К счастью, человек обладает одним замечательным свойством, не присущим животным и тем более растениям, – большинство людей помнят своих предков, а это значит, что можно строить и реконструировать родословные и выбирать из популяции те из них, которые генетику хотелось бы получить в эксперименте. На втором этапе материал максимально приближен к гибридологическому эксперименту, он представлен потомками родителей с известными генотипами. По своей идеологии этот этап очень близок первому. По-прежнему анализируют бинарные признаки, по прежнему тестируют доминантные или рецессивные модели. Предположения почти идентичны тем, что были на первом этапе: предполагается, что генотипы родителей известны и признак контролируется главным образом майоргенами. Разница заключается в том, что эмпирический материал берется из популяции, а не из гибридологического эксперимента. Каким образом выбираются из популяции скрещивания с известными генотипами? При доминантном наследовании наиболее информативными являются скрещивания особей с нормальным гомозиготным и с гетерозиготным генотипами.
К счастью, человек обладает одним замечательным свойством, не присущим животным и тем более растениям, – большинство людей помнят своих предков, а это значит, что можно строить и реконструировать родословные и выбирать из популяции те из них, которые генетику хотелось бы получить в эксперименте. На втором этапе материал максимально приближен к гибридологическому эксперименту, он представлен потомками родителей с известными генотипами. По своей идеологии этот этап очень близок первому. По-прежнему анализируют бинарные признаки, по прежнему тестируют доминантные или рецессивные модели. Предположения почти идентичны тем, что были на первом этапе: предполагается, что генотипы родителей известны и признак контролируется главным образом майоргенами. Разница заключается в том, что эмпирический материал берется из популяции, а не из гибридологического эксперимента. Каким образом выбираются из популяции скрещивания с известными генотипами? При доминантном наследовании наиболее информативными являются скрещивания особей с нормальным гомозиготным и с гетерозиготным генотипами. Их легко выбрать из популяции, если мутантный аллель встречается с низкой частотой – практически все родительские пары, в которых один из супругов болен, а другой здоров, будут иметь требуемые генотипы. Потомки всех таких скрещиваний объединяются, и проводится анализ точно так же, как на первом этапе. Дополнительно используется проверка того, что распределение семей по числу больных потомков подчиняется биномиальному распределению.
Их легко выбрать из популяции, если мутантный аллель встречается с низкой частотой – практически все родительские пары, в которых один из супругов болен, а другой здоров, будут иметь требуемые генотипы. Потомки всех таких скрещиваний объединяются, и проводится анализ точно так же, как на первом этапе. Дополнительно используется проверка того, что распределение семей по числу больных потомков подчиняется биномиальному распределению.
При рецессивном контроле признака информативным является скрещивание двух гетерозигот. Популяционным аналогом этого скрещивания являются семьи, в которых оба родителя здоровы, а хотя бы один ребенок болен. Очевидно, что при таком условии в выборку никогда не попадут те семьи, где у гетерозиготных родителей чисто случайно не появилось ни одного больного ребенка. Поэтому полученные обычным образом оценки сегрегационных частот оказываются завышенными. Для коррекции этого смещения необходимо отказаться от объединения потомков аналогичных скрещиваний и рассматривать индивидуальные семьи.
Так же, как на первом этапе, здесь тестируются наиболее простые модели доминантного и рецессивного контроля признака. Развитие метода идет по трем направлениям. Во-первых, так же, как на первом этапе, усложняются модели, но эти усложнения, как правило, лежат в рамках моногенных диаллельных моделей. Во-вторых, происходит мощнейшая разработка так называемой ascertainment problem – проблемы коррекции смещения, вызванного неслучайностью выбора: формулируется понятие пробанда и разрабатываются основные принципы решения проблемы. И в-третьих, отказ от объединения потомков и затем введение дополнительного параметра (вероятности выбора пробанда) приводят к заметному усложнению оценки сегрегационной частоты. Возникает необходимость разработки специальных алгоритмов получения оценок, разработки специальных таблиц и формул, ведь это еще докомпьютерная эра и все вычисления делаются вручную.
Значительно усложняется анализ сцепления. Это уже не гибридологический эксперимент, и фаза сцепления генов у дигетерозиготного родителя, как правило, неизвестна. При этом уменьшается информативность данных и накладываются более жесткие требования к точности анализа. Именно в эти годы появляются две принципиально важные для анализа сцепления разработки: предлагается непараметрический метод тестирования сцепления, основанный на анализе идентичности генов по происхождению, и предлагается специальный критерий Lod score, благодаря которому был достигнут огромный прогресс в локализации генов, ответственных за менделевские признаки.
При этом уменьшается информативность данных и накладываются более жесткие требования к точности анализа. Именно в эти годы появляются две принципиально важные для анализа сцепления разработки: предлагается непараметрический метод тестирования сцепления, основанный на анализе идентичности генов по происхождению, и предлагается специальный критерий Lod score, благодаря которому был достигнут огромный прогресс в локализации генов, ответственных за менделевские признаки.
Тем не менее, второй этап развития генетического анализа – это анализ простых признаков и простых моделей. Лимитирующим здесь оказывается предположение об известных генотипах родителей. В самом деле, только при рецессивной или доминантной модели можно выбрать из популяции семьи, удовлетворяющие этому условию. Получается замкнутый круг – высказывается предположение о модели наследования, при ее справедливости устанавливаются генотипы родителей, а затем по фенотипам потомков этих родителей тестируется выбранная модель.
Выход из этой ситуации наметился лишь в 1970 г., когда было снято предположение о том, что генотипы родителей известны. Сначала было предложено рассматривать распределение фенотипов не только потомков, но и родителей, проводя суммирование по всем возможным генотипам родителей. А буквально на следующий год, в 1971 г., были сформулированы основы того, что сейчас называется комплексным сегрегационным анализом. Были введены три математические компоненты, описывающие распределение генотипов в популяции, правило передачи аллелей из поколения в поколение и правило построения фенотипа на заданном генотипе. Было показано, что в терминах этих трех распределений можно записать вероятность совместного распределения признаков у членов родословной. Родословная эта может иметь произвольную структуру, признак может быть формализован любым способом, в том числе быть количественным, и модели наследования могут не ограничиваться простыми рецессивными или доминантными отношениями. Кроме того, вычленение в отдельную компоненту вероятностей передачи генов от родителей потомкам позволяет ввести специальные критерии для проверки менделевской сегрегации генов.
Развитие метода продолжается главным образом по двум направлениям: 1) Создаются новые более сложные модели наследования. Прежде всего в модель в явном виде вводится майоргенная, полигенная и средовая компоненты. Появляется возможность учесть общие внутрисемейные эффекты, фенотипические корреляции между родственниками, наличие факторов риска развития болезни, разный возраст проявления болезни и т.д. 2) Анализ родословных произвольной структуры возможен лишь с помощью вычислительной техники, поэтому отдельное направление посвящено созданию алгоритмов и пакетов программ для генетического анализа.
Как видно, на третьем этапе генетический анализ выходит за рамки ограничений, характерных для классического генетического анализа, доступными для анализа становятся количественные признаки, модели наследования включают в себя различные генетические и средовые факторы. Теоретически проблема анализа комплексных признаков может считаться решенной. К сожалению, на практике все не так просто.
Как отмечалось выше, был предложен специальный критерий для проверки менделевской сегрегации генов. В основе его лежит сравнение трех гипотез, отличающихся друг от друга предположениями о переходных вероятностях. Менделевская гипотеза фиксирует те значения этих вероятностей, которые соответствуют менделевским правилам. Общая, или неограниченная, модель не накладывает никаких ограничений на эти параметры, и средовая модель предполагает их равными. Менделевская сегрегация генов считается установленной, когда первая и вторая гипотезы не отличаются друг от друга, а вторая и третья значимо отличимы. В специальном генетико-стохастическом эксперименте было показано, что использование этого комплексного критерия препятствует ошибочному установлению менделизма. Однако, если майоргенный контроль не установлен, не обязательно, что его нет на самом деле. Причиной ложного отвержения менделизма чаще всего бывает генетическая гетерогенность, ошибки диагностики или некорректная формализация количественного признака, а также неадекватность модели наследования. В настоящее время существует множество подходов к решению этой проблемы: создаются критерии гетерогенности выборки, методы трансформации количественных данных, расширяются и усложняются модели наследования признаков. Однако основная проблема комплексных признаков заключается в том, что они настолько сложны и многообразны, что очень трудно, если не невозможно, описать их наследование единой функцией так, чтобы, варьируя значения параметров, можно было формализовать любую ситуацию. Даже если бы это удалось, число оцениваемых параметров было бы настолько велико, что вряд ли удалось бы собрать столько эмпирических данных, чтобы оценить эти параметры. Поэтому при анализе комплексных признаков приходится вести большую предварительную работу, изучая половые и возрастные особенности их проявления, выделяя модифицирующие факторы и формируя модель наследования, наиболее полно отражающую специфику анализируемого признака. Пока нет и речи о том, чтобы сделать анализ комплексных признаков автоматическим.
В настоящее время существует множество подходов к решению этой проблемы: создаются критерии гетерогенности выборки, методы трансформации количественных данных, расширяются и усложняются модели наследования признаков. Однако основная проблема комплексных признаков заключается в том, что они настолько сложны и многообразны, что очень трудно, если не невозможно, описать их наследование единой функцией так, чтобы, варьируя значения параметров, можно было формализовать любую ситуацию. Даже если бы это удалось, число оцениваемых параметров было бы настолько велико, что вряд ли удалось бы собрать столько эмпирических данных, чтобы оценить эти параметры. Поэтому при анализе комплексных признаков приходится вести большую предварительную работу, изучая половые и возрастные особенности их проявления, выделяя модифицирующие факторы и формируя модель наследования, наиболее полно отражающую специфику анализируемого признака. Пока нет и речи о том, чтобы сделать анализ комплексных признаков автоматическим.
Та же ситуация наблюдается и для анализа сцепления. Прекрасно зарекомендовавший себя и обеспечивший огромный прогресс в картировании менделевских признаков критерий Lod score предполагает, что единственным оцениваемым параметром является коэффициент рекомбинации, а все параметры модели наследования известны. Пока есть проблемы с установлением модели наследования, анализ сцепления также не может быть переведен на автоматический уровень. Обычно используются два методических подхода: анализ генов-кандидатов и сканирование генома. Первый подход предполагает, что известны те ключевые звенья в цепи биохимических реакций, которые в принципе могут повлиять на проявление признака. Тестирование генов, ответственных за эти звенья, на причастность к контролю анализируемого признака осуществляется обычно непараметрическими методами анализа. Они хороши тем, что не требуют задания модели наследования, однако недостатком их является пониженная мощность. Другой путь – сканирование генома – заключается в том, что все хромосомы насыщаются анонимными маркерами, и предполагаемый ген, ответственный за развитие признака, «зажимается» между двумя соседними маркерами. Из-за невысокой информативности данных установить можно только тесное сцепление. Но для этого надо, чтобы маркеров было много и они плотно покрывали хромосому. Однако чем больше маркеров, тем больше повторных сравнений и тем выше уровень значимости критериев. Возможно, что именно из-за этих недостатков используемых подходов так редко удается добиться успешного картирования генов, ответственных за развитие комплексных признаков.
Из-за невысокой информативности данных установить можно только тесное сцепление. Но для этого надо, чтобы маркеров было много и они плотно покрывали хромосому. Однако чем больше маркеров, тем больше повторных сравнений и тем выше уровень значимости критериев. Возможно, что именно из-за этих недостатков используемых подходов так редко удается добиться успешного картирования генов, ответственных за развитие комплексных признаков.
Сейчас развитие методов генетического анализа идет по пути совершенствования анализа сцепления. Большие надежды возлагаются на многоточечное картирование и на новые подходы, объединяющие параметрический и непараметрический методы, сегрегационный и рекомбинационный анализ.
Таким образом, несмотря на большой прогресс генетического анализа, мы еще весьма далеки от того, чтобы довести анализ комплексных признаков до автоматизма, который в свое время обеспечил прорыв в картировании и идентификации генов, ответственных за развитие менделевских признаков. К сожалению, ощущается явная нехватка специалистов в этой области, поскольку в мире насчитывается от силы два десятка лабораторий, где все эти годы продолжались разработки методов генетического анализа. Наш сектор является единственным в России центром, где ведутся такие исследования.
Наш сектор является единственным в России центром, где ведутся такие исследования.
Т.И. Аксенович, Информационный вестник ВОГиС, 1999, №10.
Источник: Ген Эксперт
Искусственный интеллект. Генетический алгоритм и его применения / Offсянка
🇷🇺🇰🇿
Сегодня 09 февраля 2023
О сайте Реклама Рассылка Контакты
3DNews Offсянка Искусственный интеллект. Генетический ал… Самое интересное в новостях Генетический алгоритм: принцип естественного отбора как инструмент искусственного интеллекта. Хотите узнать, как это работает? Сегодняшняя статья — об одном из самых применяемых алгоритмов поиска ⇣ Содержание „Выживает не самый сильный и не самый умный, а тот, кто лучше всех приспосабливается к изменениям.“ “Даже если у одного прочитавшего из тысячи появится интерес к этим алгоритмам, и он решит углубиться — это уже отлично. Внимание, вопрос: помните ли вы движущие силы эволюции по Дарвину? Эти знания нужны для понимания работы генетического алгоритма, так как он пытается имитировать процессы, действительно происходящие в природе. Итак, основной предпосылкой эволюции является наследственная изменчивость, а её движущими силами — борьба за существование и естественный отбор. Используя генетический алгоритм, вы действуете по сути как творец и сами устанавливаете законы эволюции, позволяющие достичь оптимальности в популяции. В идеале должны выработаться необходимые для выживания и адаптации качества, которые и будут искомым решением. Посмотрим, как реализуются эти принципы в генетическом алгоритме. ⇡#Что это?Генетический алгоритм (ГА) — это алгоритм поиска и оптимизации, прообразом которого стал биологический принцип естественного отбора. ⇡#Как он работает?Первый этап — создание популяции. В данном случае популяция — это не совокупность биологических особей, а множество возможных решений имеющейся проблемы, которые образуют пространство поиска (space search). Второй этап — подсчёт функции пригодности (приспособленности, fitness function). Данная функция принимает на вводе потенциальное решение проблемы (candidate solution), а выдаёт значение, оценивающее его пригодность. В случае с классическим генетическим алгоритмом целевая функция и функция пригодности — это одно и то же. Далее проверим, выполнено ли условие остановки алгоритма. Алгоритм прекратит исполняться, если достигнуто ожидаемое оптимальное значение, если полученное значение больше не улучшается или по истечении заданного времени/количества итераций. После остановки происходит выбор самой приспособленной хромосомы (по наибольшему значению функции). Если же условие остановки не выполнено, то по результатам естественного отбора будет производиться селекция хромосом для производства потомков. Третий этап – Блок-схема генетического алгоритма Скрещивание, мутация и селекция – это генетические операторы. Одноточечный кроссинговер (single-point crossover): есть пара родительских хромосом с набором генов L, для них случайным образом выбирается так называемая точка скрещивания Px – это некая позиция гена в хромосоме. Одноточечный кроссинговер Двухточечный кроссинговер (two-point crossover): обмен генетическим материалом, (то есть, битами) происходит в двух точках скрещивания. Двухточечный кргоссинговер Однородный кроссинговер (uniform crossover): значение каждого бита в хромосоме потомка определяется согласно случайно сгенерированной маске кроссинговера. Если в маске стоит 0 – берется ген от первого родителя, если 1 – от второго. Однородный кроссинговер Для более глубокого погружения в тему эволюционных алгоритмов реокмендуем изучить статью F.Herrera, M.Losano, A.M.Sanches Hybrid Crossover Operators for Real-Coded Genetic Algorithms: An Experimental Study. Мутация — генетический оператор, который с некой вероятностью меняет один или несколько «генов» в случайных позициях «хромосомы». Плюсы ГА Этот алгоритм имеет уникальные сильные стороны:
Минусы ГА
⇡#Для каких задач используется ГА?С помощью ГА решается целый спектр задач: от разработки антенн NASA до программ распознавания структуры белков. В финансах ГА успешно используется для моделирования экономических агентов с ограниченно рациональным поведением: финансового прогнозирования, инвестирования, и т. д. Одна из интересных задач — оптимизация финансового портфеля (portfolio optimization). В теории игр ГА применяется, например, для разрешения дилеммы узника. Его можно применять в играх с двумя участниками для оптимизации стратегий. В робототехнике ГА прекрасно применяется для управления человекоподобным роботом, оптимизации планирования маршрута (routing). В авиации — для системы контроля полетов. Кстати об авиации: ученые General Electric и Ренсселеровского политехнического института применили ГА для конструирования турбины реактивного двигателя, который применяется в современных авиалайнерах. Можно использовать ГА и в более близких для нас ситуациях, например, для планирования расписания на производстве или в крупном учебном заведении. В первом случае фитнес-функция может задавать количество деталей, изготовленных за определенное количество времени, а во втором – «штрафовать» конфликтующие ветки расписания. Что дальше? Читатель, который хочет знать больше, может обратиться к материалам GECCO conference — это конференция, которая посвящена эволюционному программированию, там самые горячие новости и обновления. Математическая сторона хорошо описана здесь: https://loginom.ru/blog/ga-math Ещё по применениям ГА на английском языке: https://www.brainz.org/15-real-world-applications-genetic-algorithms/ Недавно вышла привлекающая внимание книга: Buontempo, Frances. Genetic algorithms and machine learning for programmers: create AI models and evolve solutions. Pragmatic Bookshelf, 2019. Материалы, которые использовались для написания этой статьи:
Другие материалы цикла:
⇣ Содержание
Если Вы заметили ошибку — выделите ее мышью и нажмите CTRL+ENTER. Материалы по теме Постоянный URL: https://3dnews.ru/1050009/iskusstvenniy-intellekt-geneticheskiy-algoritm-i-ego-primeneniya Теги: исследование, генетика, ии ⇣ Комментарии |


 Как и в природе, вероятность скрещивания на несколько порядков выше вероятности мутации. Скрещивание поддерживает бесконечное разнообразие в популяции, это перераспределение генетического материала родителей, благодаря которому у потомков появляются новые сочетания генов. Но о каких «генах» и «хромосомах» может идти речь вне контекста живой природы? В генетическом алгоритме «хромосома» — набор параметров, определяющих предлагаемое возможное решение, а «ген» — это одна из «букв» строки «хромосомы», как правило, имеющая двоичное значение. Как мы помним, в результате селекции отбираются самые приспособленные хромосомы — к ним и применяются эти генетические операторы. Наверное, вы сейчас думаете, каким же образом может происходить скрещивание у таких «хромосом». Возможно несколько сценариев, упомянем лишь некоторые из них.
Как и в природе, вероятность скрещивания на несколько порядков выше вероятности мутации. Скрещивание поддерживает бесконечное разнообразие в популяции, это перераспределение генетического материала родителей, благодаря которому у потомков появляются новые сочетания генов. Но о каких «генах» и «хромосомах» может идти речь вне контекста живой природы? В генетическом алгоритме «хромосома» — набор параметров, определяющих предлагаемое возможное решение, а «ген» — это одна из «букв» строки «хромосомы», как правило, имеющая двоичное значение. Как мы помним, в результате селекции отбираются самые приспособленные хромосомы — к ним и применяются эти генетические операторы. Наверное, вы сейчас думаете, каким же образом может происходить скрещивание у таких «хромосом». Возможно несколько сценариев, упомянем лишь некоторые из них. К [1; Px] одной родительской хромосомы присоединяется [Px+1; L] другой, и получается первый потомок. Второй потомок получается также скрещиванием, но «в обратную сторону».
К [1; Px] одной родительской хромосомы присоединяется [Px+1; L] другой, и получается первый потомок. Второй потомок получается также скрещиванием, но «в обратную сторону». Для чего он нужен? Зачем мутации (изменения в генетическом коде) существуют в природе, могут ли они способствовать лучшей выживаемости вида? Эта статья не о генетике, но не будем забывать, что именно она послужила источником вдохновения для Холланда, создателя генетического алгоритма (1975). Потомки, которые подверглись воздействию генетических операторов, образуют новую популяцию – и в ней начинается очередная итерация ГА. Снова идет подсчет функции пригодности, происходит естественный отбор, а дальше алгоритм либо остановится, если заданное условие выполнено, либо вновь перейдет к селекции. Если хочется посмотреть интересное применение, можно почитать разбор задачи коммивояжера (travelling salesman problem) и задачи об укладке рюкзака (knapsack problem) с применением ГА. Обе эти задачи являются задачами комбинаторной оптимизации, то есть в конечном множестве объектов мы ищем оптимальный. Сами того не подозревая, мы решаем подобные задачи каждый день. Теперь посмотрим на преимущества и недостатки этого метода.
Для чего он нужен? Зачем мутации (изменения в генетическом коде) существуют в природе, могут ли они способствовать лучшей выживаемости вида? Эта статья не о генетике, но не будем забывать, что именно она послужила источником вдохновения для Холланда, создателя генетического алгоритма (1975). Потомки, которые подверглись воздействию генетических операторов, образуют новую популяцию – и в ней начинается очередная итерация ГА. Снова идет подсчет функции пригодности, происходит естественный отбор, а дальше алгоритм либо остановится, если заданное условие выполнено, либо вновь перейдет к селекции. Если хочется посмотреть интересное применение, можно почитать разбор задачи коммивояжера (travelling salesman problem) и задачи об укладке рюкзака (knapsack problem) с применением ГА. Обе эти задачи являются задачами комбинаторной оптимизации, то есть в конечном множестве объектов мы ищем оптимальный. Сами того не подозревая, мы решаем подобные задачи каждый день. Теперь посмотрим на преимущества и недостатки этого метода.




Основные принципы генетики: Генетика Менделя
Основные принципы генетики: Генетика МенделяМенделя Генетика
Гибриды домашних лошадей
Для тысяч лет фермеры и скотоводы селекционно разводят свои растения и животных производить больше полезного гибриды . Это был своего рода случайный процесс, поскольку фактические механизмы, управляющие наследованием, были неизвестны.Знание из этих генетических механизмов, наконец, появились в результате тщательной лабораторной селекции. эксперименты, проведенные за последние полтора века.
Грегор Мендель
1822-1884По В 1890-х годах изобретение более совершенных микроскопов позволило биологам обнаружить основные факты клеточного деления и полового размножения. Генетика в центре внимания исследования затем перешли к пониманию того, что на самом деле происходит при передаче наследственных признаков от родителей к детям. Для объяснения наследственности было предложено несколько гипотез, но Грегор Мендель , малоизвестный среднеевропейский монах, был единственным, кто понял это более или менее меньше прав. Его идеи были опубликованы в 1866 году, но в значительной степени остались непризнанными.
Горох обыкновенный
Пока Исследования Менделя касались растений, основных принципов наследственности. которые он обнаружил, также применимы к людям и другим животных, потому что механизмы наследственности в основном одинаковы для всех сложных живых организмов. формы.
Через селективное скрещивание растений гороха обыкновенного ( Pisum sativum ) на протяжении многих поколений, Мендель обнаружил, что определенные черты проявляются у потомства без какого-либо смешения родительские характеристики.
1. цвет цветка фиолетовый или белый 5. цвет семянжелтый или зеленый 2. расположение цветка пазушное или терминал 6. форма стручка надута или суженный 3. длина штока длинная или короткая 7. цвет стручка желтый или зеленый 4. форма семян круглая или морщинистый Это наблюдение, что эти признаки не проявляются у потомков растений с промежуточными формами было критически важно, потому что ведущая теория в биологии в было время, когда унаследованные черты смешивались из поколения в поколение. Большинство ведущих ученых XIX в.век принял эту «теорию смешения». Чарльз Дарвин предложил другая столь же неверная теория, известная как «пангенезис». . Это утверждало, что наследственные «частицы» в наших телах подвержены влиянию вещей. мы делаем в течение нашей жизни. Считалось, что эти модифицированные частицы мигрируют через кровь к половым клеткам и впоследствии может быть унаследована следующим поколение. По сути, это была вариация неверного представления Ламарка о «наследование приобретенных признаков».
Мендель выбрал обычные садовые растения гороха в центре его исследований, потому что они могут быть легко выращиваются в больших количествах, и их воспроизводством можно управлять. Растения гороха имеют как мужских, так и женских половых органов. В результате они могут либо самоопыляться или перекрестно опыляться с другим растением. В своих экспериментах, Менделю удалось избирательно перекрестно опылить чистопородных растения с определенными признаками и наблюдайте за результатом на протяжении многих поколений. Это послужило основой для его выводов о природе генетическое наследование.
Репродуктивная
конструкции
цветы
 Знание
из этих генетических механизмов, наконец, появились в результате тщательной лабораторной селекции.
эксперименты, проведенные за последние полтора века.
Знание
из этих генетических механизмов, наконец, появились в результате тщательной лабораторной селекции.
эксперименты, проведенные за последние полтора века. до 1900, что было намного позже его смерти. Его ранняя взрослая жизнь прошла в относительной безвестности.
проведение фундаментальных генетических исследований и преподавание
математика, физика и греческий язык в средней школе в Брно (сейчас в Чехии). В
В последние годы он стал настоятелем своего монастыря и оставил
научная работа.
до 1900, что было намного позже его смерти. Его ранняя взрослая жизнь прошла в относительной безвестности.
проведение фундаментальных генетических исследований и преподавание
математика, физика и греческий язык в средней школе в Брно (сейчас в Чехии). В
В последние годы он стал настоятелем своего монастыря и оставил
научная работа. Например, цветки гороха либо фиолетовые, либо
белый — промежуточные окраски не появляются в потомстве перекрестноопыляемых растений гороха. Мендель наблюдал
семь признаков, которые легко распознаются и, по-видимому, встречаются только в одной из двух форм:
Например, цветки гороха либо фиолетовые, либо
белый — промежуточные окраски не появляются в потомстве перекрестноопыляемых растений гороха. Мендель наблюдал
семь признаков, которые легко распознаются и, по-видимому, встречаются только в одной из двух форм:

В перекрестноопыляемые растения, дающие либо желтые, либо зеленые семена гороха исключительно, Мендель обнаружил, что первое потомство поколения (f1) всегда имеет желтые семена.
 Однако
следующее поколение (f2) постоянно имеет соотношение желтого к зеленому 3: 1.
Однако
следующее поколение (f2) постоянно имеет соотношение желтого к зеленому 3: 1.
Это соотношение 3:1 встречается в более поздних поколениях как хорошо. Мендель понял, что эта лежащая в основе регулярность была ключом к понимание основных механизмов наследования.
Он пришел к трем важным выводам из эти экспериментальные результаты:
1. что наследство каждая черта определяется «единицами» или «факторами», которые передаются потомкам в неизменном виде. (эти единицы теперь называются генами ) 2. , что физическое лицо наследует один такая единица от каждого родителя для каждого признака 3. , что черта может не отображаться в индивидуально, но может передаваться следующему поколению. Важно понимать, что в этом эксперименте исходные родительские растения были гомозиготными для цвет семян гороха. То есть каждый из них имел две одинаковые формы (или аллели ) принадлежащий ген этого признака — 2 желтых или 2 зеленых. Все растения поколения f1 были гетерозиготный . Другими словами, они каждый унаследовал два разных аллеля — по одному от каждого родительского растения. Это становится яснее, когда мы смотрим на фактический генетический состав, или генотип гороха, а не только фенотип , или наблюдаемые физические характеристики.
Обратите внимание, что каждое из растений поколения f1 (показано выше) унаследовали аллель Y от одного родителя и аллель G от другого.
Со всеми семь растений гороха признаков, изученных Менделем, одна форма оказалась доминирующей над другой, то есть он маскировал присутствие другого аллеля. Например, когда генотип по окраске семян гороха – YG (гетерозигота), фенотип – желтый. Однако доминантный желтый аллель не изменяет рецессивный зеленый в любом случае. Оба аллеля могут передаваться следующему поколению. без изменений.
Наблюдения Менделя из эти эксперименты можно свести к двум принципам:
1. принцип разделения 2. принцип независимой Ассортимент Согласно принцип сегрегация , для любого конкретного признака пара аллелей каждого родителя разделяется и только один аллель переходит от каждого родителя к потомству.
Разделение аллели в продукции половых клеток
 Когда растения f1 размножаются, каждое из них имеет
равные шансы передачи аллелей Y или G каждому
потомство.
Когда растения f1 размножаются, каждое из них имеет
равные шансы передачи аллелей Y или G каждому
потомство. Какой аллель в
пара аллелей родителей передается по наследству случайно. Теперь мы знаем, что это
сегрегация аллелей происходит в процессе образования половых клеток
(то есть мейоз
).
Какой аллель в
пара аллелей родителей передается по наследству случайно. Теперь мы знаем, что это
сегрегация аллелей происходит в процессе образования половых клеток
(то есть мейоз
).По принципу независимого ассортимента потомству передаются разные пары аллелей независимо друг от друга. В результате появляются новые комбинации генов в ни один из родителей не возможен. Например, горох унаследовал способность появление пурпурных цветов вместо белых не делает более вероятным, что это воля также наследуют способность производить желтые семена гороха в отличие от зеленых. Так же, принцип независимого ассортимента объясняет, почему человеческая наследственность того или иного цвет глаз не увеличивает и не уменьшает вероятность наличия 6 пальцев на каждой руке.
Эти два принципа наследование, наряду с пониманием единиц наследование и господство, были начала нашей современной науки генетики. Однако Мендель этого не понимал. есть исключения из этих правил. Некоторые из этих исключений будут рассмотрены в третий раздел этого руководства и в Синтетическая теория эволюции руководство.
Сосредоточившись на Менделе как на отце генетики, современная биология часто забывает, что его экспериментальные результаты также опровергли Теория Ламарка о наследовании приобретенных признаков, описанная в Ранние теории учебник по эволюции. Менделю редко приписывают это, потому что его работа оставалась по существу неизвестной до тех пор, пока идеи Ламарка не были широко отвергается как маловероятное.
ПРИМЕЧАНИЕ: Некоторые биологи называют «принципы» Менделя «законами».
ПРИМЕЧАНИЕ. Одна из причин, по которой Мендель провел его селекционные эксперименты с растениями гороха заключались в том, что он мог наблюдать наследование формирует до двух поколений в год. Сегодня генетики обычно проводить свои селекционные эксперименты с видами, которые воспроизводят гораздо больше быстро, так что количество времени и денег, требуемых значительно уменьшенный. Для этой цели обычно используются плодовые мушки и бактерии. сейчас. Плодовые мушки размножаются примерно через 2 недели после рождения, а бактерии, такие как кишечная палочка, обнаруженная в нашей пищеварительной системе, размножаются всего за 3-5 часов.
Вернуться в меню Практический тест Следующий Тема
 Сегодня мы знаем, что это связано с тем, что гены независимого
разные признаки расположены в разных хромосомах
.
Сегодня мы знаем, что это связано с тем, что гены независимого
разные признаки расположены в разных хромосомах
.
Copyright 1997-2013 Деннис
О’Нил. Все права защищены.
Все права защищены.
кредиты иллюстрации
Русский перевод
Введение в генетику – Основы биологии
«Генетика» — это наука о том, как наследуются признаки. Черта определяется как изменение внешнего вида наследуемой характеристики. Он стремится понять, как черты передаются из поколения в поколение. Прежде чем вы начнете изучать детали наследования, давайте рассмотрим некоторые темы, которые важны для понимания генетики.
Напомним, что гены представляют собой сегменты ДНК, длина которых обычно составляет несколько сотен или тысяч оснований. Каждый ген направляет производство белка в процессе синтеза белка: ДНК транскрибируется для получения мРНК; мРНК обеспечивает кодирование рибосомы для производства цепи аминокислот. Прочтите этот раздел книги, если вам нужно вернуться к этой теме: Как гены управляют производством белков?
Центральная догма – ДНК используется для производства РНК используется для производства белка. Фото кредит: ? Напомним, что эукариотические гены находятся в хромосомах и что каждая эукариотическая хромосома обычно содержит сотни или тысячи генов. У большинства эукариот, включая человека и других животных, каждая клетка содержит две копии каждой хромосомы. Причина, по которой у нас есть две копии каждого гена, заключается в том, что мы наследуем по одной копии от каждого родителя.
У большинства эукариот, включая человека и других животных, каждая клетка содержит две копии каждой хромосомы. Причина, по которой у нас есть две копии каждого гена, заключается в том, что мы наследуем по одной копии от каждого родителя.
В отличие от эукариот прокариоты имеют одну кольцевую хромосому. Это означает, что у них есть одна копия каждого гена.
Прочтите этот раздел книги, если вам нужно повторить эту тему: Как устроена ДНК в клетке
В женской клетке человеческого тела 23 пары хромосом. Эти хромосомы видны внутри ядра (вверху), удаляются из клетки во время клеточного деления (справа) и располагаются в соответствии с длиной (слева) в порядке, называемом кариотипом. На этом изображении хромосомы были подвергнуты флуоресцентному окрашиванию, чтобы их можно было различить. (кредит: «718 Bot»/Wikimedia Commons, Национальное исследование генома человека) Хромосомы наследуются потомством от родителей через яйцеклетку или сперматозоид. Внутри одной яйцеклетки или одного сперматозоида находится по одной копии каждого хронометра (всего 23 у человека). Когда яйцеклетка оплодотворяется спермой, полученная зигота (оплодотворенная яйцеклетка) будет содержать две копии каждой хромосомы, как и каждый из ее родителей.
Когда яйцеклетка оплодотворяется спермой, полученная зигота (оплодотворенная яйцеклетка) будет содержать две копии каждой хромосомы, как и каждый из ее родителей.
Мейоз — это процесс образования яйцеклеток и сперматозоидов. Яйцеклетки и сперматозоиды также известны как гамет . Во время мейоза одна копия каждой парной хромосомы перемещается в гамету. Клетки с одной копией каждой хромосомы известны как «9».0184 гаплоидный ». Это разделение, или сегрегация, 90 184 гомологичных 90 185 (спаренных) хромосом означает также, что только одна из копий гена перемещается в гамету.
Потомство образуется, когда эта гамета соединяется с гаметой от другого родителя и восстанавливаются две копии каждого гена (и хромосомы). Прочтите этот раздел книги, если вам нужна дополнительная информация по этой теме: Обзор мейоза
. Во время мейоза ДНК копируется один раз, затем клетка делится дважды. Это производит клетки с вдвое меньшим количеством генетической информации, чем исходная клетка (1 копия каждой хромосомы).