Рецептор — все статьи и новости
Рецептор (от лат. receptum — «принимающий») — чувствительное образование на окончаниях нервных клеток (дендритов) живого организма, способное воспринимать внешние и внутренние раздражения и преобразовывать их в нервные импульсы.
Образования, реагирующие на информацию извне, называются экстерорецепторами, а изнутри — интерорецепторами. К основным источникам раздражения относятся высокие звуковые частоты, изменение кровяного давления, химические вещества, перепады температур, изменения электрического и магнитного поля, свет, вкус, удары, смена положения тела в пространстве. Некоторые рецепторы способны воспринимать только один источник раздражения, другие — сразу несколько. Информация о любых раздражителях передается в центральную нервную систему.
Образования, реагирующие на разные раздражители, имеют неодинаковую форму. Своим строением рецепторы могут напоминать колбы, шайбы, шары, нити, спирали, палочки, волоски. Рецепторы, получающие информацию на относительно далеком расстоянии, называются дистантными, а те, которые реагируют на источник раздражения в непосредственной близости — контактными. При частых контактах рецепторов с раздражителями активность первых может снижаться. В таком случае говорят, что рецептор адаптировался к раздражителю. Быстрее всего привыкают тактильные рецепторы, медленнее — реагирующие на химические вещества.
При частых контактах рецепторов с раздражителями активность первых может снижаться. В таком случае говорят, что рецептор адаптировался к раздражителю. Быстрее всего привыкают тактильные рецепторы, медленнее — реагирующие на химические вещества.
В роли рецепторов могут выступать как свободные дендриты, так и части нервных окончаний в глубине тканей. Для таких «спрятанных» рецепторов существует специальный термин — «инкапсулированные». Большая часть этих образований — кожные. У многих инкапсулированных рецепторов есть своё особое название, которое они получили в честь открывших их ученых: тельца Мейснера, диски Меркеля (в исследование которых внёс свой вклад и отечественный ученый Александр Догель), тельца Пачини, тельца Руффини, колбы Краузе. Большинство из названных образований было исследовано еще в XIX веке.
Источник картинки: http://bit.ly/2dqWv1w
РЕЦЕПТОРЫ | Психологическая энциклопедия 1vc0
Оглавление раздела ощущения:
Рецептор — орган, специально приспособленный для рецепции раздражений, легче, чем прочие органы или нервные волокна, поддаётся раздражению; он отличается особенно низкими порогами раздражения, т. е. его чувствительность, обратно пропорциональная порогу, особенно высока. В этом первая особенность рецептора как специализированного аппарата: обладая особенно большой чувствительностью, он специально приспособлен для рецепции раздражений.
е. его чувствительность, обратно пропорциональная порогу, особенно высока. В этом первая особенность рецептора как специализированного аппарата: обладая особенно большой чувствительностью, он специально приспособлен для рецепции раздражений.
При этом рецепторы приспособлены для рецепции не любых раздражителей. Каждый рецептор специализируется применительно к определённому раздражителю. Формируясь в процессе развития под воздействием определённого вида раздражителей, каждый рецептор является поэтому приспособленным для рецепции именно этого вида раздражителей. Так, образуются тангорецепторы, приспособленные к рецепции прикосновения, густорецепторы — для рецепции вкусовых раздражений, стиборецепторы — для обонятельных, приспособленные для рецепции звука и света фоно- и фоторецепторы — слух, зрение. Каждый из этих рецепторов установлен на особый вид раздражителя, который является адекватным для него.
Таким образом, специальная приспособленность к рецепции раздражений, выражающаяся в особо высокой чувствительности, — во-первых, и приспособленность к рецепции специальных раздражителей, т. е. специализация рецепторов по виду раздражителей, — во-вторых, составляют основные черты, характеризующие рецепторный аппарат.
е. специализация рецепторов по виду раздражителей, — во-вторых, составляют основные черты, характеризующие рецепторный аппарат.
В парадоксальной форме специализация органов чувств, или рецепторов, выражается в том, что и неадекватный раздражитель, воздействуя на определённый рецептор, может вызвать специфические для него ощущения. Так, сетчатка даёт световые ощущения при воздействии на неё как светом, так и электрическим током или давлением («искры из глаз сыплются» при ударе). С другой стороны, механический раздражитель может дать ощущение давления, звука или света в зависимости от того, воздействует ли он на осязание, слух или зрение. Получающееся таким образом нарушение адекватного соотношения между ощущением и раздражителем представляет собой всё же лишь исключение. Тем не менее, основываясь на этих фактах и опираясь на специализацию «органов чувств», И. Мюллер выдвинул свой принцип специфической энергии органов чувств. Основу его составляет бесспорное положение, заключающееся в том, что все специфицированные ощущения находятся в определённом соотношении с гистологически специфицированными органами, их обусловливающими.
На этой основе И. Мюллер выдвигает другую идею, согласно которой ощущение зависит не от природы раздражителя, а от органа или нерва, в котором происходит процесс раздражения, и является выражением его специфической энергии. Посредством зрения, например, по И. Мюллеру, познаётся несуществующий во внешнем мире свет, поскольку глаз наш доставляет впечатление света и тогда, когда на него действует электрический или механический раздражитель, т. е. в отсутствие физического света. Ощущение света признаётся выражением специфической энергии сетчатки: оно — лишь субъективное состояние сознания. Включение физиологических процессов в соответствующем аппарате в число объективных, опосредующих условий ощущения превращается, таким образом, в средство отрыва ощущения от его внешней причины и признания субъективности ощущения. Из связи субъекта с объектом ощущение превращается во включённую между субъектом и объектом завесу.
Из связи субъекта с объектом ощущение превращается во включённую между субъектом и объектом завесу.
На этой основе и сложилась та концепция, которую сформулировал Гельмгольц. Плеханов, опираясь на неё, выдвинул свою теорию иероглифов, которую Ленин подверг острой критике: «Бесспорно, что изображение никогда не может всецело сравниться с моделью, но одно дело изображение, другое дело символ, условный знак. Изображение необходимо и неизбежно предполагает объективную реальность того, что «отображается». «Условный знак», символ, иероглиф суть понятия, вносящие совершенно ненужный элемент агностицизма». [В. И. Ленин, Соч., т. XIII, 1931. стр. 193.]
Стоит подойти к интерпретации того позитивного фактического положения, которое лежит в основе субъективно-идеалистической надстройки, возведённой над нею И. Мюллером, чтобы те же факты предстали в совсем ином освещении. В процессе биологической эволюции сами органы чувств формировались в реальных взаимоотношениях организма со средой, под воздействием внешнего мира.
 Критерием для различения одних от других служит действие, практика, контролирующая объективность наших ощущений как субъективного образа объективного мира.
Критерием для различения одних от других служит действие, практика, контролирующая объективность наших ощущений как субъективного образа объективного мира.Элементы психофизики
Наличие зависимости ощущений от внешних раздражений заставляет поставить вопрос о характере этой зависимости, т. е. об основных закономерностях, которым она подчиняется. Это центральный вопрос так называемой психофизики. Её основы заложены исследованиями Э. Г. Вебера и Г. Т. Фехнера. Оформление она получила в «Элементах психофизики» (1859) Фехнера, оказавших значительное влияние на дальнейшие исследования. Основной вопрос психофизики — это вопрос о порогах. Различают
Исследования по психофизике установили прежде всего, что не всякий раздражитель вызывает ощущение. Он может быть так слаб, что не вызовет никакого ощущения. Мы не слышим множества вибраций окружающих нас тел, не видим невооружённым глазом множества постоянно вокруг нас происходящих микроскопических изменений. Нужна известная минимальная интенсивность раздражителя для того, чтобы вызвать ощущение. Эта минимальная интенсивность раздражения называется нижним абсолютным порогом. Нижний порог даёт количественное выражение для чувствительности: чувствительность рецептора выражается величиной, обратно пропорциональной порогу: , где E — чувствительность и J — пороговая величина раздражителя.
Нужна известная минимальная интенсивность раздражителя для того, чтобы вызвать ощущение. Эта минимальная интенсивность раздражения называется нижним абсолютным порогом. Нижний порог даёт количественное выражение для чувствительности: чувствительность рецептора выражается величиной, обратно пропорциональной порогу: , где E — чувствительность и J — пороговая величина раздражителя.
За вопросом о том, имеет ли вообще место ощущение определённого вида (зрительное, слуховое и т. д.), неизбежно следует вопрос об условиях различения различных раздражителей. Оказалось, что наряду с абсолютными существуют разностные пороги различения. Э. Вебер установил, что требуется определённое соотношение между интенсивностями двух раздражителей для того, чтобы они дали различные ощущения. Это соотношение выражено в законе, установленном Э. Вебером:
Оказалось, что наряду с абсолютными существуют разностные пороги различения. Э. Вебер установил, что требуется определённое соотношение между интенсивностями двух раздражителей для того, чтобы они дали различные ощущения. Это соотношение выражено в законе, установленном Э. Вебером:
ΔJ÷J=K,
где J обозначает раздражение, ΔJ — его прирост, K — постоянная величина, зависящая от рецептора.
Так, в ощущении давления величина прибавки, необходимой для получения едва заметной разницы, должна всегда равняться приблизительно 1/30 исходного веса, т. е. для получения едва заметной разницы в ощущении давления к 100 г нужно добавить 3,4 г, к 200 — 6,8 г, к 300 — 10,2 г и т. д. Для силы звука эта константа равна 1/10, для силы света — 1/100 и т. д.
Дальнейшие исследования показали, что закон Э. Вебера действителен лишь для раздражителей средней величины: при приближении к абсолютным порогам величина прибавки перестаёт быть постоянной величиной. Наряду с этим ограничением закон Э. Вебера допускает, как оказалось; и расширение. Он применим не только к едва заметным, но и ко всяким различиям ощущений. Различия между парами ощущений кажутся нам равными, если равны геометрические соотношения соответствующих раздражителей. Так, увеличение силы освещения от 25 до 50 свечей даёт субъективно такой же эффект, как увеличение от 50 до 100.
Наряду с этим ограничением закон Э. Вебера допускает, как оказалось; и расширение. Он применим не только к едва заметным, но и ко всяким различиям ощущений. Различия между парами ощущений кажутся нам равными, если равны геометрические соотношения соответствующих раздражителей. Так, увеличение силы освещения от 25 до 50 свечей даёт субъективно такой же эффект, как увеличение от 50 до 100.
Исходя из закона Э. Вебера, Г. Фехнер сделал допущение, что едва заметные разницы в ощущениях можно рассматривать как равные, поскольку все они — величины бесконечно малые, и принять их как единицу меры, при помощи которой можно численно выразить интенсивность ощущений как сумму (или интеграл) едва заметных (бесконечно малых) увеличений, считая от порога абсолютной чувствительности. В результате он получил два ряда переменных величин — величины раздражителей и соответствующие им величины ощущений. Ощущения растут в арифметической прогрессии, когда раздражители растут в геометрической прогрессии.
E = K · log J + C,
где K и C суть некоторые константы. Эта формула, определяющая зависимость интенсивности ощущений (в единицах едва заметных перемен) от интенсивности соответствующих раздражителей, и представляет собой так называемый психофизический закон Вебера-Фехнера.
Допущенная при этом Фехнером возможность суммирования бесконечных, а не только конечных разностей ощущений, большинством исследований считается произвольной. Помимо того нужно отметить, что ряд явлений, вскрытых новейшими исследованиями чувствительности, не укладывается в рамки закона Вебера-Фехнера. Особенно значительное противоречие с законом Вебера-Фехнера обнаруживают явления протопатической чувствительности, поскольку ощущения в области протопатической чувствительности не обнаруживают постепенного нарастания по мере усиления раздражения, а по достижении известного порога сразу же появляются в максимальной степени.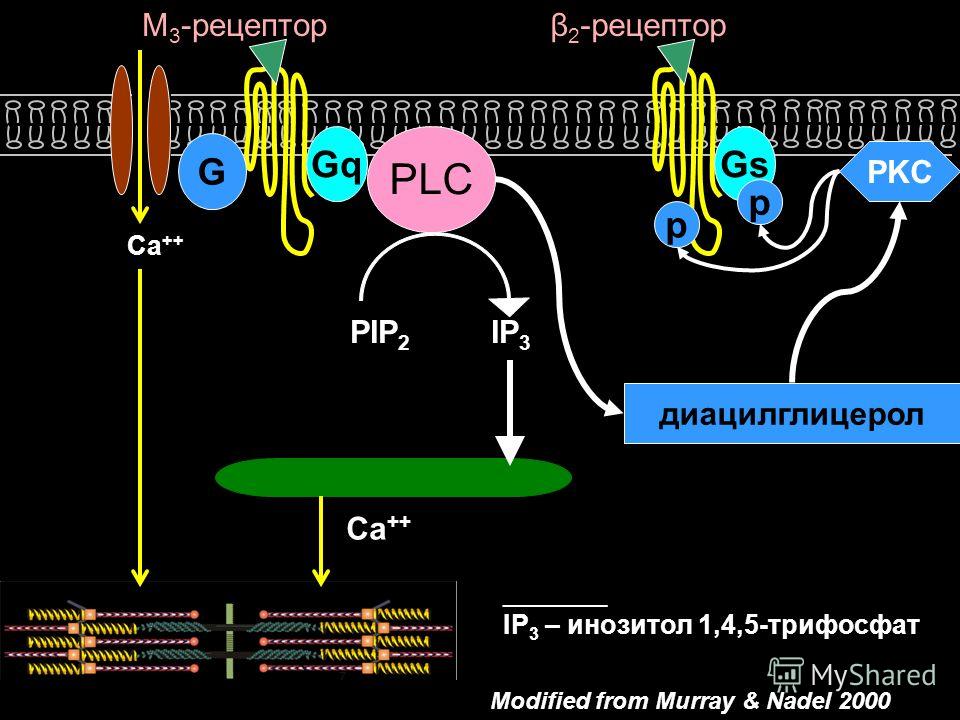 Они приближаются по своему характеру к типу реакций по принципу «всё или ничего». Не согласуются, по-видимому, с законом Вебера-Фехнера и некоторые данные современной электрофизиологии органов чувств.
Они приближаются по своему характеру к типу реакций по принципу «всё или ничего». Не согласуются, по-видимому, с законом Вебера-Фехнера и некоторые данные современной электрофизиологии органов чувств.
Дальнейшие исследования Г. Гельмгольца, подтверждённые П. П. Лазаревым, заменили первоначальную формулировку закона Вебера-Фехнера более сложной формулой, выражающей очень общий принцип, управляющий всеми явлениями раздражения. Однако и попытка Лазарева выразить переход раздражения в ощущение в математических уравнениях не охватывает всего многообразия процессов чувствительности и перехода раздражения в ощущение.
Для определения порогов была разработана целая система методов психофизического исследования. Из них основные: 1) метод едва заметных различий: прогрессивно изменяют — увеличивают или уменьшают — раздражитель, пока испытуемый не начнёт или не перестанет замечать разницу; 2) метод истинных и ложных случаев: испытуемому предъявляют для сравнения два любых раздражителя и предлагают определить, какой из них больше; 3) метод средних ошибок, или метод константности: испытуемый должен подобрать к данному ему раздражителю равные. При всех этих методах пороги определяются как статистические средние.
При всех этих методах пороги определяются как статистические средние.
Значение измерения порогов заключается в том, что они являются главной основой для точного, количественно выраженного определения дифференциальных различий в сенсорной области — от вида к виду, от индивида к индивиду и у одного и того же индивида в различных условиях — в зависимости от утомления, упражнения, образования и т. д. Тем самым они дают возможность исследовать и значение всех влияющих на сенсорную область высших факторов и косвенно устанавливают их уровень. Они поэтому доставляют данные для ряда выводов, имеющих и практическую значимость.
Пороги и, значит, чувствительность органов никак не приходится представлять как некие раз и навсегда фиксированные неизменные лимиты. Целый ряд исследований советских авторов показал их чрезвычайную изменчивость. Так, А. И. Богословский [А. И. Богословский, Опыт выработки сенсорных условных рефлексов у человека, «Физиологический журнал СССР», 1938, стр. 1017. ], К. X. Кекчеев [К. X. Кекчеев, «Бюллетень эксперим. биологии и медицины», 1935, вып. 5—6. стр. 358.] и А. О. Долин [А. О. Долин, «Архив биологических наук», 1936, вып. 1—2.] показали, что чувствительность органов чувств может изменяться посредством образования интерсенсорных условных рефлексов (которые подчиняются вообще тем же законам, что и обычные двигательные и секреторные условные рефлексы). Очень отчётливо явление сенсибилизации было в последнее время выявлено рядом исследовании в отношении слуховой чувствительности. Так, А. И. Бронштейн [А. И. Бронштейн, О синтезирующем влиянии звукового раздражения на орган слуха // Бюллетень экспериментальной биологии и медицины. 1936. Т. I. Вып. 4: Сообщения 1 и 2; Т. II. Вып. 5: Сообщение 3.] констатировал понижение порогов слышимости под влиянием повторяющихся звуковых раздражений. Б. М. Теплов обнаружил резкое понижение порогов различения высоты в результате очень непродолжительных упражнений (см. раздел «Слуховые ощущения» этой главы). В.
], К. X. Кекчеев [К. X. Кекчеев, «Бюллетень эксперим. биологии и медицины», 1935, вып. 5—6. стр. 358.] и А. О. Долин [А. О. Долин, «Архив биологических наук», 1936, вып. 1—2.] показали, что чувствительность органов чувств может изменяться посредством образования интерсенсорных условных рефлексов (которые подчиняются вообще тем же законам, что и обычные двигательные и секреторные условные рефлексы). Очень отчётливо явление сенсибилизации было в последнее время выявлено рядом исследовании в отношении слуховой чувствительности. Так, А. И. Бронштейн [А. И. Бронштейн, О синтезирующем влиянии звукового раздражения на орган слуха // Бюллетень экспериментальной биологии и медицины. 1936. Т. I. Вып. 4: Сообщения 1 и 2; Т. II. Вып. 5: Сообщение 3.] констатировал понижение порогов слышимости под влиянием повторяющихся звуковых раздражений. Б. М. Теплов обнаружил резкое понижение порогов различения высоты в результате очень непродолжительных упражнений (см. раздел «Слуховые ощущения» этой главы). В. И. Кауфман — в противовес тенденции К. Сишора (Seashore), Г. М. Уиппла и др. рассматривать индивидуальные различия порогов звуковысотной чувствительности исключительно как неизменяющиеся природные особенности организма — экспериментально показал, во-первых, зависимость порогов (так же как самого типа) восприятия высотных разностей от характера музыкальной деятельности испытуемых (инструменталисты, пианисты и т. д.) и, во-вторых, изменяемость этих порогов (и самого типа) восприятия высотных разностей. Кауфман поэтому приходит к тому выводу, что способность различения высоты звука зависит от конкретных особенностей деятельности данной личности и может в известной мере изменяться в зависимости от неё. [В. И. Кауфман, Восприятие малых высотных разностей. Сб. «Исследования по проблеме чувствительности» под ред. В. П. Осипова и Б. Г. Ананьева, т. XIII, 1940.] Н. К. Гусев пришёл к аналогичным результатам о роли практики дегустации в развитии вкусовой чувствительности. [Там же.]
И. Кауфман — в противовес тенденции К. Сишора (Seashore), Г. М. Уиппла и др. рассматривать индивидуальные различия порогов звуковысотной чувствительности исключительно как неизменяющиеся природные особенности организма — экспериментально показал, во-первых, зависимость порогов (так же как самого типа) восприятия высотных разностей от характера музыкальной деятельности испытуемых (инструменталисты, пианисты и т. д.) и, во-вторых, изменяемость этих порогов (и самого типа) восприятия высотных разностей. Кауфман поэтому приходит к тому выводу, что способность различения высоты звука зависит от конкретных особенностей деятельности данной личности и может в известной мере изменяться в зависимости от неё. [В. И. Кауфман, Восприятие малых высотных разностей. Сб. «Исследования по проблеме чувствительности» под ред. В. П. Осипова и Б. Г. Ананьева, т. XIII, 1940.] Н. К. Гусев пришёл к аналогичным результатам о роли практики дегустации в развитии вкусовой чувствительности. [Там же.]
Вообще пороги чувствительности не представляют собой абсолютной, неизменной величины. Экспериментальное исследование показывает, что даже такие явления, которые обусловлены в основном, казалось бы, периферическими факторами, не предопределяются органом самим по себе. Так, например, снижение световой чувствительности периферического зрения в ходе темновой адаптации, вызванное предварительным «засветом» периферии сетчатки, существенно зависит от центральных психологических факторов и может быть (как показывает проводимое в Институте психологии исследование Семёновской) снято при внимании.
Экспериментальное исследование показывает, что даже такие явления, которые обусловлены в основном, казалось бы, периферическими факторами, не предопределяются органом самим по себе. Так, например, снижение световой чувствительности периферического зрения в ходе темновой адаптации, вызванное предварительным «засветом» периферии сетчатки, существенно зависит от центральных психологических факторов и может быть (как показывает проводимое в Институте психологии исследование Семёновской) снято при внимании.
Пороги чувствительности существенно сдвигаются в зависимости от отношения человека к той задаче, которую он разрешает, дифференцируя те или иные чувственные данные. Один и тот же физический раздражитель одной и той же интенсивности может оказаться и ниже и выше порога чувствительности и, таким образом, быть или не быть замеченным в зависимости от того, какое значение он приобретает для человека: появляется ли он как безразличный момент окружения для данного индивида или становится имеющим определённое значение показателем существенных условий его деятельности. Поэтому, для того чтобы исследование чувствительности дало сколько-нибудь законченные результаты и привело к практически значимым выводам, оно должно, не замыкаясь в рамках одной лишь физиологии, перейти и в план психологический. Психологическое исследование имеет, таким образом, дело не только с «раздражителем», но и с предметом, и не только с органом, но и с человеком. Этой более конкретной трактовкой ощущения в психологии, связывающей его со всей сложной жизнью личности в её реальных взаимоотношениях с окружающим миром, обусловлено особое значение психологического и психофизиологического, а не только физиологического, исследования для разрешения вопросов, связанных с нуждами практики.
Поэтому, для того чтобы исследование чувствительности дало сколько-нибудь законченные результаты и привело к практически значимым выводам, оно должно, не замыкаясь в рамках одной лишь физиологии, перейти и в план психологический. Психологическое исследование имеет, таким образом, дело не только с «раздражителем», но и с предметом, и не только с органом, но и с человеком. Этой более конкретной трактовкой ощущения в психологии, связывающей его со всей сложной жизнью личности в её реальных взаимоотношениях с окружающим миром, обусловлено особое значение психологического и психофизиологического, а не только физиологического, исследования для разрешения вопросов, связанных с нуждами практики.
Психофизиологические закономерности
Характеристика ощущений не исчерпывается психофизическими закономерностями. Для чувствительности органа имеет значение и физиологическое его состояние (или происходящие в нём физиологические процессы). Значение физиологических моментов сказывается прежде всего в явлениях адаптации, в приспособлении органа к длительно воздействующему раздражителю; приспособление это выражается в изменении чувствительности — понижении или повышении её. Примером может служить факт быстрой адаптации к одному какому-либо длительно действующему запаху, в то время как другие запахи продолжают чувствоваться так же остро, как и раньше.
Примером может служить факт быстрой адаптации к одному какому-либо длительно действующему запаху, в то время как другие запахи продолжают чувствоваться так же остро, как и раньше.
Уменьшение интенсивности ощущения при длительном воздействии раздражителя находит себе объяснение в уменьшении частоты нервных импульсов, пробегающих по афферентному нерву, идущему от рецептора в центральную нервную систему. Эдриан (Adrian) установил, что все рецепторы могут быть разбиты на две группы: 1) рецепторы с быстрой адаптацией к раздражителю и быстрым привыканием к нему, что выражается ослаблением ощущения при продолжающемся раздражении органа чувств (тактильные рецепторы), и 2) рецепторы с медленной адаптацией, при деятельности которых ощущение почти не ослабевает (проприорецепторы).
С адаптацией тесно связано и явление контраста, которое сказывается в изменении чувствительности под влиянием предшествующего (или сопутствующего) раздражения. Так, в силу контраста обостряется ощущение кислого после ощущения сладкого, ощущение холодного после горячего и т. д. Следует отметить также свойство рецепторов задерживать ощущения, выражающееся в более или менее длительном последействии раздражений. Так же как ощущение не сразу достигает своего окончательного значения, оно не сразу исчезает после прекращения раздражения, а держится некоторое время и лишь затем постепенно исчезает. Благодаря задержке при быстром следовании раздражений одного за другим происходит слияние отдельных ощущений в единое, слитное целое, как, например, при восприятии мелодий, кинокартины и пр.
Дифференциация и специализация рецепторов не исключает их единства, выражающегося в их взаимодействии. Это взаимодействие рецепторов выражается, во-первых, во влиянии, которое раздражение одного рецептора оказывает на пороги другого. Так, зрительные раздражения влияют на пороги слуховых, а слуховые раздражения — на пороги зрительных (Лазарев, Кравков), точно так же на пороги зрительных ощущений оказывают влияние и обонятельные ощущения (см. дальше).
На взаимодействии рецепторов основан метод сенсибилизации одних органов чувств и, в первую очередь, глаза и уха путём действия на другие органы чувств слабыми или кратковременными, адекватными для них, раздражениями.
Взаимосвязь ощущений проявляется, во-вторых, в так называемой синестезии. Под синестезией разумеют такое слияние качеств различных сфер чувствительности, при котором качества одной модальности перекосятся на другую, разнородную, — например, при цветном слухе качества зрительной сферы — на слуховую. Формой синестезии, относительно часто наблюдающейся, является так называемый цветной слух (audition colorée). У некоторых людей (например у А. Н. Скрябина; в ряде случаев, которые наблюдал А. Бине; у мальчика, которого исследовал А. Ф. Лазурский; у очень музыкального подростка, которого имеет возможность наблюдать автор) явление цветного слуха выражено очень ярко. Отдельные выражения, отражающие синестезии различных видов ощущений, получили права гражданства в литературном языке; так, например, говорят о кричащем цвете, а также о тёплом или холодном колорите и о тёплом звуке (тембре голоса), о бархатистом голосе.
Теоретически природа этого явления не вполне выяснена. Иные авторы склонны объяснять его общностью аффективных моментов, придающих ощущениям различных видов один и тот же общий им эмоционально-выразительный характер.
Взаимодействие рецепторов выражается, наконец, в той взаимосвязи ощущений различных рецепторов, которое постоянно происходит в каждом процессе восприятия любого предмета или явления. Такое взаимодействие осуществляется в совместном участии различных ощущений, например зрительных и осязательных, в познании какого-нибудь предмета или его свойства, как-то — форма, фактура и т. п. (Даже тогда, когда непосредственно в восприятии участвует лишь один рецептор, ощущения, которые он нам доставляет, бывают опосредованы данными другого. Так, при осязательном распознавании формы предмета, когда зрение почему-либо выключено, осязательные ощущения опосредуются зрительными представлениями.) В самом осязании имеет место взаимодействие собственно кожных ощущений прикосновения с мышечными, кинестетическими ощущениями, к которым при ощущении поверхности предмета примешиваются ещё и температурные ощущения. При ощущении терпкого, едкого и т. п. вкуса какой-нибудь пищи к собственно вкусовым ощущениям присоединяются, взаимодействуя с ними, ощущения осязательные и лёгкие болевые. Это взаимодействие осуществляется и в пределах одного вида ощущений. В области зрения, например, расстояние влияет на цвет, ощущения глубины — на форму и т. д. Из всех форм взаимодействия эта последняя, конечно, важнейшая, потому что без неё вообще не существует восприятия действительности.
Receptor Autoradiography Protocol for the Localized Visualization of Angiotensin II Receptors
Обзор метаболического пути ренин-ангиотензин системы показана на рисунке 1 и прямой акцент на подтипы рецепторов ангиотензина II (AT 1 R и AT 2 R) описывается на рисунке 2. Рисунок 3 показывает передачу корональной мозга разделы на предметные стекла микроскопа, которые затем проходят через процедуру рецептора авторадиографии с использованием заданной концентрации 125 I-Siang II , как показано на рисунке 4. Рисунок 5 иллюстрирует стадию сушки для слайдов в авторадиографии рецепторов , которые затем наносимой на картоне как показано на рисунке 6, а впоследствии развиты. в таблице 2 перечислены реагенты , используемые для тионин процедуры окрашивания для срезов тканей слайд — монтажа , описанных на рисунке 7. Калибровка стандартные единицы измерения для количественной оценки определяются баСЭД на дату анализа , как показано в таблице 3. На рисунке 8 показано создание стандартной кривой , относящейся 125 I концентрации (фмоль / г ткани) заснять экспозиции (относительная оптометрическое плотности). Рисунок 11 иллюстрирует различие между «NSP» и «всего» группы, а также этикетки ткани, а на рисунке 10 описывает установку порога до почти нулевого значения , чтобы получить точное среднее значение для всей области сканирования (Scan Area пикселей). Таблица 4 показывает количественное процесс получения специфически связывающийся на основе вычитания ‘неспецифический’ из » общего значения. «Неспецифический» связывания , который содержит лозартан и / или PD123319 создается количеством радиолиганда , связанного с не-AT 1, или AT 2 рецепторов. Все радиолиганда , связанного с АТ 1 рецепторов в присутствии PD123319 дает общее » связывания. F igure 11 отображает конечные измеренных площадей ПВЯ в общей сложности против неспецифического связывания , смежному с тионин окрашенном секции для анатомического подтверждения ПВН.
Рисунок 1:. Метаболический путь системы ренин — ангиотензин (RAS) в организме Печень выпускает Зимоген ангиотензиноген, который расщепляется овально-секретируется ренина, образуя ангиотензин I. ангиотензин — превращающего фермента (АПФ) преобразует Ang I в ангиотензин II ( анг II), главным предшественником сердечно-сосудистых заболеваний. Анг II вызывает вазоконстрикцию и повышает кровяное давление, сердечный выброс, солевой аппетит, жажда, и задержку воды. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рис . 2: Функции типа ангиотензина 1 и 2 рецепторов (AT 1 R, AT 2 R) AT 1 R патологически влияет на сердечно — сосудистые заболевания, напрямую способствует увеличению жажды и соли аппетит и оказывает многие другие периферийные клеточные активности, а также центральный психологические эффекты. AT 2 R является физиологическим антагонистом рецептора AT 1, помогая в эффективности AT 1 R антагонистов 36 , таким образом , имеющие благотворное воздействие на сосудистую структуру. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рисунок 3: Cryosectioning головного мозга на предметные стекла микроскопа при толщине 20 мкм. В зависимости от температуры криостата, секции будут иногда показывать сгибы, трещины, скручивание, или становятся более плотными на микротома лезвия или держателя ножа. В таких случаях секция отбрасывается и делается отметка, чтобы указать, что дополнительные 20 мкм отделит этот раздел из предыдущего раздела. Секции должны быть собраны на слайдах в вертикальном направлении, чтобы максимизировать общую площадь , необходимую для заполнения слайда и , следовательно , собрать наибольшее количество разделов на каждом слайде. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рис . 4: Пример экспериментального проектирования и протокола для рецепторного Авторадиография Differentiданию ‘Всего’ (-2) и «неспецифический» (-1) непосредственно связано с наличием и отсутствием антагонистов рецепторов подтипа. Протокол может меняться в зависимости от количества слайдов, и будет увеличиваться с шагом 2 для каждой пары полных и неспецифических слайдов. концентрация радиолиганда, предварительно определяется получают путем разбавления запаса в AM5. Затем этот раствор равномерно распределяется по инкубационных контейнеров. Прямые отсчеты до и после инкубации , чтобы установить точную концентрацию радиолиганда. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рис . 5: Конфигурация для сушки станции в авторадиографии рецептора Время высыхания будет варьироваться в зависимости от эффективности ударасушилки, а также возможность промокните лишнюю воду после последнего полоскания. Слайды будут сушиться после того, как раздел получается от прозрачного до белого. Если слайды не полностью высушенный то радиолиганда может диффундировать от рецептора , производящей нечеткое изображение, которое не является специфическим для рецептора сайтов связывания. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рисунок 6:.. Разоблачение обработанные слайды рентгеновской пленки с использованием специализированной кассеты слайдов настройки рекомендуется следовать шаблону , где облегчила сравнение обеих групп Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рисунок 7: Настройка для тионин окрашиванию. Слайды загружаются в держатель слайдов и погружена в последовательные решения. Выбор времени позволяет эффективно окрашивания вещества Нисслю (шероховатой эндоплазматической сети) нейронов. Установочный среда приложения к слайдам следующих обезвоживания позволяет для крепления покровного к слайду, сохраняя при этом секции для сравнительного анализа. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рис . 8: Установка стандарта калибровки Каждый фильм содержит свои индивидуальные калибровочные стандарты, основанные на пасте мозга и время , в которое было сделано эксперимент , Линии регрессии, описывающая лучшие значения подходят порождается собственной программой системы визуализации для измеренных стандартов мозга пасты. Это преобразует относительную оптометрическое плотность (пруток) значение из калибровочных стандартов (кал БППП, в красном овале) в единицах связывания радиолиганда, в этом примере единицы (фмоль / г, обведены красным) являются fmole Радиолигандом связаны на грамм влажного вес ткани (фмоль / г). Измерение стандартов пасты мозга осуществляется с помощью кругового инструмента и помещен в середине образца, показанный справа в псевдоцвете с помощью значка просмотра изображения (обведено красным). Круг для измерения стандартной калибровки не должны покрывать всю выборку, а измерить даже площадь стандарта. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
/53866fig9.jpg «/>
Рис . 9: Определение групп по количеству проб, неспецифическое / общее, так и в регионах для чтения образцов Поскольку несколько мозгов, области мозга, а также несколько экспериментальных групп количественно вместе, крайне важно проводить различие между всеми факторами. Субъект указывает, что образец мозга, а также может указывать на группу; раздел указывает на то, является ли это неспецифическая (NSP) или полное, в то время как область позволяет идентифицировать несколько областей мозга, которые в настоящее время выборки. В баре образец инструмента (красный прямоугольник) описывает различные инструменты выборки, например, круг, квадрат, шаблон, Freehand, ластика; который может быть использован, чтобы очертить область, которую необходимо измерить. Значения ROD, полученные в области инструмента выборки записываются на электронную таблицу, как преобразованные единиц связывания, фмоль / г (обведено красным). Кроме того, общая площадь сканирования в пробоотборник записывается в колонке «Сканировать пикселей» в то время как общая тArget площадь (количество пикселей , которые превышают пороговое значение ( как описано на рисунке 10 легенде) записывается в колонке «Общая площадь Tgt пикселей». Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рисунок 10: Определение порога для образцов для измерения на основе стандартов Область сканирования является уникальным для каждого фильма, и зависит от стандартов , установленных.. С помощью инструмента порога (обведено красным), чтобы установить пороговые значения для диапазона плотности для области сканирования в фмоль / г позволит для областей измеренных (очерчены диагональными стрелками) падать между диапазоном от 0, до точки, где кривая калибровки начинает асимптоты, указывающего, что пленка приближается к максимальной экспозиции, за которой дополнительный Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Рисунок 11: Типичные изображения авторадиографии связывания с рецептором к корональных участков женского спонтанно гипертензивных крыс (SHR) при ~ 1,8 мм каудально брегмы, отображающие паравентрикулярной ядро гипоталамуса (PVN) и ядра слоем вспомогательного обонятельного тракта (BAOT ). (A </ сильный>) Общее связывание AT 1 R связывания (красно- PVN, желто- BAOT). (B) неспецифическое связывание с лозартаном (антагонист AT1R) и PD123319 (антагонист AT2R). (С) Общее связывание AT 1 R связывание с установкой такой произвольный порог, чтобы очертить форму PVN (черной заливкой) , превышающей пороговое значение в пределах зоны сканирования , которая записана в Total TGT Area). (D) тионин витражное секция для анатомического подтверждения структур , показывающих высокий общий уровень связывания, а также для сравнения с другими областями мозга или других экспериментальных групп. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
| Реагенты | Количество |
| NaCl | 8,76г |
| Na 2 EDTA | 1,86 г |
| Bacitracin | 141 мг |
| 500 мМ двухосновного Na 2 PO 4 | 100 мл |
| Дистиллированная Деионизированная вода | 900 мл |
| Отрегулируйте рН до 7,1 до 7,2 при помощи NaOH или HCl | |
Таблица 1: среды для анализа (AM5) Химические компоненты буфера , используемого в процессе подготовки ткани..
Таблица 2:. Тионин Stain РЕАГЕНТЫ тионин пятно состоит из дистиллированной воды, 1,0 М ацетата натрия и 1,0 М раствора ледяная уксусная кислота, титровали до рН 4,3 , перед добавлением 0,5% -ного раствора тионин.
| Дни прошлое ссылки Дата | |||||||||
| партия | фмоль / г сырого веса | Ссылка Дата | 1 | 2 | 3 | 4 | 5 | ||
| стандарт | нКи / мг | фмоль / г | ДПМ / г | фмоль / г | FOM / г | фмоль / г | фмоль / г | фмоль / г | фмоль / г |
| номер | 30 мая | 30 мая | 30 мая | 30 мая | 31 мая | 1-Jun | 2 июня | 3 июня | 4 июня |
| 1 Чонг> | 9706 | 4463 | 21547320 | 4463 | 4411 | 4361 | 4311 | 4261 | 4212 |
| 2 | 5838 | 2684 | 12960360 | 2684 | 2653 | 2623 | 2593 | 2563 | 2533 |
| 3 | 2798 | 1286 | 6211560 | 1286 | 1272 | 1257 | 1243 | 1228 | 1214 |
| 4 | 1451 | 667 | 3221220 | 667 | 659 | 652 | 644 | 637 | 630 |
| 5 | 787 | 362 | 1747140 | 362 | 358 | 354 | 350 | 345 | 342 |
| 6 | 385 | 177 | 854700 | 177 | 174 | 173 | 171 | 169 | 167 |
Таблица 3:. Ежедневная Decay Rate Расчеты для калибровочных стандартов Ежедневная электронная таблица скорость затухания для калибровочных стандартов необходимо будет создан на основе даты , на котором был выполнен эксперимент. Эта таблица основана на скорости первого порядка с постоянным периодом полураспада (T?) На 60 дней. N = N 0 * е кт, где N = концентрация 125 I, в момент времени Т относительно концентрации в момент времени 0 (N 0), а к = ln2 / т 1/2.
| Паравентрикулярной Nucleous гипоталамуса | цель | Плотность-фмоль / г | Область сканирования | Общая Целевая зона | Всего* | |
| Всего | 1 | 996 | 4383 | 4383 | 996 | |
| Всего | 2 | 921 | 3362 | 3362 | 921 | |
| Всего | 3 | 818 | 3445 | 3445 | 818 | специфически связывающийся |
| Всего | 4 | 710 | 2906 | 2906 | 710 | 967 |
| В СРЕДНЕМ | 861 | 3524 | 3524 | 861 | 895 | |
| 763 | </ TR>||||||
| Раздел | цель | Плотность-фмоль / г | Область сканирования | Общая Целевая зона | Неспецифический * | 678 |
| NSP | 1 | 53 | 3072 | 1737 | 30 | 826 |
| NSP | 2 | 52 | 2959 | 1455 | 26 | |
| NSP | 3 | 86 | 3180 | 2035 | 55 | |
| NSP | 4 | 53 | 3293 | +1995 | 32 | |
| В СРЕДНЕМ | 61 | 3126 | 1,805.5 | 36 | ||
| Кровать Nucleus аксессуара Olfacтори Тракт | ||||||
| Раздел | цель | Дэнс-фмоль / г | Область сканирования | Общая Целевая зона | Всего* | |
| Всего | 1 | 439 | 3632 | 3632 | 439 | |
| Всего | 2 | 435 | 3632 | 3632 | 435 | |
| Всего | 3 | 355 | 3632 | 3620 | 354 | |
| Всего | 4 | 435 | 3632 | 3631 | 435 | специфически связывающийся |
| Всего | 5 | 342 | 3632 | 3630 | 342 | 414 |
| Всего | 6 | 334 | 3632 | 3606 | 331 | 393 |
| В СРЕДНЕМ | 390 | 3632 | 3625 | 389 | 321 | |
| 394 | ||||||
| 315 | ||||||
| Раздел | цель | Дэнс-фмоль / г | Область сканирования | Общая Целевая зона | Неспецифический * | 320 |
| NSP | 1 | 49 | 3632 | +1846 | 25 | 360 |
| NSP | 2 | 64 | 3632 | 2362 | 42 | |
| NSP | 3 | 53 | 3632 | 2275 | 33 | > |
| NSP | 4 | 64 | 3632 | 2339 | 41 | |
| NSP | 5 | 51 | 3632 | +1879 | 26 | |
| NSP | 6 | 41 | 3632 | 966 | 11 | |
| В СРЕДНЕМ | 54 | 3632 | +1945 | 30 | ||
| * Общее количество и Non-speciic представляют собой единицы плотности фмоль / г. | ||||||
| Итого = плотность * общая целевая область / область сканирования | ||||||
| Неспецифический = плотность * общая целевая область / область сканирования | ||||||
| Общая целевая область пикселей, что превысило пороговое значение, устанавливают как можно ближе к 0,00 фмоль / г, как это возможно. | ||||||
| Пиксели, не превышающей порогзначение получают значение 0 для оценки общей плотности. | ||||||
| Пиксели, не превышающие пороговое значение, получают значение 0 для оценки неспецифического плотности. | ||||||
| Специфическое связывание Общий минус неспецифическое связывание, выраженной в единицах плотности фмоль / г. | ||||||
Таблица 4: Количественный анализ паравентрикулярной ядра гипоталамуса и слоем ядра аксессуар обонятельный тракт данных экспорта программного обеспечения обработки изображений в виде таблицы в фмоль / г, область сканирования и общей целевой области в пикселях.. «Неспецифический» связывание вычитается из » общего связывания с целью получения конкретного AT 1 количественного связывания.
Subscription Required. Please recommend JoVE to your librarian.
Ожирение у детей и подростков (недостаточность рецептора меланокортина)
Генетическое исследование, предназначенное для диагностики ожирения, ассоциированного с рецептором меланокортина 4-го типа. Мутации в гене MC4R являются наиболее распространенной генетической причиной тяжелого ожирения, проявляющегося в раннем возрасте.
Состав исследования
Синонимы английские
MC4R.
Метод исследования
Автоматическое секвенирование ДНК.
Генетические маркеры
Рецептор меланокортина 4 (MC4R) (все мутации).
Какой биоматериал можно использовать для исследования?
Буккальный (щечный) эпителий, венозную кровь.
Как правильно подготовиться к исследованию?
- Подготовки не требуется.
Общая информация об исследовании
До 6 % людей с тяжелым ожирением, развивающимся с детства, несут мутации в гене рецептора меланокортина 4-го типа (MC4R). Поэтому недостаточность MC4R – самая распространенная причина моногенного ожирения у человека.
Клиническая картина ожирения, связанного с недостаточностью рецептора меланокортина, может проявляться уже на первом году жизни:
- повышенный аппетит, переедание, отсутствие чувства сытости,
- быстрый набор веса (за счёт регулярного переедания),
- увеличение жировой, мышечной ткани сочетается с высоким ростом,
- как правило, не наблюдается нарушений функций щитовидной железы,
- половое развитие соответствует возрасту,
- задержек психомоторного развития, нарушения интеллекта нет,
- раннее развитие гиперинсулинемии, которая в 30 % случаев впоследствии приводит к сахарному диабету 2-го типа.
В зрелом возрасте это грозит метаболическим синдромом, сопутствующими сердечно-сосудистыми заболеваниями, сахарным диабетом 2-го типа, ретинопатией (невоспалительным поражением сетчатки глаза). Кроме того, люди с ожирением более склонны к развитию стеатоза печени, обструктивного апноэ во время сна (остановки дыхания), ортопедических осложнений.
Ген MC4R кодирует нейрональный меланокортиновый рецептор, который является фактором регуляции пищевого поведения (подавляющим аппетит) и энергетического баланса.
Экспрессия гена MC4R широко распространена в центральной нервной системе: таламусе, гипоталамусе, стволе мозга и коре. Распространение в этих регионах мозга рецепторов свидетельствует об их возможном участии в регуляции вегетативных и нейроэндокринных функций. Существуют данные, что MC4R экспрессируется также в спинном мозге.
Различные по степени тяжести формы ожирения, связанного с геном MC4R, являются следствием изменений последовательности гена, в результате чего белок – меланокортиновый рецептор – становится неактивен или менее активен. Тяжесть ожирения зависит от типа мутации и от того, находится ли она в гомозиготном или гетерозиготном состоянии.
Белок, кодируемый геном MC4R, формирует меланокортин, отвечающий за чувство насыщения. Соответственно, при мутации гена MC4R ребенок может переедать просто потому, что не способен контролировать аппетит по физиологическим причинам.
Ранняя диагностика причин ожирения позволяет существенно сократить его физические и психологические последствия. Родителям информация, что MC4R-ассоциированное ожирение у ребенка связано с тем, что у него проблемы с достижением сытости, указывает на необходимость с ранних лет регулировать пищевое поведение ребенка, следить за его массой тела.
В настоящее время несколько препаратов для целенаправленной терапии данного варианта ожирения проходят клинические испытания.
Генетическое исследование особенно важно для дифференциальной диагностики ожирения, вызванного мутацией гена MC4R, с другими формами ожирения; это необходимо для подбора адекватного метода терапии и определения прогноза заболевания.
Спектр мутаций в гене MC4R очень широк. Тест заключается в исследовании полной кодирующей области гена рецептора меланокортина 4 (MC4R) и позволяет выявить любые возможные мутации, приводящие к ожирению.
С помощью предлагаемого генетического исследования можно либо подтвердить недостаточность MC4R, либо исключить ее и перейти к поиску других причин, в том числе более редких форм наследственно обусловленного ожирения.
Конечно, даже наличие гена не означает, что у человека будет ожирение. Проявится ген или нет, зависит от образа жизни, пищевых привычек. Однако при любой наследственности ожирение легче предотвратить, чем потом избавляться от него.
Когда назначается исследование?
- При переедании у детей раннего возраста.
- При тяжелом ожирении с раннего возраста без нарушения полового и умственного развития.
- При дифференциальной диагностике с другими формами ожирения.
- Когда известно, что у родственников пациента было ожирение в раннем возрасте.
Что означают результаты?
Результатом генетического исследования является заключение о наличии или отсутствии мутаций гена MC4R;. выдается инф-ция о генотипе и наличии/отсутствии риска мутаций.
Скачать пример результатаТакже рекомендуется
- Ген, ассоциированный с жировой массой и ожирением (FTO). Выявление мутации G(45+52261)A (регуляторная область гена)
- Лептин
- Глюкоза в плазме
- Инсулин
- Гликированный гемоглобин (HbA 1c)
- Тиреотропный гормон (ТТГ)
- Тироксин свободный (Т4 свободный)
Литература
- Farooqi IS,et al. Clinical spectrum of obesity and mutations in the melanocortin 4 receptor gene. N Engl J Med 2003; 348: 1085−1095.
- Lubrano-Berthelier C. et al., Melanocortin 4 receptor mutations in a large cohort of severely obese adults: prevalence, functional classification, genotype-phenotype relationship, and lack of association with binge eating, J Clin Endocrinol Metab. 2006.
Ген окситоцинового рецептора оказался геном оптимизма
Исследователи из Калифорнийского университета в Лос-Анджелесе (США) обнаружили «ген оптимизма»: им оказался ген окситоцинового рецептора. Окситоцин известен как гормон материнства, но, кроме того, есть данные о его участии в формировании социального поведения человека: он улучшает социальные навыки, такие как эмпатия, с ним человек легче и лучше чувствует себя в компании. Что неудивительно, поскольку этот гормон выполняет также нейромедиаторную функцию.
Работа окситоцина напрямую зависит от наличия рецепторов к нему на поверхности клеток. Разные учёные уже сообщали, что ген этого рецептора существует в двух версиях: в определённом участке последовательности ДНК может стоять либо аденин (А-версия), либо гуанин (G-версия). Считается, что наличие того или иного варианта гена соответствует психологическому профилю человека: насколько он устойчив к стрессу, легко ли поддаётся унынию и т. п.
В нынешнем исследовании принимали участие 326 добровольцев, каждый из которых прошёл психологический тест. Психологи оценивали каждого участника по таким параметрам, как самооценка, оптимистичность характера, сила воли, подверженность депрессиям. После этого испытуемыми занялись молекулярные генетики: в их задачу входил генетический анализ образцов ДНК каждого участника эксперимента. Те, у кого в гене окситоцинового рецептора в определённом положении находились две или одна адениновые «буквы», проявляли бóльшую предрасположенность к депрессии. Те, у кого там были две G, лучше справлялись с подавленным психологическим состоянием.
Обладатели двух G в гене рецептора окситоцина отличались повышенной самооценкой, силой воли выше среднего и были склонны к оптимистичному взгляду на вещи. Всё это давало им психологический ресурс, за счёт которого можно было выйти из эмоционального кризиса. Таким образом, предрасположенность к депрессии и тоске может быть связана с тем, как именно нейроны воспринимают окситоцин, каким вариантом рецептора к нему они располагают.
Результаты своих исследований учёные представили к публикации в журнал PNAS.
Вместе с тем авторы работы предостерегают от упрощённого толкования своих результатов: дескать, если природа наградила тебя G-версией гена, то у тебя нет ни самооценки, ни оптимизма, и пребывать тебе вечно в депрессии. Окситоциновый рецептор никак не влияет на способность человека освоить некие психологические инструменты, позволяющие справиться со стрессом. Например, если знать о наличии «депрессивного» варианта гена у ребёнка, можно с самого раннего возраста приучать его обходить стороной стрессовые ситуации, объяснять, как с ними справляться, наконец, просто оказывать ему поддержку, чтобы депрессия не начала питать саму себя с детства. А о возможностях генетической терапии, которые неизбежно приходят в голову в таких случаях, пожалуй, лучше умолчать…
Подготовлено по материалам Калифорнийского университета в Лос-Анджелесе.
Источник: Компьюлента
Управление персоналом, образование, личное развитие. Тесты. Внимание. Память. IQ-тесты. Effecton Studio. Эффектон
У человека сильнее всего развиты зрение и слух, соответственно они и наиболее изучены, хотя есть и другие чувства, которые также важны для человека в его повседневной жизни.
Вибрационные ощущения
Со слуховыми ощущениями можно связать вибрационную чувствительность, т.к. у них общая природа отражаемых физических явлений. Вибрационные ощущения отражают колебания упругой среды. Этот вид чувствительности можно назвать «контактным слухом». Специальных вибрационных рецепторов у человека не обнаружено. Считается, что вибрационное чувство является одним из самых древних видов чувствительности, и отражать вибрации внешней и внутренней среды могут все ткани организма.
В жизни человека вибрационная чувствительность подчинятся слуховой и зрительной. Познавательное значение вибрационной чувствительности возрастает в тех видах деятельности, где вибрации становятся сигналом неисправностей в работе машины. В жизни глухих и слепоглухих вибрационная чувствительность компенсирует потерю слуха. Организм здорового человека непродолжительные вибрации тонизируют, длительные и интенсивные — утомляют и вызывают болезненные явления.
Обоняние
Рецептор обонятельных ощущений — это окончания обонятельного нерва в носовой полости, он относится к дистантным. Микроскопические частицы веществ, попадающие в носовую полость с воздухом, являясь раздражителями, вызывают обонятельные ощущения.
У животных обоняние — основной дистантный рецептор, ориентируясь по запаху, животное находит пищу или избегает опасности. Сексуальное поведение животных зависит от выработки особых веществ — феромонов. Существует теория, что и у людей феромоны играют не последнюю роль в вопросах пола.
Человеку в современном мире нет необходимости следовать обонятельным ощущениям, ориентируясь в окружающей среде. Функция обоняния у человека подавляется зрением и слухом. Отсутствие в языке специальных слов для обозначения обонятельных ощущений свидетельствует об их недостаточном развитии и нестойкости. Обычно говорят: «запах моря», «запах роз», «запах конюшни».
Обонятельная чувствительность тесно связана с вкусовой, помогает распознавать качества пищи. Обоняние предупреждает об опасной для организма воздушной среде, позволяет различать в ряде случаев химический состав веществ.
Вкус
Вкусовые ощущения — контактные, возникающие при соприкосновении органа чувств (языка) с самим предметом. Чувство вкуса обнаруживает молекулы, растворенные в слюне.
Существуют четыре основных качества вкусовых раздражителей: кислое, сладкое, горькое, соленое. Из комбинаций этих четырех ощущений, к которым присоединяются движения языка, и возникает комплекс вкусовых ощущений.
Вначале сенсорный процесс происходит во вкусовых сосочках, причем каждый из сосочков имеет от 50 до 150 рецепторных клеток, которые быстро изнашиваются от соприкосновения с пищей и затем обновляются. Затем сенсорные сигналы поступают по нервам в задний мозг, таламус и вкусовую кору, обрабатывающую вкусовые ощущения.
Вкусовые ощущения, как и обонятельные, повышают аппетит человека. Анализируя качество пищи, вкусовые ощущения выполняют также защитную функцию и важны для выживания. При голодании вкусовая чувствительность повышается, при насыщении или пресыщении — понижается.
Кожа
В кожных покровах имеется несколько самостоятельных анализаторных систем:
- тактильная (ощущения прикосновения),
- температурная,
- болевая.
Все виды кожной чувствительности относятся к контактной чувствительности. Наибольшее скопление тактильных клеток — на ладони, на кончиках пальцев и на губах. Кожные рецепторы передают информацию в спинной мозг, контактируя с двигательными нейронами, что делает возможным рефлекторные действия такие, как, например, как отдергивание руки от огня. Осязание — это тактильные ощущения руки вместе с мышечно-суставной чувствительностью.
Температурная чувствительность регулирует теплообмен между организмом и окружающей средой. Распределение тепловых и холодовых рецепторов по коже неравномерно. Наиболее чувствительна к холоду спина, наименее — грудь.
Сильное давление на поверхность тела вызывает болевое ощущение. Рецепторные окончания болевой чувствительности расположены под кожей, глубже, чем тактильные рецепторы. Там, где больше тактильных рецепторов болевых рецепторов меньше. Тактильная чувствительность дает знания о качествах предмета, а болевая чувствительность дает сигнал о вреде, наносимом раздражителем.
Проприоцептивная чувствительность
Кинестезия
Кинестезическими ощущениями называют ощущения движения и положения отдельных частей тела. Рецепторы кинестезических ощущений расположены в мышцах и сухожилиях. Раздражение в этих рецепторах возникает под влиянием растяжения и сокращения мышц.
Большое количество двигательных рецепторов расположено в пальцах рук, языке и губах, так как этими органами необходимо осуществлять точные и тонкие рабочие и речевые движения. Деятельность двигательного анализатора позволяет человеку координировать и контролировать свои движения. Упражнения для рук пакета «Комфорт» улучшают кровоснабжение, снижают напряжение и утомление, способствуя лучшей координации движений и повышению умственной работоспособности.
Понятно, что развитие кинестезических ощущений является одной из важнейших задач обучения.
Речевые кинестезии формируются в младенческом и дошкольном периодах развития человека. Обучение иностранному языку требует выработки таких речевых кинестезий, которые не характерны для родного языка.
Вестибулярное чувство
Статическая, или гравитационная, чувствительность отражает положение нашего тела в пространстве. Рецепторы ее расположены в вестибулярном аппарате внутреннего уха: полукружные каналы и вестибулярные мешочки преобразуют сигналы об относительном движении и силе тяжести и передают их в мозжечок и участок коры височной области. Резкие и частые изменения положения тела относительно плоскости земли, такие, как качание на качелях или морская качка приводят к головокружению — «морской болезни».
Достаточно ли у человека органов чувств?
Ощущения обеспечивают организму адекватную ориентировку в окружающей среде. Удалось ли человеку познать окружающий мир глубже, если бы он имел больше органов чувств?
Философы-идеалисты делали вывод об ограниченности познавательных возможностей человека, связывая это с ограниченностью органов чувств и разнообразием явлений в окружающем мире.
Материалисты считали, что имеющихся органов чувств достаточно для полного познания мира. Познание идет вглубь, познавательная сила человека состоит в том, что к деятельности его органов чувств прибавляется деятельность мышления, которое раздвигает рамки познавательных возможностей.
Эксклюзивный материал сайта «www.effecton.ru — психологические тесты и коррекционные программы». Заимствование текста и/или связанных материалов возможно только при наличии прямой и хорошо различимой ссылки на оригинал. Все права защищены.
Соматические мутации в гене рецептора к андрогенам как причина возникновения синдрома резистентности к андрогенам | Калинченко
Актуальность
Синдром резистентности к андрогенам (СРА) или синдром тестикулярной феминизации (СТФ) (MIM 300681/312300) – патологическое состояние, характеризующееся полной или частичной нечувствительностью тканей к андрогенам. СРА является одной из наиболее частых причин нарушения формирования пола при кариотипе 46,XY (НФП 46,XY). Частота встречаемости данной патологии, по данным литературы, составляет примерно 1:20 000–40 000 новорожденных с кариотипом 46,XY [1].
СРА обусловлен мутацией в гене рецептора андрогенов (AR), расположенном на длинном плече X-хромосомы (Xq11-12) и состоящем из 8 экзонов. Белок AR принадлежит к семейству внутриклеточных рецепторов, локализующихся в цитоплазме, и включает лигандсвязывающий домен, ДНК-связывающий домен и домен, активирующий транскрипцию гена-мишени. После взаимодействия с лигандом рецептор приобретает дополнительное сродство к ядерным акцепторным структурам и проникает внутрь ядра, где активирует транскрипцию генов-мишеней. В настоящее время известно более 1000 различных патологических вариантов гена AR. В 70% случаев обнаруживаемые мутации наследуются от матери, являющейся носителем мутантного гена. В 30% случаев патологические замены у пациента возникают de novo.
В зависимости от степени потери активности AR клинические проявления заболевания варьируют от правильного женского фенотипа при рождении (так называемая полная форма синдрома резистентности к андрогенам (пСРА)) до неполной маскулинизации наружных половых органов (НПО) – неполной формы СРА (нпСРА). Также выделяют мягкую форму СРА (мСРА), которая диагностируется у взрослых мужчин при обследовании по поводу бесплодия или истиной гинекомастии. Соответственно у пациентов последней группы строение половых органов остается нормальным [2, 3]. СРА относится к X-сцепленным заболеваниям, при которых женщина является носителем патологического варианта AR, тогда как рожденные от нее мальчики (46,XY) в 50% случаев будут больными, а девочки (46,XX) в 50% окажутся гетерозиготными носителями, как и мать.
В настоящее время всем пациентам с подозрением на СРА проводится молекулярно-генетический анализ гена AR и при выявлении мутации рекомендуется молекулярно-генетическое обследование матери с целью проведения медико-генетического консультирования пары. Однако в литературе имеются единичные описания случаев выявления гетерозиготной мутации при секвенировании гена AR у пациента с 46,XY и подозрением на СРА. Этот редкий феномен для заболеваний с Х-сцепленным типом наследования возможен лишь в случае возникновения мутации на постзиготном этапе развития плода [4]. Поскольку патологические изменения в гене AR на постзиготном уровне могут происходить на любом этапе деления зиготы, фенотип пациентов значительно варьирует. Более того, в ряде случаев возможна значимая вирилизация пациентов в период пубертата, что объясняется незначительным присутствием мутантного варианта AR в клетках [4].
При классическом X-сцепленном типе наследования СРА рекомендуется выбор женского пола воспитания. Однако при СРА с гетерозиготными мутациями выбор тактики более сложен, учитывая возможную удовлетворительную маскулинизацию в период полового созревания.
Ниже приводится описание фенотипических, гормональных и молекулярно-генетических особенностей 8 пациентов с различными формами СРА, у которых выявлены гетерозиготные мутации в гене AR. У матерей пробандов изменений в этом гене выявлено не было, что позволило исключить наследование выявленных мутаций.
Описание случаев
Пациент 1. При рождении – НПО по женскому типу, но в половых губах обнаружены образования, напоминающие тестикулы; по данным УЗИ, структура этих образований схожа с яичками, кровоток в норме. Зарегистрирована в женском поле. Кариотип 46,XY. В возрасте 3 мес при МРТ органов малого таза в структуре больших половых губ визуализированы гонады размером до 14×9 мм справа и 9×5 мм слева, матка и яичники отсутствуют. В гормональном профиле: ЛГ– 0,4 мМЕ/мл; ФСГ – 1,13 мМЕ/мл; тестостерон – 0,88 нг/мл; эстрадиол – 11,0 пг/мл. Заподозрен диагноз «Синдром резистентности к андрогенам, полная форма». В возрасте 6 лет: женский фенотип, мошонкообразные половые губы, яички пальпаторно определяются в паховой области с двух сторон; при пробе с ХГЧ – уровень тестостерона – 12,5 нмоль/л (см. таблицу). При секвенировании по Сэнгеру в гене AR выявлен ранее не описанный гетерозиготный вариант с.2276_2290 delACTCCAGGATGCTCT: p.N759_Y764del. С учетом психосоциальной адаптации к женскому полу и расположения гонад проведена гонадэктомия.
Клинические, гормональные и молекулярно-генетические характеристики пациентов
Пациент | Пол воспитания | Форма | Фенотип | Тестостерон в ходе пробы с ХГЧ, нмоль/л | Гонадэктомия | Мутация в AR |
1 | ж | Полная | Женский, в половых губах гонады | 12,5 (6 лет) | В 6 лет | c.2276_2290del:p.759_764del |
2 | ж | Полная | Женский | – | В 4 года | c.2107T>C:p.C703R |
3 | м | Неполная | Гонады в мошонке, промежностная форма гипоспадии | 18 (11 мес) | c.2426A>C:p.K809T | |
4 | ж | Полная | Женский | 11,3 (6 лет) | В 6 лет | c.2599G>A:p.V867M |
5 | м/ж | Неполная | Гонады в мошонке, промежностная форма гипоспадии | 41,1 (2 мес) | c.1018G>T:p.E340X | |
6 | м | Неполная | Мошоночная гипоспадия, гонады в мошонке | – | c.2395C>G:p.Q799E. | |
7 | м/ж | Неполная | Мошоночная гипоспадия, гонады в мошонке, микропенис | Нет | c.2635T>G:p.F879V | |
8 | ж | Полная | Женский | 15 (5 лет) | В 5 лет | с.3458G>A:p.M781I |
Примечание. м/ж – смена паспортного пола с мужского на женский.
Пациент 2. При рождении – НПО по женскому типу, зарегистрирована в женском поле. В 4 года при проведении герниопластики по поводу двусторонних паховых грыж в грыжевых мешках выявлены тестикулы, которые были удалены. При дообследовании установлен кариотип 46,XY. При молекулярно-генетическом исследовании с использованием панели НФП 46,XY в гене AR выявлена гетерозиготная мутация c.2107T>C:p.C703R, описанная ранее при пСРА.
Пациент 3. При рождении промежностная гипоспадия, яички в расщепленной мошонке с двух сторон, микропенис. Зарегистрирован в мужском поле. Кариотип 46,XY. При пробе с ХГЧ в возрасте 11 мес уровень тестостерона составил 18,4 нмоль/л, что позволило предположить нпСРА. Молекулярно-генетическое исследование с использованием панели НФП 46,XY обнаружило ранее не описанный гетерозиготный вариант гена AR: c.2426A>C:p.K809T. Строение НПО (2–3 по шкале Прадера) подтверждало диагноз нпСРА. От смены пола воспитания родители отказались. В возрасте 1 года проведен первый этап маскулинизирующей пластики.
Пациент 4. При рождении – строение НПО по женскому типу. В возрасте 9 мес при проведении герниопластики по поводу двусторонней паховой грыжи в паховых каналах обнаружены гонады, которые были сохранены и перемещены в брюшную полость. Кариотип 46,XY. В возрасте 6 лет: НПО сформированы правильно, по женскому типу; в паховой области справа пальпируется яичко объемом 2 мл. При пробе с ХГЧ уровень тестостерона 11,3 нмоль/л (см. таблицу). Учитывая высокий подъем уровня тестостерона, заподозрена полная форма СРА. При секвенировании по Сэнгеру в гене AR выявлен гетерозиготный вариант c.2599G>A:p.V867M. Аналогичная гетерозиготная мутация была описана ранее при нпСРА [5]; в гемизиготном состоянии данная мутация описана как при пСРА, так и при нпСРА [6, 7]. Учитывая психосоциальную адаптацию в женском поле, проведена гонадэктомия.
Пациент 5. При рождении – неправильное строение НПО (микропенис до 1 см, головка не сформирована, кавернозные тела слабо развиты, промежностная форма гипоспадии, яички с обеих сторон у входа в мошонку). Кариотип 46,ХY. Пациент зарегистрирован в мужском поле. В возрасте 2 мес в гормональном профиле: ЛГ 1,39 МЕд/л (норма 0–1,5), ФСГ – 0,66 МЕд/л (0–2), тестостерон – 1,94 нмоль/л. При пробе с ХГЧ – подъем уровня тестостерона до 41 нмоль/л. Заподозрен диагноз «СРА, неполная форма». При секвенировании по Сэнгеру в гене AR выявлен гетерозиготный вариант c.1018G>T:р.E340X, описанный ранее при пСРА. Из-за невозможности достижения адекватной маскулинизации рекомендована адаптация ребенка в женском поле.
Пациент 6. При рождении – неправильное строение НПО: мошоночная форма гипоспадии, половой член до 2 см, искривлен, кавернозные тела развиты слабо, гонады в мошонке. На основании электролитных нарушений (гипонатриемия до 118 ммоль/л, гиперкалиемия до 6 ммоль/л) и данных гормонального анализа (тестостерон 0,1 нмоль/л, 17ОНР 2,8 нг/мл при норме 0,1–1,7 нг/мл) установлен диагноз «Врожденная дисфункция коры надпочечников, сольтеряющая форма». Однако при динамическом наблюдении всегда отмечались низкие уровни 17ОНР и АКТГ. В возрасте 2,5 года при пробе с АКТГ («Синактен-Депо») выброс кортизола составил 1250 нмоль/л, что позволило исключить любые формы первичной надпочечниковой недостаточности. Для уточнения причины неправильного строения НПО проведено молекулярно-генетическое исследование с использованием панели НФП 46,XY и в гене AR выявлен описанный ранее гетерозиготный вариант c.2395C>G:p.799E. Учитывая строение НПО (2–3 по шкале Прадера), подтверждающее диагноз нпСРА, с родителями была проведена беседа о возможности смены пола воспитания ребенка на женский. От смены пола родители отказались.
Пациент 7. При рождении – неправильное строение НПО (мошоночная форма гипоспадии, гонады в расщепленной мошонке, головка искривленного полового члена сформирована хорошо, кавернозные тела удовлетворительно развиты). При неонатальном скрининге выявлено умеренное повышение уровня 17ОНР (до 40 нмоль/л) и была заподозрена ВДКН. В возрасте 2 мес мультистероидный анализ не выявил нарушений стероидогенеза, уровень 17ОНР соответствовал возрастной норме. Молекулярно-генетическое исследование с использованием панели НФП 46,XY обнаружило в гене AR гетерозиготную мутацию c.2635T>G:p.F879V, описанную ранее при нпСРА. Для оценки чувствительности тканей к андрогенам был проведен курс лечения смесью эфиров тестостерона в дозе 50 мг 1 раз в 28 дней внутримышечно в течение 3 мес. На фоне терапии отмечено умеренное увеличение полового члена и мошонки, появление ее складчатости. Учитывая достаточную степень маскулинизации НПО при рождении и положительную реакцию на лечение эфирами тестостерона, рекомендовано воспитание ребенка в мужском паспортном поле.
Пациент 8. При рождении – правильное строение НПО по женскому типу; в 2 года при проведении герниопластики по поводу двусторонних паховых грыж выявлены и удалены тестикулы. Кариотип 46,XY. В возрасте 15 лет обращение к эндокринологу с жалобами на отсутствие вторичных половых признаков. Идентифицирует себя в женском поле, психологически адаптирована. На момент обследования: ЛГ– 32 МЕД/л (2,4–12,6), ФСГ – 65 МЕД/л (3,5–12,5). Учитывая отсутствие гонад, дальнейшее гормональное обследование не проводилось. При молекулярно-генетическом исследовании с использованием панели НФП 46,XY в гене AR выявлена описанная ранее гетерозиготная мутация с.3458G>A, p.M781I. С целью развития вторичных половых признаков назначена заместительная терапия эстрогенами. В настоящее время пациентка замужем, воспитывает приемного ребенка.
Обсуждение
Хотя СРА является самой частой причиной НФП 46,XY, именно этот синдром остается наиболее сложным в плане выбора пола воспитания, особенно при нпСРА. Если при пСРА (учитывая правильный женский фенотип при рождении) выбор в пользу женского пола однозначен, то при нпСРА в большинстве случаев выбор пола воспитания определяется решением медицинского персонала, принимающего роды. У пациентов с нпСРА, особенно при строении НПО ближе к феминному (2–3 по шкале Прадера), выбор в пользу мужского пола неизбежно приведет к развитию истинной гинекомастии в период полового созревания и обусловит высокий риск малигнизации тестикул [8]. Несмотря на терапию супрафизиологическими дозами половых стероидов, достаточное развитие вторичных половых признаков (рост волос в андрогензависимых зонах, изменение голоса, увеличение половых органов) в этих случаях невозможно. Поэтому если при рождении выбран мужской пол, то при верификации СРА рекомендуется смена паспортного пола на женский, что предсказуемо приводит к психологической травме в семье и нередко к отказу родителей от смены пола ребенку по социальным причинам. В связи с этим максимально ранняя диагностика СРА является неотъемлемой частью правильного лечения пациентов данной группы.
Молекулярно-генетическая диагностика СРА позволяет с высокой достоверностью устанавливать диагноз [9], но отсутствие наиболее распространенных мутаций препятствует разработке методов быстрого установления диагноза. Тем не менее имеются немногочисленные публикации о выявлении гетерозиготных мутаций в гене AR у данной группы пациентов. Так, в 1993 г. О. Hiort и соавт. [7] впервые описали пациента с нпСРА, у которого была выявлена соматическая мутация, приводящая к функционально значимой замене глутамина на гистидин в кодоне 733. Изначально эта мутация была расценена как гемизиготная, однако авторы обратили внимание на сохранение у пациента частичной чувствительности к андрогенам. Рестрикционный анализ выявил гетерозиготный вариант мутации; наличие клеток с функционально активным рецептором могло объяснить несоответствие тяжести мутации фенотипу. В 1997 г. аналогичное наблюдение было опубликовано A. Boehmer и соавт. [10]: у пациента с нпСРА была выявлена замена TTA на TGA в 172-м кодоне, приведшая к образованию стоп-кодона. Несоответствие клинической картины тяжести мутации заставило повторно проанализировать данные секвенирования; при этом было обнаружено ранее пропущенное «необычное» изменение – наличие мутации в гетерозиготном состоянии.
В 2005 г. B. Kohler и др. [11] в обзорной статье (7 пациентов с соматическими мутациями) впервые описали 3 случая пСРА. Аналогичные 4 случая пСРА при соматических мутациях в гене AR представлены и нами. Надежное объяснение данного феномена до настоящего времени отсутствует. Возможно, генетическое исследование, проводимое на клетках крови, не учитывает распределение клеток в других тканях. Теоретически исследование экспрессии рецептора андрогенов в удаленных гонадах могло бы подтвердить или опровергнуть данную гипотезу. Следует учитывать и то обстоятельство, что при проведении генетического анализа оценивается постнатальное состояние органогенеза, тогда как внутриутробно, в период формирования НПО, число клеток с мутантным типом AR может значительно превосходить количество клеток с диким типом AR. В процессе дифференцировки клеток клетки с мутантным типом рецептора могли быть менее жизнеспособными, что приводило бы к изменению соотношения в пользу клеток с нормальным AR [11].
Существование различных фенотипов при соматических мутациях требует персонализированного подхода к пациентам с целью правильного выбора пола воспитания и срока проведения гонадэктомии. Последнее важно учитывать при выборе женского пола, так как при достаточном количестве клеток с функционально активными рецепторами в период полового созревания может происходить спонтанная маскулинизация [4]. Это же позволяет воспитывать часть пациентов с СРА в мужском поле.
Выявление соматического мозаицизма при СРА также важно для медико-генетического консультирования семьи. Хотя заболевание является X-сцепленным, отсутствие мозаицизма у матери позволяет исключить риск рождения еще одного сибса с данной патологией.
Заключение
Хотя СРА относят к Х-сцепленным заболеваниям, у ряда пациентов изменения в AR могут возникать на постзиготном этапе, приводя к гетерозиготным мутациям. В зависимости от времени возникновения мутации соотношение клеток с функционально активным и неактивным AR у пациентов может быть разным, что обусловливает выраженный фенотипический полиморфизм и сложность прогнозирования течения заболевания в пубертатном периоде. Накопление клинического опыта, возможно, позволит выработать определенный алгоритм обследования и лечения таких пациентов.
Дополнительная информация
Источник финансирования. Работа выполнена при содействии Фонда поддержки и развития филантропии «КАФ».
Согласие пациентов. Законные представители пациентов добровольно подписали информированное согласие на публикацию персональных медицинских сведений в обезличенной форме в журнале «Проблемы эндокринологии».
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией настоящей статьи.
Участие авторов. Концепция и дизайн исследования, сбор материала, анализ полученных данных, написание текста – Н.Ю. Калинченко, А.Н. Тюльпаков; сбор материала, анализ полученных данных – А.А. Колодкина; проведение молекулярно-генетического исследования – В.М. Петров, Е.В. Васильев, А.Н. Тюльпаков. Все авторы внесли существенный вклад в проведение работы и подготовку статьи, прочли и одобрили итоговую версию до публикации.
1. Bangsboll S, Qvist I, Lebech PE, Lewinsky M. Testicular feminization syndrome and associated gonadal tumors in Denmark. Acta Obstet Gynecol Scand. 1992;71(1):63–66. doi: https://doi.org/10.3109/0001634920900795
2. Gottlieb B, Lombroso R, Beitel LK, Trifiro MA. Molecular pathology of the androgen receptor in male (in)fertility. Reprod Biomed Online. 2005;10(1):42–48. doi: https://doi.org/10.1016/s1472-6483(10)60802-4
3. Hughes IA, Werner R, Bunch T, Hiort O. Androgen insensitivity syndrome. Semin Reprod Med. 2012;30(5):432–442. doi: https://doi.org/10.1055/s-0032-1324728
4. Gottlieb B, Beitel LK, Trifiro MA. Somatic mosaicism and variable expressivity. Trends Genet. 2001;17(2):79–82. doi: https://doi.org/10.1016/s0168-9525(00)02178-8
5. Hiort O, Sinnecker GH, Holterhus PM, et al. Inherited and de novo androgen receptor gene mutations: investigation of single-case families. J Pediatrics. 1998;132(6):939–943. doi: https://doi.org/10.1016/s0022-3476(98)70387-7
6. Lubahn DB, Brown TR, Simental JA, et al. Sequence of the intron/exon junctions of the coding region of the human androgen receptor gene and identification of a point mutation in a family with complete androgen insensitivity. Proc Natl Acad Sci USA. 1989;86(23):9534–9538. doi: https://doi.org/10.1073/pnas.86.23.9534
7. Hiort O, Huang Q, Sinnecker GH, et al. Single strand conformation polymorphism analysis of androgen receptor gene mutations in patients with androgen insensitivity syndromes: application for diagnosis, genetic counseling, and therapy. J Clin Endocrinol Metab. 1993;77(1):262–266. doi: https://doi.org/10.1210/jcem.77.1.8325950
8. Rutgers JL, Scully RE. The androgen insensitivity syndrome (testicular feminization): a clinicopathologic study of 43 cases. Int J Gynecol Pathol. 1991;10(2):126–144. doi: https://doi.org/10.1097/00004347-199104000-00002
9. Quigley CA, De Bellis A, Marschke KB, et al. Androgen receptor defects: historical, clinical, and molecular perspectives. Endocr Rev. 1995;16(3):271–321. doi: https://doi.org/10.1210/edrv-16-3-271
10. Boehmer AL, Brinkmann AO, Niermeijer MF, et al. Germ-line and somatic mosaicism in the androgen insensitivity syndrome: implications for genetic counseling. Am J Hum Genet. 1997;60(4):1003–1006.
11. Kohler B, Lumbroso S, Leger J, et al. Androgen insensitivity syndrome: somatic mosaicism of the androgen receptor in seven families and consequences for sex assignment and genetic counseling. J Clin Endocrinol Metab. 2005;90(1):106–111. doi: https://doi.org/10.1210/jc.2004-0462
Сенсорные рецепторы | Психология Вики
Оценка |
Биопсихология |
Сравнительный |
Познавательная |
Развивающий |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клиническая |
Образовательная |
Промышленное |
Профессиональные товары |
Мировая психология |
Биологический: Поведенческая генетика · Эволюционная психология · Нейроанатомия · Нейрохимия · Нейроэндокринология · Неврология · Психонейроиммунология · Физиологическая психология · Психофармакология (Указатель, Схема)
В сенсорной системе сенсорный рецептор — это структура, которая распознает стимул во внутренней или внешней среде организма.В ответ на стимулы сенсорный рецептор инициирует сенсорную трансдукцию, создавая градиентные потенциалы или потенциалы действия в той же или соседней клетке.
Содержание
- 1 Расположение
- 2 функции
- 3 Классификация по адекватному стимулу
- 4 Классификация по местонахождению
- 5 Классификация по морфологии
- 6 См. Также
- 7 Артикул
Сенсорный рецептор может быть специализированной частью плазматической мембраны или отдельной клеткой, связанной с окончанием нейрона.
Сенсорные рецепторы, отвечающие за вкус и запах, содержат рецепторы, которые связываются с определенными химическими веществами. Рецепторы запаха в нейронах обонятельных рецепторов, например, активируются путем взаимодействия с молекулярными структурами молекулы запаха. Точно так же вкусовые рецепторы (вкусовые рецепторы) во вкусовых рецепторах взаимодействуют с химическими веществами в пище, создавая потенциал действия.
Другие рецепторы, такие как механорецепторы и фоторецепторы, реагируют на физические раздражители. Например, фоторецепторные клетки содержат специализированные белки, такие как родопсин, для преобразования физической энергии света в электрические сигналы.Некоторые типы механорецепторов активируют потенциалы действия при физическом растяжении их мембран.
Сенсорный рецептор функционирует как первый компонент сенсорной системы.
Сенсорные рецепторы реагируют на определенные стимулы. Модальность стимула, на которую реагирует сенсорный рецептор, определяется адекватным стимулом сенсорного рецептора.
Сенсорный рецептор реагирует на свои стимулы, инициируя сенсорную трансдукцию.
Классификация по адекватному стимулу [править | править источник]
Адекватный стимул сенсорного рецептора — это стимул, для которого он обладает адекватным аппаратом сенсорной трансдукции.Адекватный стимул можно использовать для классификации сенсорных рецепторов:
- Барорецепторы реагируют на давление
- Хеморецепторы реагируют на химические раздражители
- Электромагнитные рецепторы реагируют на инфракрасное излучение, видимый свет или магнитные поля. [1]
- Гидрорецепторы реагируют на изменение влажности
- Механорецепторы реагируют на механическое напряжение или механическое напряжение
- Ноцицепторы реагируют на вредные или потенциально вредные раздражители
- Осморецепторы реагируют на осмолярность жидкости (например, в гипоталамусе
- Фоторецепторы реагируют на свет
- Проприоцепторы обеспечивают чувство положения
- Терморецепторы реагируют на температуру, либо тепло, либо холод, либо и то, и другое.
Классификация по местоположению [править | править источник]
Сенсорные рецепторы можно классифицировать по местонахождению:
- Кожные рецепторы — это сенсорные рецепторы, обнаруженные в дерме или эпидермисе.
- Мышечные веретена содержат механорецепторы, определяющие растяжение мышц.
Классификация по морфологии [править | править источник]
Соматические сенсорные рецепторы у поверхности кожи обычно можно разделить на группы в зависимости от морфологии:
- Свободные нервные окончания характеризуют ноцицепторы и терморецепторы и называются так, потому что терминальные ветви нейрона немиелинизированы и распространяются по дерме и эпидермису.
- Инкапсулированные рецепторы включают остальные типы кожных рецепторов. Инкапсуляция существует для специализированного функционирования.
- Перехватчики
- Нейроанализаторы
- Проприоцепция
- Sense
- Сенсорный нейрон
- ↑ Майкл Дж. Грегори. Сенсорные системы. Общественный колледж Клинтона. URL-адрес, доступ осуществлен 06.06.2013.
| Нервная система — Сенсорная система — редактировать |
|---|
| Особые чувства : Визуальная система | Слуховая система | Обонятельная система | Вкусовая система Соматосенсорная система : Ноцицепция | Терморецепция | Вестибулярная система | Механорецепция (давление, вибрация и проприоцепция) | Эквилибриоцепция |
| На этой странице используется содержимое Википедии под лицензией Creative Commons (просмотреть авторов). |
Сенсорные рецепторы | Психология Вики
Оценка |
Биопсихология |
Сравнительный |
Познавательная |
Развивающий |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клиническая |
Образовательная |
Промышленное |
Профессиональные товары |
Мировая психология |
Биологический: Поведенческая генетика · Эволюционная психология · Нейроанатомия · Нейрохимия · Нейроэндокринология · Неврология · Психонейроиммунология · Физиологическая психология · Психофармакология (Указатель, Схема)
В сенсорной системе сенсорный рецептор — это структура, которая распознает стимул во внутренней или внешней среде организма.В ответ на стимулы сенсорный рецептор инициирует сенсорную трансдукцию, создавая градиентные потенциалы или потенциалы действия в той же или соседней клетке.
Содержание
- 1 Расположение
- 2 функции
- 3 Классификация по адекватному стимулу
- 4 Классификация по местонахождению
- 5 Классификация по морфологии
- 6 См. Также
- 7 Артикул
Сенсорный рецептор может быть специализированной частью плазматической мембраны или отдельной клеткой, связанной с окончанием нейрона.
Сенсорные рецепторы, отвечающие за вкус и запах, содержат рецепторы, которые связываются с определенными химическими веществами. Рецепторы запаха в нейронах обонятельных рецепторов, например, активируются путем взаимодействия с молекулярными структурами молекулы запаха. Точно так же вкусовые рецепторы (вкусовые рецепторы) во вкусовых рецепторах взаимодействуют с химическими веществами в пище, создавая потенциал действия.
Другие рецепторы, такие как механорецепторы и фоторецепторы, реагируют на физические раздражители. Например, фоторецепторные клетки содержат специализированные белки, такие как родопсин, для преобразования физической энергии света в электрические сигналы.Некоторые типы механорецепторов активируют потенциалы действия при физическом растяжении их мембран.
Сенсорный рецептор функционирует как первый компонент сенсорной системы.
Сенсорные рецепторы реагируют на определенные стимулы. Модальность стимула, на которую реагирует сенсорный рецептор, определяется адекватным стимулом сенсорного рецептора.
Сенсорный рецептор реагирует на свои стимулы, инициируя сенсорную трансдукцию.
Классификация по адекватному стимулу [править | править источник]
Адекватный стимул сенсорного рецептора — это стимул, для которого он обладает адекватным аппаратом сенсорной трансдукции.Адекватный стимул можно использовать для классификации сенсорных рецепторов:
- Барорецепторы реагируют на давление
- Хеморецепторы реагируют на химические раздражители
- Электромагнитные рецепторы реагируют на инфракрасное излучение, видимый свет или магнитные поля. [1]
- Гидрорецепторы реагируют на изменение влажности
- Механорецепторы реагируют на механическое напряжение или механическое напряжение
- Ноцицепторы реагируют на вредные или потенциально вредные раздражители
- Осморецепторы реагируют на осмолярность жидкости (например, в гипоталамусе
- Фоторецепторы реагируют на свет
- Проприоцепторы обеспечивают чувство положения
- Терморецепторы реагируют на температуру, либо тепло, либо холод, либо и то, и другое.
Классификация по местоположению [править | править источник]
Сенсорные рецепторы можно классифицировать по местонахождению:
- Кожные рецепторы — это сенсорные рецепторы, обнаруженные в дерме или эпидермисе.
- Мышечные веретена содержат механорецепторы, определяющие растяжение мышц.
Классификация по морфологии [править | править источник]
Соматические сенсорные рецепторы у поверхности кожи обычно можно разделить на группы в зависимости от морфологии:
- Свободные нервные окончания характеризуют ноцицепторы и терморецепторы и называются так, потому что терминальные ветви нейрона немиелинизированы и распространяются по дерме и эпидермису.
- Инкапсулированные рецепторы включают остальные типы кожных рецепторов. Инкапсуляция существует для специализированного функционирования.
- Перехватчики
- Нейроанализаторы
- Проприоцепция
- Sense
- Сенсорный нейрон
- ↑ Майкл Дж. Грегори. Сенсорные системы. Общественный колледж Клинтона. URL-адрес, доступ осуществлен 06.06.2013.
| Нервная система — Сенсорная система — редактировать |
|---|
| Особые чувства : Визуальная система | Слуховая система | Обонятельная система | Вкусовая система Соматосенсорная система : Ноцицепция | Терморецепция | Вестибулярная система | Механорецепция (давление, вибрация и проприоцепция) | Эквилибриоцепция |
| На этой странице используется содержимое Википедии под лицензией Creative Commons (просмотреть авторов). |
Сенсорные рецепторы | Психология Вики
Оценка |
Биопсихология |
Сравнительный |
Познавательная |
Развивающий |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клиническая |
Образовательная |
Промышленное |
Профессиональные товары |
Мировая психология |
Биологический: Поведенческая генетика · Эволюционная психология · Нейроанатомия · Нейрохимия · Нейроэндокринология · Неврология · Психонейроиммунология · Физиологическая психология · Психофармакология (Указатель, Схема)
В сенсорной системе сенсорный рецептор — это структура, которая распознает стимул во внутренней или внешней среде организма.В ответ на стимулы сенсорный рецептор инициирует сенсорную трансдукцию, создавая градиентные потенциалы или потенциалы действия в той же или соседней клетке.
Содержание
- 1 Расположение
- 2 функции
- 3 Классификация по адекватному стимулу
- 4 Классификация по местонахождению
- 5 Классификация по морфологии
- 6 См. Также
- 7 Артикул
Сенсорный рецептор может быть специализированной частью плазматической мембраны или отдельной клеткой, связанной с окончанием нейрона.
Сенсорные рецепторы, отвечающие за вкус и запах, содержат рецепторы, которые связываются с определенными химическими веществами. Рецепторы запаха в нейронах обонятельных рецепторов, например, активируются путем взаимодействия с молекулярными структурами молекулы запаха. Точно так же вкусовые рецепторы (вкусовые рецепторы) во вкусовых рецепторах взаимодействуют с химическими веществами в пище, создавая потенциал действия.
Другие рецепторы, такие как механорецепторы и фоторецепторы, реагируют на физические раздражители. Например, фоторецепторные клетки содержат специализированные белки, такие как родопсин, для преобразования физической энергии света в электрические сигналы.Некоторые типы механорецепторов активируют потенциалы действия при физическом растяжении их мембран.
Сенсорный рецептор функционирует как первый компонент сенсорной системы.
Сенсорные рецепторы реагируют на определенные стимулы. Модальность стимула, на которую реагирует сенсорный рецептор, определяется адекватным стимулом сенсорного рецептора.
Сенсорный рецептор реагирует на свои стимулы, инициируя сенсорную трансдукцию.
Классификация по адекватному стимулу [править | править источник]
Адекватный стимул сенсорного рецептора — это стимул, для которого он обладает адекватным аппаратом сенсорной трансдукции.Адекватный стимул можно использовать для классификации сенсорных рецепторов:
- Барорецепторы реагируют на давление
- Хеморецепторы реагируют на химические раздражители
- Электромагнитные рецепторы реагируют на инфракрасное излучение, видимый свет или магнитные поля. [1]
- Гидрорецепторы реагируют на изменение влажности
- Механорецепторы реагируют на механическое напряжение или механическое напряжение
- Ноцицепторы реагируют на вредные или потенциально вредные раздражители
- Осморецепторы реагируют на осмолярность жидкости (например, в гипоталамусе
- Фоторецепторы реагируют на свет
- Проприоцепторы обеспечивают чувство положения
- Терморецепторы реагируют на температуру, либо тепло, либо холод, либо и то, и другое.
Классификация по местоположению [править | править источник]
Сенсорные рецепторы можно классифицировать по местонахождению:
- Кожные рецепторы — это сенсорные рецепторы, обнаруженные в дерме или эпидермисе.
- Мышечные веретена содержат механорецепторы, определяющие растяжение мышц.
Классификация по морфологии [править | править источник]
Соматические сенсорные рецепторы у поверхности кожи обычно можно разделить на группы в зависимости от морфологии:
- Свободные нервные окончания характеризуют ноцицепторы и терморецепторы и называются так, потому что терминальные ветви нейрона немиелинизированы и распространяются по дерме и эпидермису.
- Инкапсулированные рецепторы включают остальные типы кожных рецепторов. Инкапсуляция существует для специализированного функционирования.
- Перехватчики
- Нейроанализаторы
- Проприоцепция
- Sense
- Сенсорный нейрон
- ↑ Майкл Дж. Грегори. Сенсорные системы. Общественный колледж Клинтона. URL-адрес, доступ осуществлен 06.06.2013.
| Нервная система — Сенсорная система — редактировать |
|---|
| Особые чувства : Визуальная система | Слуховая система | Обонятельная система | Вкусовая система Соматосенсорная система : Ноцицепция | Терморецепция | Вестибулярная система | Механорецепция (давление, вибрация и проприоцепция) | Эквилибриоцепция |
| На этой странице используется содержимое Википедии под лицензией Creative Commons (просмотреть авторов). |
Введение в Sensation | Безграничная психология
Знакомство с Sensation
Ощущение включает передачу информации от сенсорных рецепторов в мозг и позволяет человеку познавать окружающий мир.
Цели обучения
Объясните, как мозг и сенсорные рецепторы работают вместе в процессе ощущения
Основные выводы
Ключевые моменты
- Ощущения — это входные данные о физическом мире, регистрируемые нашими сенсорными рецепторами, такими как глаза, уши, рот, нос и кожа.
- Восприятие — это процесс, с помощью которого мозг выбирает, организует и интерпретирует ощущения; на него часто влияют обучение, память, эмоции и ожидания.
- Чувства человека включают зрение, звук, вкус, запах и осязание, а также кинестезию и вестибулярные органы чувств.
- Информация, поступающая от наших органов чувств, поступает через сенсорные рецепторы тела, которые затем преобразуют входящую энергию в нервные импульсы. Эти нервные импульсы попадают в кору головного мозга, где они интерпретируются и организуются в процессе восприятия.
Ключевые термины
- рецептор : Любая специализированная клетка или структура, которая реагирует на сенсорные стимулы.
- кинестезия : проприоцепция или ощущение статической позиции; восприятие положения и осанки тела; также, в более широком смысле, включая движение тела.
- вестибулярный : относящийся к полости тела или относящийся к ней.
Ощущение и восприятие — два отдельных процесса, которые очень тесно связаны.Ощущения — это информация о физическом мире, полученная нашими сенсорными рецепторами, а восприятие — это процесс, с помощью которого мозг выбирает, организует и интерпретирует эти ощущения. Другими словами, чувства — это физиологическая основа восприятия. Восприятие одних и тех же органов чувств может варьироваться от одного человека к другому, потому что мозг каждого человека интерпретирует стимулы по-разному в зависимости от обучения, памяти, эмоций и ожиданий этого человека.
Чувства
Существует пять классических человеческих чувств: зрение, звук, вкус, запах и осязание.Два других чувства, кинестезия и вестибулярные чувства, получили широкое признание ученых. Кинестезия — это восприятие положения частей тела, обычно известное как «осознание тела». Вестибулярные органы чувств определяют силу тяжести, линейное ускорение (например, ускорение или замедление на прямой дороге) и вращательное ускорение (например, ускорение или замедление на повороте). И кинестезия, и вестибулярные органы чувств помогают нам балансировать.
Сенсорная информация (например, вкус, свет, запах, давление, вибрация, тепло и боль) воспринимается через сенсорные рецепторы тела.Эти сенсорные рецепторы включают глаза, уши, рот, нос, руки и ноги (и кожу в целом). Рецепторы палочек и колбочек в сетчатке глаза воспринимают свет; реснички в ухе воспринимают звук; химические рецепторы в носовой полости и во рту воспринимают запах и вкус; и мышечные веретена , , а также рецепторы давления, вибрации, тепла и боли в коже воспринимают многочисленные ощущения прикосновения.
Специализированные клетки сенсорных рецепторов преобразуют поступающую энергию (например,г., свет) в нервные импульсы. Эти нервные импульсы поступают в кору головного мозга, которая состоит из слоев нейронов с множеством входных сигналов. Эти слои нейронов действуют как мини-микропроцессоры, и их работа состоит в том, чтобы организовывать ощущения и интерпретировать их в процессе восприятия.
ФМРТ и органы чувств : Эта диаграмма фМРТ показывает некоторую нервную активацию, которая происходит во время ощущения. Например, при визуальной стимуляции активируется затылочная доля.
Мотор Гомункул
«Моторный гомункул» — это теоретическое физическое представление человеческого тела внутри мозга. Это неврологическая «карта» анатомических отделов тела. Внутри первичной моторной коры моторные нейроны расположены упорядоченным образом — параллельно структуре физического тела, но в инвертированном виде. Пальцы ног представлены в верхней части полушария головного мозга, в то время как рот представлен в нижней части полушария, ближе к части мозга, известной как боковая борозда.Эти представления лежат вдоль складки коры головного мозга, называемой центральной бороздой. Гомункул разделен пополам по всему мозгу, с моторным представлением каждой стороны тела, представленной на противоположной стороне мозга.
Количество коры головного мозга, предназначенное для любой данной области тела, пропорционально количеству нервов в этой области, а не ее физическому размеру. Области тела с более или менее сложными сенсорными или моторными связями у гомункула представлены более крупными.Те, у которых меньше или менее сложные связи, представлены как меньшие. В результате получилось изображение искаженного человеческого тела с непропорционально большими руками, губами и лицом (потому что в этих областях есть огромное количество нервных окончаний).
Моторный гомункул : Моторный гомункул — это теоретическая визуализация участков коры головного мозга, которые соответствуют моторным и сенсорным функциям в теле.
Абсолютные пороги чувствительности
Абсолютный порог — это самая низкая интенсивность, при которой может быть обнаружен стимул.
Цели обучения
Объясните, что такое сенсорный абсолютный порог и как на него можно повлиять
Основные выводы
Ключевые моменты
- Абсолютный порог — это наименьший обнаруживаемый уровень любого сенсорного стимула.
- Существует несколько факторов, которые могут влиять на уровень абсолютного порога, включая когнитивные процессы, адаптацию к стимулу, а также мотивацию и ожидания человека.
- Сенсорная адаптация происходит, когда наши органы чувств больше не воспринимают постоянный стимул.
- Мотивации и ожидания человека могут повлиять на то, будет ли стимул обнаружен на абсолютном пороге, и какой уровень дополнительного стимула может потребоваться человеку для обнаружения стимула.
Ключевые термины
- сенсорный порог : точка, в которой стимул вызывает ощущение у человека; ниже сенсорного порога ощущений не будет.
- стимул : Все, что эффективно воздействует на любой из сенсорных аппаратов живого организма, включая физические явления, как внутренние, так и внешние по отношению к телу.
Абсолютные пороги
Порог — это минимальный уровень, при котором может произойти данное событие. В нейробиологии и психофизике существует несколько типов сенсорного порога. Порог распознавания — это уровень, на котором стимул можно не только обнаружить, но и распознать; дифференциальный порог — это уровень, на котором может быть воспринята разница в обнаруженном стимуле; конечный порог — это уровень, выше которого стимул больше не обнаруживается. Однако, возможно, наиболее важным сенсорным порогом является абсолютный порог, который представляет собой наименьший обнаруживаемый уровень стимула.
Абсолютный порог определяется как самая низкая интенсивность, при которой может быть обнаружен стимул. (Недавно теория обнаружения сигнала предложила более детальное определение абсолютного порога: самая низкая интенсивность, при которой стимул будет определяться в определенном процентном соотношении времени, часто в 50%.) Классическим примером абсолютного порога является тест запаха в который аромат испускается в окружающую среду. Абсолютным порогом в этом сценарии будет наименьшее количество аромата, необходимое для того, чтобы субъект обнаружил запах.
Обоняние — не единственное чувство с абсолютными порогами. Представьте, что вы находитесь в комнате, и кто-то позади вас медленно увеличивает громкость радио; абсолютный порог — это самая тихая громкость, при которой вы заметите, что они делают. Звуковые пороги могут быть больше, чем громкость; они также могут относиться к частоте. Например, люди не слышат собачий свист. Это потому, что собачьи свистки имеют частоту, превышающую абсолютный порог частоты для человеческого слуха.
Точно так же минимальное количество света, необходимое для того, чтобы что-то увидеть в темноте, является абсолютным порогом зрения. У каждого чувства есть абсолютный порог.
Свет в конце туннеля: абсолютный порог зрения : В темном пространстве спасительная грация человека может быть минимальным количеством света, необходимым для стимуляции глаза в темной среде и предупреждения мозга о том, что он видит свет.
Влияние на абсолютный порог
Существует несколько факторов, которые могут влиять на уровень абсолютного порога, включая адаптацию к стимулу и индивидуальные мотивации и ожидания.
Сенсорная адаптация
Сенсорная адаптация происходит, когда наши органы чувств больше не воспринимают стимул из-за постоянного контакта с ним нашего сенсорного рецептора. Если вы когда-либо входили в комнату с ужасным запахом, но через несколько минут понимали, что уже почти не замечаете этого, значит, вы испытали сенсорную адаптацию. Подобно порогам, адаптация может происходить в любом смысле, будь то забывание о том, что радио включено, пока вы работаете, или не замечая, что вода в бассейне холодная после того, как вы некоторое время плавали.
Когнитивные процессы
Кроме того, мотивация и ожидания человека также могут влиять на то, будет ли стимул обнаружен на абсолютном пороге. Например, когда вы находитесь в переполненной комнате, где происходит много разговоров, вы склонны сосредотачивать свое внимание на человеке, с которым разговариваете. Поскольку вы сосредоточены на одном стимуле, абсолютный порог (в данном случае минимальная громкость, при которой вы можете слышать) для этого стимула ниже, чем это было бы в противном случае.
Ожидания также могут повлиять на абсолютный порог. Если вы находитесь в темном коридоре в поисках крошечного сияния ночника, ваше ожидание увидеть его снижает абсолютный порог, при котором вы действительно сможете его увидеть.
Пороговые значения сенсорной разницы
Минимальная величина изменения сенсорной стимуляции, необходимая для распознавания того, что изменение произошло, называется просто заметной разницей.
Цели обучения
Определите едва заметную разницу (JND) в терминах сенсорных стимулов
Основные выводы
Ключевые моменты
- Незаметная разница (JND) — это наименьшее обнаруживаемое различие между исходным и вторичным уровнями сенсорного стимула.Это разница в уровне стимула, необходимого человеку, чтобы признать, что изменение произошло.
- Абсолютный порог — это самый низкий уровень, на котором может быть обнаружен стимул. Это точка, в которой наконец-то ощущается запах или наконец слышен звук.
- Порог распознавания — это уровень, на котором стимул может быть не только обнаружен, но и распознан. Это момент, когда мы распознаем определенный запах как дым или звук как сигнал тревоги.
- Конечный порог — это уровень, выше которого стимул больше не обнаруживается.Это точка, в которой сенсорный стимул настолько силен, что сенсорные рецепторы больше не обнаруживают стимул.
Ключевые термины
- сенсорный рецептор : сенсорное нервное окончание, распознающее раздражитель во внутренней или внешней среде организма.
- абсолютный порог : самый низкий уровень, при котором стимул может быть обнаружен в 50% случаев.
Просто заметная разница
Незаметная разница (JND), также известная как разница или дифференциальный порог, представляет собой наименьшее обнаруживаемое различие между начальным и вторичным уровнями сенсорного стимула.Другими словами, это разница в уровне стимула, необходимого человеку, чтобы признать, что изменение произошло.
Увеличение громкости : Порог разницы — это величина изменения стимула, необходимая для распознавания того, что изменение произошло. Если кто-то изменяет громкость динамика, порог различия — это величина, которую необходимо изменить, чтобы слушатели заметили разницу.
Влияние на заметную разницу
JND обычно представляет собой фиксированную пропорцию эталонного сенсорного уровня.Например, подумайте о том, чтобы держать гирю в пять фунтов (контрольный уровень), а затем добавить гирю в один фунт. Это увеличение веса значительно по сравнению с контрольным уровнем (увеличение веса на 20%). Однако, если вы держите гирю в пятьдесят фунтов (новый контрольный уровень), вы вряд ли заметите разницу при добавлении одного фунта. Это связано с тем, что разница в количестве дополнительного веса от контрольного уровня не намного больше (увеличение веса на 2%), чем контрольный уровень.
Абсолютный порог — это минимальная громкость радио, которая нам понадобится для того, чтобы заметить, что оно вообще было включено. Однако определение только что заметной разницы, количество изменений, необходимых для того, чтобы заметить, что радио стало громче, зависит от того, насколько изменилась громкость по сравнению с тем, где она началась. Можно лишь немного увеличить громкость, так что разница в громкости незаметна. Это похоже на добавление всего одного фунта веса, когда вы держите 50 фунтов.
Сенсорная адаптация
Сенсорная адаптация — это снижение отзывчивости сенсорной системы, которая сталкивается с постоянным раздражителем.
Цели обучения
Различают сенсорную адаптацию и сенсибилизацию
Основные выводы
Ключевые моменты
- Сенсорная адаптация включает постепенное снижение сенсорной реакции, вызванное повторным применением определенного стимула с течением времени.
- Тельца Мейснера являются сенсорным триггером физических ощущений на коже, особенно на участках кожи, чувствительных к свету и прикосновениям.
- Примеры сенсорной адаптации: вы больше не чувствуете свою одежду после того, как оделись, больше не слышите звуки проезжающих машин и пешеходную болтовню за пределами городской квартиры после длительного воздействия этих звуков или забываете, что радио или телевизор включены. пока вы сосредоточены на повседневных делах.
Ключевые термины
- тельце : мельчайшая частица; атом; молекула.
Сенсорная адаптация, также называемая нейронной адаптацией, — это изменение реакции сенсорной системы, которая сталкивается с постоянным стимулом.Это изменение может быть положительным или отрицательным и не обязательно приводит к полному игнорированию стимула.
Одним из примеров сенсорной адаптации является длительное прикосновение. Когда вы кладете руки на стол или надеваете одежду на тело, сначала сенсорные рецепторы распознают, что они активируются, и вы чувствуете ощущение прикосновения к объекту. Однако после продолжительного воздействия сенсорные рецепторы больше не будут активироваться так сильно, и вы больше не будете осознавать, что касаетесь чего-либо.
Это следует модели сенсорной адаптации, представленной Георгом Мейснером, которая известна как «тельца Мейснера». Тельца Мейснера являются сенсорным триггером физических ощущений на коже, особенно на участках кожи, чувствительных к свету и прикосновениям. Эти тельца быстро изменяются и адаптируются при добавлении стимула. Затем они быстро снижают активность и со временем перестают реагировать на раздражитель. Когда раздражитель удаляется, тельца вновь обретают чувствительность.Например, постоянное прикосновение одежды к нашей коже приводит к нашей сенсорной адаптации к ощущениям от ношения одежды. Обратите внимание на то, что, когда вы надеваете предмет одежды, по прошествии короткого периода времени вы его больше не чувствуете; однако вы продолжаете испытывать через него другие ощущения. Это потому, что дополнительные раздражители являются новыми, и организм еще не адаптировался к ним.
Напротив, сенсибилизация — это увеличение на поведенческих реакций после повторного применения определенного стимула.В отличие от сенсорной адаптации, при которой требуется большое количество стимула, чтобы вызвать какие-либо дальнейшие ответные эффекты, при сенсибилизации требуется все меньше и меньше стимуляции, чтобы вызвать большой ответ. Например, если животное одновременно слышит громкий шум и испытывает боль, оно испугается сильнее в следующий раз, когда услышит громкий шум, даже если боли нет.
В жизни есть множество стимулов, которые мы испытываем каждый день и постепенно игнорируем или забываем, включая звуки, образы и запахи.Считается, что сенсорная адаптация и сенсибилизация являются неотъемлемым компонентом обучения и личности человека.
Прикосновение и боль | Введение в психологию
Потенциалы действия в рецепторных клетках перемещаются как нервные импульсы с разной скоростью
Когда вы наступаете на булавку, это активирует множество механорецепторов, многие из которых являются ноцицепторами. Вы могли заметить, что ощущение со временем меняется. Сначала вы чувствуете резкий удар, который заставляет вас убрать ногу, и только потом вы чувствуете волну еще более ноющей боли.Об остром ударе сигнализирует быстро проводящие А-волокна, которые проецируются в соматосенсорную кору. Эта часть коры соматотопически организована, то есть сенсорные сигналы представлены в зависимости от того, откуда они берутся в теле (см. Иллюстрацию гомункула, рис. 2). Неприятная боль, которую вы чувствуете после укола острым булавкой, — это отдельный одновременный сигнал, посылаемый от ноцицепторов в вашей стопе через тонкие C-боли или Aδ-волокна в кору островка и другие области мозга, участвующие в обработке эмоций и интероцепции (см. Рис. 3а для схематического изображения этого пути).Другими словами, опыт наступления на булавку складывается из двух отдельных сигналов: один различающий сигнал, позволяющий нам локализовать сенсорный стимул и различать, тупой это или острый удар; и один аффективный сигнал, который сообщает нам, что наступать на булавку — плохо. Обычно боль разделяют на сенсорно-дискриминационный и аффективно-мотивационный аспекты (Auvray, Myin, & Spence, 2010). Это различие соответствует, по крайней мере частично, тому, как эта информация передается от периферии к центральной нервной системе и как она обрабатывается в головном мозге (Price, 2000).
Рисунок 2 . Гомункул: Гомункул означает «маленький человек», и здесь вы видите масштабную модель человеческого тела, искаженную, чтобы отразить относительное пространство, которое части тела занимают в соматосенсорной коре. Как видите, губы, руки, ступни и гениталии посылают в мозг больше соматосенсорных проекций, чем любые другие части тела. Рисунок 2b. Кортикальное картирование сенсорного гомункула: части тела представлены в определенных местах соматосенсорной коры. Представления отображаются соматотопически: стопы расположены медиально, а плечи и руки латеральнее межполушарной щели.Лицевые структуры представлены в другом месте, нежели на коже черепа и головы; лицо ориентировано «вверх ногами», лоб обращен к плечам.
Боль необходима для выживания, но наш мозг может ее остановить, если потребуется
В апреле 2003 года альпинист Арон Ралстон оказался на дне каньона Блю Джон в штате Юта, вынужденный сделать ужасающий выбор: встретить медленную, но верную смерть — или ампутировать правую руку. Пятью днями ранее он упал в каньон — с тех пор его правая рука застряла между валуном весом 800 фунтов и крутой стеной из песчаника.Ослабев от нехватки еды и воды и близок к тому, чтобы сдаться, ему пришло в голову, как прозрение, что, если он сломает две кости в предплечье, ему удастся отрезать остальные своим карманным ножом. Мысль о том, чтобы освободиться и выжить, привела его в такое возбуждение, что он провел следующие 40 минут полностью поглощенный задачей: сначала сломал кости, используя свое тело в качестве рычага, затем сунул пальцы в руку, сжимая пучки мышечных волокон и разрывая их. одну за другой, прежде чем перерезать синие артерии и бледные, похожие на лапшу нервы.Боль была неважной. Только прорезание толстого белого главного нерва заставило его остановиться на минуту — поток боли, как он описывает, был как будто его всю руку толкали «в котел с магмой». Наконец, освободившись, он спустился со скалы и прошел еще 7 миль, пока его не спасли туристы (Ralston, 2010).
Как возможно сделать что-то настолько мучительно болезненное для себя, как это сделал Арон Ральстон, и после этого все еще умудряться ходить, говорить и мыслить рационально? Ответ кроется в мозге, где интерпретируются сигналы тела.Когда мы воспринимаем соматосенсорные и ноцицептивные сигналы от тела, этот опыт очень субъективен и податлив в зависимости от мотивации, внимания, эмоций и контекста.
Рисунок 3 . Пути обработки боли. Слева — восходящие болевые пути: о травме одновременно сигнализируют быстро проводящие Aα или Aβ-волокна и медленно проводящие C-боли или Aδ-волокна. Быстрые А-волокна сигнализируют о давлении, растяжении и других движениях ткани в соматосенсорную кору через ядра спинного столба.С-боль и Aδ-волокна отправляют информацию о боли от ноцицепторов в ткани или коже и передают эти сигналы нейронам второго порядка в спинном роге спинного мозга. Затем нейроны второго порядка переходят на противоположную сторону, где они образуют восходящий спиноталамический тракт. Этот тракт передает сигналы ядрам в продолговатом и среднем мозге на пути к таламусу (Т). Таламус передает информацию соматосенсорной и островной коре, а также кортикальным областям, опосредующим различные аспекты переживания боли, такие как аффективные реакции в поясной коре головного мозга.Справа — нисходящие пути модуляции боли: информация из окружающей среды и определенные мотивационные состояния могут активировать этот путь сверху вниз. Несколько областей лимбического отдела переднего мозга, включая переднюю поясную извилину и островную кору, ядра миндалины и гипоталамуса (H), проецируются в периакведуктальный серый (PAG) среднего мозга, который затем модулирует восходящую передачу боли от афферентной системы боли косвенно через ростральную вентромедиальный мозг (RVM) в стволе мозга. Эта модулирующая система производит анальгезию за счет высвобождения эндогенных опиоидов и использует ON- и OFF-клетки для осуществления либо ингибирующего (зеленый), либо вспомогательного (красный) контроля ноцицептивных сигналов в спинном роге спинного мозга.
Модель мотивации – решения и нисходящая модуляция боли
Согласно модели мотивации и принятия решений , мозг автоматически и непрерывно оценивает плюсы и минусы любой ситуации, взвешивая надвигающиеся угрозы и доступные награды (Fields, 2004, 2006). Все, что важнее для выживания, чем предотвращение боли, активирует нисходящую модуляторную систему боли в головном мозге — нисходящую систему, включающую несколько частей мозга и ствол мозга, которая подавляет ноцицептивную передачу сигналов, чтобы можно было уделить внимание более важным действиям.
В крайнем случае Арона, его действия, вероятно, были основаны на таком бессознательном процессе принятия решений — принимая во внимание его гомеостатическое состояние (его голод, жажда, воспаление и разрушение его раздавленной руки, медленно влияющие на остальную часть его тела), сенсорный ввод доступный (сладкий запах его растворяющейся кожи, тишина вокруг него, указывающая на его одиночество) и его знания об угрозах, стоящих перед ним (смерть или мучительная боль, которая не убьет его), по сравнению с потенциальными наградами (выживание, встречи с семьей очередной раз).История Арона иллюстрирует эволюционное преимущество способности отключать боль: нисходящая система модуляции боли позволяет нам выполнять действия, которые могут спасти жизнь.
Однако, когда кто-то достиг безопасности или получил награду, исцеление становится более важным. Та же самая нисходящая система может затем «вызвать» болевые ощущения от тела, чтобы способствовать исцелению и мотивировать нас избегать потенциально болезненных действий. Чтобы облегчить или подавить ноцицептивные сигналы от тела, нисходящая система модуляции боли использует набор ВКЛЮЧЕННЫХ или ВЫКЛЮЧЕННЫХ клеток в стволе мозга, которые регулируют, какая часть ноцицептивного сигнала достигает мозга.Нисходящая система зависит от передачи сигналов опиоидов, и анальгетики, такие как морфин, снимают боль через этот контур (Petrovic, Kalso, Petersson, & Ingvar, 2002).
Обезболивающая сила вознаграждения
Размышления о хороших вещах, таких как его близкие и предстоящая жизнь, были, вероятно, решающими для выживания Арона. Обещания награды может быть достаточно, чтобы облегчить боль. Ожидание облегчения боли (уменьшение боли часто является наилучшим возможным результатом, если вы испытываете боль, т. Е. Это награда) от лечения, способствует эффекту плацебо, когда облегчение боли происходит, по крайней мере, частично за счет нисходящей модуляции вашего мозга. цепи, и такое облегчение зависит от собственной опиоидной системы мозга (Eippert et al., 2009; Eippert, Finsterbusch, Bingel, & Buchel, 2009; Левин, Гордон и Филдс, 1978). Вкусная еда, прослушивание хорошей музыки или приятные прикосновения к коже также уменьшают боль как у животных, так и у людей, предположительно, благодаря тому же механизму в мозге (Leknes & Tracey, 2008).
В ставшем теперь уже классическом эксперименте Дам и Герц (1984) либо кормили крыс нормальной крысиной едой, либо позволяли им лакомиться очень полезными конфетами в шоколаде (крысы любят сладости), стоя на металлической тарелке, пока они не научились точно, чего ожидать при размещении. там.Когда тарелку нагревали до вредного / болезненного уровня, крысы, ожидавшие леденцов, выдерживали температуру в два раза дольше, чем крысы, ожидающие нормальной еды. Более того, этот эффект полностью исчез, когда опиоидная (эндорфиновая) система крыс была заблокирована лекарством, что указывает на то, что обезболивающий эффект ожидания вознаграждения был вызван высвобождением эндорфина.
Для Арона-альпиниста стресс от осознания надвигающейся смерти и ожидание награды за выживание, вероятно, наводнили его мозг эндорфинами, что способствовало волне возбуждения и эйфории, которую он испытал, выполняя ампутацию ». как пятилетний ребенок, получивший рождественские подарки »(Ralston, 2010).Это изменило его переживание боли от сильного повреждения тканей, которое он причинял, и позволило ему сосредоточиться на освобождении. Оказывается, наш мозг может модулировать восприятие того, насколько неприятна боль, сохраняя при этом способность ощущать интенсивность ощущения (Rainville, Duncan, Price, Carrier, & Bushnell, 1997; Rainville, Feine, Bushnell, & Дункан, 1992). Социальные награды, например, держание за руку парня или подруги, обладают обезболивающим эффектом. Даже просмотр его фотографии может иметь аналогичные эффекты — на самом деле, изображение человека, с которым мы чувствуем близость, не только снижает субъективную оценку боли, но также снижает активность связанных с болью областей мозга (Eisenberger et al., 2011). Таким образом, наиболее распространенные действия, которые нужно делать, когда вы хотите помочь кому-то пережить болезненный опыт, — присутствовать и держать человека за руку, — таким образом, они имеют ощутимый положительный эффект.
Сила разума
Контекст боли и прикосновения имеет большое влияние на то, как мы их интерпретируем. Только представьте, как иначе было бы Арону, если бы кто-нибудь ампутировал ему руку против его воли и без видимой причины. Продолжительную боль от травм будет легче перенести, если инцидент, вызвавший их, дает положительный контекст — например, военная рана, свидетельствующая о храбрости и целеустремленности солдата, — или фантомная боль от руки, отрезанной для продолжения жизни.
Относительное значение боли проиллюстрировано недавним экспериментом, в котором одно и то же умеренно болезненное тепло применялось к участникам в двух разных контекстах — в одном контрольном контексте, где альтернативой было безболезненное тепло; и другой, где альтернативой была очень болезненная жара. В контексте контроля, где умеренная жара была наименее предпочтительным исходом, она (неудивительно) была оценена как болезненная. В другом контексте это был наилучший возможный результат, и здесь точно такая же умеренно болезненная жара была фактически оценена как приятных — потому что это означало, что очень болезненной жары удалось избежать.Это несколько неожиданное изменение восприятия — когда боль становится приятной, потому что она представляет собой облегчение от чего-то худшего — подчеркивает важность того значения, которое люди приписывают своей боли, что может иметь решающее влияние на лечение боли (Leknes et al., 2013). В случае прикосновения, знание того, кто или что гладит вашу кожу, может иметь решающее значение — попробуйте подумать о слизнях в следующий раз, когда кто-то поглаживает вашу кожу, если вы хотите проиллюстрировать этот момент.
У боли и удовольствия есть общие модуляторные системы — еще один общий атрибут состоит в том, что нам не обязательно получать ее сами, чтобы испытать ее.Что вы почувствовали, когда прочитали об Ароне, разрезавшем свои собственные ткани, или о том, что «Томас» бессознательно разрушил свои собственные кости? Вы съежились? Вполне вероятно, что некоторые из областей вашего мозга, обрабатывающие аффективные аспекты боли, были активны, хотя ноцицепторы в вашей коже и глубоких тканях не срабатывали. Боль может ощущаться косвенно, как и зуд, приятные прикосновения и другие ощущения. Таня Сингер и ее коллеги обнаружили в исследовании фМРТ, что некоторые из тех областей мозга, которые были активны, когда участники чувствовали боль на собственной коже (передняя поясная извилина и островок), также были активны, когда им давали сигнал о том, что любимый человек чувствует боль.Те, кто был наиболее «чутким», также показали самые сильные реакции мозга (Singer et al., 2004). Аналогичный эффект был обнаружен для приятного прикосновения: задний островок у участников, просматривающих видео, где нежно поглаживают чужую руку, демонстрирует такую же активацию, как если бы они сами получали прикосновение (Morrison, Bjornsdotter, & Olausson, 2011).
рецепторов и личность | Психология сегодня
Большинство черт личности передаются по наследству.Тем не менее, есть большие сомнения в том, можно ли генетически детерминировать сложное человеческое поведение. Рецепторы нейротрансмиттеров являются продуктами экспрессии генов. Возможно, количество специфических рецепторов в мозге связывает генотип и личность.
История Полевки
Некоторые из наиболее убедительных доказательств роли нервных рецепторов в личности получены не от людей, а от полевок. Луговые полевки живут парами с самцами, ухаживающими за детенышами.Среди горных полевок самцы и самки ненадолго встречаются для спаривания, а затем расходятся.
«Семейно-ценностное» поведение самцов степной полевки связано с большим запасом рецепторов окситоцина в мозгу (1, это «гормон объятий», который обеспечивает тесные социальные контакты у всех млекопитающих, включая человека).
Когда количество рецепторов окситоцина было экспериментально увеличено у горных полевок, они стали напоминать степных полевок, впервые установив парные связи (2).(Эта манипуляция заключалась в прикреплении рецепторов к вирусам простуды, которые служили переносчиками их в мозг).
Успех эксперимента показал, что на поведенческие склонности сильно влияет популяция нейрорецепторов в головном мозге. То же самое, вероятно, верно и в отношении людей, и теоретики личности давно отметили связь между типами личности и системами нейротрансмиттеров.
Личность и нейротрансмиттеры
Различные теоретики личности коррелируют личностные черты с активностью в различных областях мозга. Например, боязнь связана с миндалевидным телом, а стремление к удовольствию связано с проходящим через прилежащее ядро.
Такие пути используют определенные нейротрансмиттеры, а система удовольствия использует дофамин, а боязнь основана на путях серотонина.
Активность нейротрансмиттерных систем можно использовать для объяснения основных параметров личности. Например, экстраверсия связана с повышенной активностью дофамина, которая способствует исследовательскому поведению.
Активность серотонина связана с сознательностью, покладистостью и уровнем тревожности. Таким образом, можно связать некоторые из основных параметров личности с уровнем активности различных нейротрансмиттерных систем в головном мозге.Если это рассуждение верно, то как это помогает объяснить, как наследуются типы личности?
Личность и гены дофаминовых рецепторов
Когда один нейрон стимулирует другой, нейромедиатор высвобождается в синапсе и пересекает синаптическую щель, прежде чем соединиться с рецептором и изменить электрическую активность нейрона-реципиента.
Как сложные белковые молекулы, рецепторы являются продуктами экспрессии генов и, следовательно, обеспечивают вероятные механизмы, посредством которых типы личности могут передаваться по наследству.Обоснование этого состоит в том, что, например, если присутствует больше дофаминовых рецепторов, человек будет лучше искать ощущения.
Мы знаем, что это вряд ли будут простые отношения. В конце концов, количество доступного нейромедиатора также важно, как и присутствие любого разрушающего фермента или других химических веществ, которые взаимодействуют с молекулами нейротрансмиттера.
Возможно, из-за этих осложнений, известие о том, что гены дофамина идентифицированы как коррелят поиска сенсаций, было принято с некоторой помпой.Признак поиска ощущений представляет некоторый интерес для врачей, поскольку он позволяет прогнозировать более раннее употребление наркотиков, вызывающих привыкание, и другие виды рискованного поведения (3).
Гены дофамина довольно слабо коррелируют с поиском ощущений — на их долю приходится менее 4 процентов вариабельности признака в одном из наиболее проведенных исследований, в котором изучались четыре гена дофамина (имеющие 273 однонуклеотидных полиморфизма или вариантов) у 635 неродственных людей. .
Почему рецепторное воздействие на личность слабое
Наследственность поиска ощущений составляет около 58 процентов, и это устанавливает предел максимального количества индивидуальных различий, которые могут быть объяснены генами дофамина.Исследователи отмечают, что на их исследование приходится 6,6% наследственных различий (93,4% остаются необъясненными).
Важные материалы для чтения
Хотя стремление к ощущениям очень передается по наследству, как и другие черты личности, большинство этих эффектов не связано с генами, кодирующими дофаминовые рецепторы.
Возможно, здесь задействовано много других генов, включая те, которые наиболее активны на ранней стадии развития и влияют на появление дофаминергических путей.Конечно, есть возможные генетические эффекты на другие системы нейротрансмиттеров, которые влияют на поиск ощущений. Затем можно было бы указать количество деградирующих ферментов и нейромодуляторов.
Более того, мы начинаем понимать, что на то, как гены экспрессируются в головном мозге, влияет психологический опыт (4). Таким образом, цикл жестокого воспитания теперь связан со сниженной экспрессией глюкокортикоидных рецепторов и повышенным метилированием гена BDNF (нейротрофический фактор мозга) в лобной коре (что само по себе связано с психопатологией, помимо того, что играет роль в обучении и памяти, 5).
В конце концов, взаимодействие рецептор-нейромедиатор — это всего лишь один шаг в сложной цепи событий, с помощью которой нейроны обрабатывают информацию и запускают поиск ощущений. Мы далеки от понимания генетической основы личности.
Список литературы
1 Инсел Р. и Хулихан Т. (1995). Гендерный механизм парных связей: формирование предпочтений окситоцина и партнера у моногамных полевок. Поведенческая неврология, 109, 782-789.
2 Барбер, Н. (2004). Доброта в жестоком мире: эволюция альтруизма. Амхерст, штат Нью-Йорк: Прометей.
3 Дерринджер Дж., Крюгер М. Р. Ф., Дик Д. М. и др. (2010). Предсказание поиска ощущений от дофаминовых генов. Психологическая наука, 21, 1282-1290.
4 Мур, Д. С. (2015). Развивающийся геном: введение в поведенческую эпигенетику. Нью-Йорк: Издательство Оксфордского университета.
5 Франклин, Т. Б., и Мэнсуи, И. М. (2010). Эпигенетическая наследственность у млекопитающих: доказательства воздействия неблагоприятных экологических явлений.Нейробиология болезней, 39, 61-65.
5.5 Другие чувства — Психология 2e
Цели обучения
К концу этого раздела вы сможете:- Опишите основные функции химических органов чувств
- Объясните основные функции соматосенсорной, ноцицептивной и термоцептивной сенсорных систем
- Опишите основные функции вестибулярной, проприоцептивной и кинестетической сенсорных систем
Зрение и слух на протяжении многих лет привлекали невероятное внимание исследователей.Хотя еще многое предстоит узнать о том, как работают эти сенсорные системы, мы гораздо лучше понимаем их, чем другие наши сенсорные модальности. В этом разделе мы исследуем наши химические чувства (вкус и запах) и чувства нашего тела (прикосновение, температура, боль, равновесие и положение тела).
Химические чувства
Вкус (вкус) и запах (обоняние) называются химическими чувствами, потому что оба имеют сенсорные рецепторы, которые реагируют на молекулы в пище, которую мы едим, или в воздухе, которым мы дышим.Между нашими химическими чувствами существует явное взаимодействие. Например, когда мы описываем вкус данной пищи, мы действительно имеем в виду вкусовые и обонятельные свойства пищи, действующие в сочетании.
Вкус (Gustation)
С начальной школы вы узнали, что есть четыре основных группы вкусов: сладкий, соленый, кислый и горький. Однако исследования показывают, что у нас есть по крайней мере шесть групп вкусов. Умами — наш пятый вкус. Умами — на самом деле японское слово, которое примерно переводится как вкусный, и оно связано со вкусом глутамата натрия (Kinnamon & Vandenbeuch, 2009).Также появляется все больше экспериментальных данных, свидетельствующих о том, что мы обладаем вкусом к жирности данной пищи (Mizushige, Inoue, & Fushiki, 2007).
Молекулы пищи и напитков, которые мы потребляем, растворяются в нашей слюне и взаимодействуют со вкусовыми рецепторами на нашем языке, во рту и горле. Вкусовые рецепторы образованы группами вкусовых рецепторных клеток с волосковыми выступами, которые выступают в центральную пору вкусового рецептора (рис. 5.21). Жизненный цикл вкусовых рецепторов составляет от десяти дней до двух недель, поэтому даже уничтожение некоторых из них сжиганием языка не даст долгосрочного эффекта; они просто отрастают снова.Молекулы вкуса связываются с рецепторами на этом участке и вызывают химические изменения в сенсорной клетке, в результате чего нервные импульсы передаются в мозг через разные нервы, в зависимости от того, где расположен рецептор. Информация о вкусе передается в мозговой слой, таламус и лимбическую систему, а также во вкусовую кору, которая находится под перекрытием лобной и височной долей (Maffei, Haley, & Fontanini, 2012; Roper, 2013).
Рисунок 5.21 (а) Вкусовые рецепторы состоят из ряда отдельных клеток вкусовых рецепторов, которые передают информацию нервам. (b) На этой микрофотографии крупным планом показана поверхность языка. (кредит а: модификация работы Йонаса Теле; кредит б: данные шкалы от Мэтта Рассела)
Запах (обоняние)
Обонятельные рецепторные клетки расположены в слизистой оболочке в верхней части носа. Небольшие волоскоподобные отростки от этих рецепторов служат местами для молекул запаха, растворенных в слизи, для взаимодействия с химическими рецепторами, расположенными на этих отростках (Рисунок 5.22). Как только молекула запаха связывает данный рецептор, химические изменения в клетке приводят к тому, что сигналы посылаются в обонятельную луковицу: луковичную структуру на кончике лобной доли, где начинаются обонятельные нервы. Из обонятельной луковицы информация отправляется в области лимбической системы и в первичную обонятельную кору, которая расположена очень близко от вкусовой коры (Lodovichi & Belluscio, 2012; Spors et al., 2013).
Рис. 5.22. Обонятельные рецепторы — это волоскоподобные части, которые простираются от обонятельной луковицы до слизистой оболочки носовой полости.
Чувствительность обонятельных систем разных видов сильно различается. Мы часто думаем, что у собак обонятельная система намного лучше, чем у нас, и действительно, собаки могут делать некоторые замечательные вещи своим носом. Есть некоторые свидетельства того, что собаки могут «чувствовать запах» опасных падений уровня глюкозы в крови, а также раковых опухолей (Wells, 2010). Необычные обонятельные способности собак могут быть связаны с увеличением количества функциональных генов обонятельных рецепторов (от 800 до 1200) по сравнению с менее чем 400, наблюдаемыми у людей и других приматов (Niimura & Nei, 2007).
Многие виды реагируют на химические сообщения, известные как феромоны, посылаемые другим человеком (Wysocki & Preti, 2004). Феромонное общение часто подразумевает предоставление информации о репродуктивном статусе потенциального партнера. Так, например, когда самка крысы готова к спариванию, она выделяет феромональные сигналы, которые привлекают внимание ближайших самцов крыс. Активация феромонов на самом деле является важным компонентом в выявлении сексуального поведения у самцов крыс (Furlow, 1996, 2012; Purvis & Haynes, 1972; Sachs, 1997).Также было проведено множество исследований (и разногласий) о феромонах у людей (Comfort, 1971; Russell, 1976; Wolfgang-Kimball, 1992; Weller, 1998).
Прикосновение, термоцепция и ноцицепция
Ряд рецепторов распределен по коже, чтобы реагировать на различные раздражители, связанные с прикосновением (рис. 5.23). Эти рецепторы включают тельца Мейснера, тельца Пачини, диски Меркеля и тельца Руффини. Тельца Мейснера реагируют на давление и низкочастотные колебания, а тельца Пачини обнаруживают переходное давление и более высокочастотные колебания.Диски Меркель реагируют на легкое давление, а тельца Руффини обнаруживают растяжение (Abraira & Ginty, 2013).
Рис. 5.23. В коже расположено множество типов сенсорных рецепторов, каждый из которых настроен на определенные стимулы, связанные с прикосновением.
Помимо рецепторов, расположенных в коже, существует также ряд свободных нервных окончаний, которые выполняют сенсорные функции. Эти нервные окончания реагируют на различные типы сенсорных стимулов и служат сенсорными рецепторами как для термоцепции (восприятия температуры), так и для ноцицепции (сигнала, указывающего на потенциальный вред и, возможно, на боль) (Garland, 2012; Petho & Reeh, 2012; Спрей, 1986).Сенсорная информация, полученная от рецепторов и свободных нервных окончаний, проходит вверх по спинному мозгу и передается в области мозгового вещества, таламуса и, в конечном итоге, в соматосенсорную кору, которая расположена в постцентральной извилине теменной доли.
Восприятие боли
Боль — это неприятное переживание, которое включает как физические, так и психологические компоненты. Ощущение боли является довольно адаптивным, потому что оно заставляет нас осознать травму и побуждает нас избавиться от причины этой травмы.Кроме того, боль также снижает вероятность получения дополнительных травм, потому что мы будем осторожнее обращаться с травмированными частями тела.
Вообще говоря, боль можно рассматривать как невропатическую или воспалительную по своей природе. Боль, сигнализирующая о каком-либо типе повреждения тканей, известна как воспалительная боль. В некоторых ситуациях боль возникает в результате повреждения нейронов периферической или центральной нервной системы. В результате болевые сигналы, посылаемые в мозг, преувеличиваются. Этот тип боли известен как невропатическая боль.Различные варианты лечения для облегчения боли варьируются от расслабляющей терапии до использования анальгетиков и глубокой стимуляции мозга. Наиболее эффективный вариант лечения для данного человека будет зависеть от ряда факторов, включая серьезность и постоянство боли, а также любые медицинские / психологические состояния.
Некоторые люди рождаются без способности чувствовать боль. Это очень редкое генетическое заболевание известно как врожденная нечувствительность к боли (или врожденная анальгезия).Люди с врожденной анальгезией могут определять разницу в температуре и давлении, но не могут испытывать боль. В результате они часто получают серьезные травмы. У маленьких детей серьезные травмы рта и языка, потому что они неоднократно кусали себя. Неудивительно, что у людей, страдающих этим заболеванием, продолжительность жизни намного короче из-за травм и вторичных инфекций пораженных участков (Национальная медицинская библиотека США, 2013).
Вестибулярное чувство, проприоцепция и кинестезия
Вестибулярное чувство способствует нашей способности сохранять равновесие и осанку.Как показано на рис. 5.24, основные органы чувств (маточный мешок, мешочек и три полукружных канала) этой системы расположены рядом с улиткой во внутреннем ухе. Вестибулярные органы заполнены жидкостью и имеют волосковые клетки, подобные тем, которые находятся в слуховой системе, которые реагируют на движение головы и силы тяжести. Когда эти волосковые клетки стимулируются, они посылают сигналы в мозг через вестибулярный нерв. Хотя при нормальных обстоятельствах мы можем не осознавать сенсорную информацию нашей вестибулярной системы, ее важность становится очевидной, когда мы испытываем укачивание и / или головокружение, связанные с инфекциями внутреннего уха (Khan & Chang, 2013).
Рис. 5.24. Основные органы чувств вестибулярной системы расположены рядом с улиткой во внутреннем ухе. К ним относятся матка, мешочек и три полукружных канала (задний, верхний и горизонтальный).
В дополнение к поддержанию равновесия вестибулярная система собирает информацию, важную для управления движением и рефлексами, которые перемещают различные части нашего тела, чтобы компенсировать изменения в положении тела. Следовательно, и проприоцепция (восприятие положения тела), и кинестезия (восприятие движения тела в пространстве) взаимодействуют с информацией, поступающей из вестибулярной системы.
Эти сенсорные системы также собирают информацию от рецепторов, которые реагируют на растяжение и напряжение в мышцах, суставах, коже и сухожилиях (Lackner & DiZio, 2005; Proske, 2006; Proske & Gandevia, 2012). Проприоцептивная и кинестетическая информация передается в мозг через позвоночник. Некоторые области коры в дополнение к мозжечку получают информацию от органов чувств проприоцептивной и кинестетической систем.
.