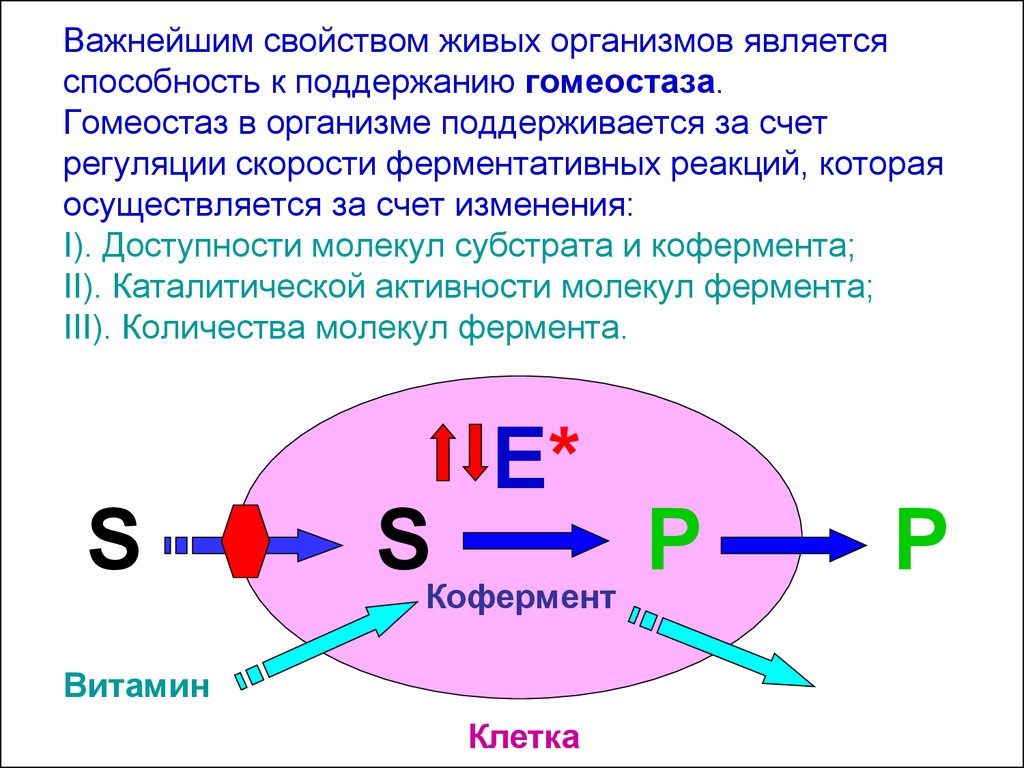
Скрининг системы гемостаза (свертывающей системы крови)
Житель района
Скидки для друзей из социальных сетей!
Скрининг системы гемостаза (то есть, свертывающей системы крови) — пожалуй, один из самых часто назначаемых тестов в современной лабораторной диагностике. И это легко объяснимо. Определение показателей гемостаза является важнейшим этапом обследования при выявлении различных нарушений сердечно-сосудистого, гинекологического, гепатологического, флебологического, эндокринного профиля и т.д. Незаменимо это исследование при мониторинге состояния пациентов в пред- и постоперационном периоде.
Данный анализ обязательно проводят при выявленной склонности человека к образованию тромбов или, напротив, при чрезмерной кровоточивости (гипер- и гипокоагуляция соответственно). Первый вариант отклонения от нормы нередко встречается у пожилых и заядлых курильщиков, у женщин, подолгу принимающих оральные контрацептивы. Также усиленная свертываемость крови может сопутствовать инфекционным заболеваниям, травмам, ожогам. О расположенности организма к чрезмерной кровоточивости могут говорить частые носовые кровотечения и склонность к образованию синяков буквально «на ровном месте».
Также усиленная свертываемость крови может сопутствовать инфекционным заболеваниям, травмам, ожогам. О расположенности организма к чрезмерной кровоточивости могут говорить частые носовые кровотечения и склонность к образованию синяков буквально «на ровном месте».
Система гемостаза выполняет несколько жизненно важных функций. Во-первых, это поддержание в жидком состоянии крови в сосудах. Во-вторых, — приостановка кровотечения (за счет загустевания и образования сгустка-эмбола) в случае травмирования сосудов. Помимо «опций» свертывания и противосвертывания есть третья функция — растворяющая (фибринолиз). Как раз она отвечает за рассасывание кровяного сгустка после восстановления целостности сосуда. Конечно же, любой сбой в этих тонких процессах способен привести к опасным для здоровья и даже жизни последствиям. Скрининг гемостаза позволяет оценить, насколько хорошо справляется система свертывания со всеми своими функциями.
1
Лабораторная диагностика в МедикСити
2
Лабораторная диагностика в МедикСити
3
Лабораторная диагностика в МедикСити
Наиболее назначаемым является стандартный анализ, который включает в себя базовые оценочные тесты свертываемости крови.
Как правило, в базовую гемостазиограмму (или коагулограмму) входят:
- фибриноген — ключевой тест состояния системы свертывания;
- протромбин — по изменениям уровня этого белка в крови судят о состоянии печени и ЖКТ;
- тромбоциты — клетки крови, участвующие в регулировании гемостаза;
- протромбиновый индекс (ПТИ), протромбиновое время (ПТВ), международное нормализованное отношение (МНО) — тесты, отображающие внешний путь свертывания крови;
- активированное частичное тромбопластиновое время (АЧТВ) — тест, отражающий работу внутреннего пути свертывания, время, необходимое для формирования кровяного сгустка;
- тромбиновое время — период, который требуется, чтобы неактивный фибриноген превратился в фибрин.
Однако многие состояния требуют проведения развернутого исследования. По сути, расширенная гемостазиограмма — тот же самый тест, но в дополнение к указанным выше базовым параметрам, в него включены столь важные показатели, как:
- антитромбин III и протеин С — факторы системы противосвертывания, недостаточный их уровень говорит о риске образования тромбов;
- D-димер — элемент системы рассасывания кровяного сгустка, большие его значения могут говорить о патологии почек, сахарном диабете, гестозе;
- волчаночный антикоагулянт — показатель, позволяющий подтвердить или опровергнуть факт наличия у пациента системной красной волчанки или иной аутоиммунной патологии;
- толерантность плазмы к гепарину — параметр, свидетельствующий о свертывающей функции, повышенный уровень может указывать на неблагополучие в печени, сердечную недостаточность, предтромбоз и др;
- растворимые фибрин-мономерные комплексы (РФМК) — продукты растворения кровяного сгустка в процессе фибринолиза, повышение их уровня может указывать на патологию соединительной ткани, сепсис, тромбоэмболию;
- время рекальцификации плазмы — показатель образования сгустка белка фибрина, данный параметр отражает свертывающий процесс в целом;
- активированное время рекальцификации (АВР) — период, необходимый для образования фибрина, укороченное время говорит о склонности организма испытуемого к образованию тромбов, удлиненное — о повышенной кровоточивости.

Система гемостаза (скрининг) в клинике «МедикСити»
В нашей клинике вы можете пройти исследование системы гемостаза, как и любое другое лабораторное исследование, за среднюю по Москве плату, в удобные для вас день и время, без очередей, ненужного волнения и спешки.
В анализ входят:
- Система гемостаза (скрининг):
- АЧТВ,Фибриноген,Антитромбин III,Тромбиновое время,МНО+Протромбин
- Фибриноген (Fibrinogen)
- Протромбин (Prothrombin), Международное нормализованное отношение, МНО (International Normalized Ratio, INR)
- Антитромбин III (Antithrombin III)
- Активированное частичное тромбопластиновое время (Activated Partial Thromboplastin Time)
- Тромбиновое время (Thrombin Time)
- Волчаночный антикоагулянт (Lupus Anticoagulant)
- D-димер (D-dimer)
- Протеин C (Protein C)
- Протеин S свободный (Protein S)
Узнать больше о возможностях лабораторной диагностики вы можете здесь.
Задать вопрос специалисту нашего многоканального контакт-центра вы можете по телефону: +7 (495) 604-12-12.
Многопрофильная клиника «МедикСити» — это профессионализм, опыт, знания и новейшие медицинские разработки! А также — комфортная обстановка и заботливый персонал!
Материал подготовлен при участии специалиста:
Рудько Гали Николаевна
Врач-кардиолог, врач функциональной диагностики
Высшая квалификационная категория, Член Российского кардиологического общества
Система гемостаза — — Статьи
Диагностика и методы исследования
Равновесие между свертывающими и противосвертывающими процессами в крови – необходимое условие существования нашего организма. Нарушение этого равновесия приводит к тяжелым последствиям: кровотечению или тромбообразованию. Поддерживается данный баланс системой гемостаза – одной из важнейших функциональных систем организма, которая решает две «противоположные» задачи:
• поддерживает жидкое состояние крови в обычных условиях;
• останавливает кровотечение при повреждении сосуда.
Свёртывающая система крови (гемостаз) нужна для остановки кровотечения, чтобы избежать значительных кровопотерь при повреждении сосудов. Механизмы гемостаза реализуются при любом повреждении эндотелия сосудистой стенки, вызванном физическими, гемодинамическими, химическими факторами, а также воспалительными процессами, действием иммунных комплексов, нарушением метаболизма (атеросклероз, коллагенозы) и др.
Механизмы гемостаза реализуются при любом повреждении эндотелия сосудистой стенки, вызванном физическими, гемодинамическими, химическими факторами, а также воспалительными процессами, действием иммунных комплексов, нарушением метаболизма (атеросклероз, коллагенозы) и др.
Свертывание крови является жизненно необходимым: мутации в генах основных белков свертывания, как правило, летальны. Система гемостаза удерживает абсолютное первенство среди множества систем нашего организма как главная непосредственная причина летальных исходов: люди болеют разными болезнями, но умирают почти всегда от нарушений системы свертывания крови.
Если причина известна, почему же с ней нельзя бороться? Разумеется, бороться можно и нужно: постоянно создаются новые методы диагностики и терапии нарушений системы свертывания. Но проблема заключается в том, что свертывание крови — крайне сложный и во многом еще загадочный биохимический процесс, который запускается при повреждении кровеносной системы и ведет к превращению жидкой плазмы крови в студенистый сгусток, который как пробка затыкает рану и останавливает кровотечение.
Система гемостаза состоит из десятков белков, которые взаимодействуют в сотнях реакций друг с другом, со стенками сосудов, с клетками крови. Нарушения этой системы крайне опасны и могут привести к кровотечению, тромбозу или другим патологиям, которые совместно отвечают за львиную долю смертности и инвалидности в современном мире. Здесь мы рассмотрим устройство этой системы и расскажем о самых современных методах ее исследования.
I. Система свертываемости крови
По современным представлениям, в остановке кровотечения задействованы:
1) сосудисто-тромбоцитарный гемостаз (первичный), в котором принимают участие стенки сосудов, тромбоциты и, отчасти, эритроциты;
2) плазменный гемостаз (вторичный) — когда в процесс свертывания крови включаются белки плазмы (плазменные факторы свертывания крови).
Такое деление гемостаза достаточно условно, так как в организме эти два звена свертывающей системы крови тесно взаимосвязаны.
1. Первичный гемостаз (сосудисто-тромбоцитарный)
Первичный гемостаз (сосудисто-тромбоцитарный)
Обеспечивает остановку кровотечения из мелких сосудов и сосудов с низким артериальным давлением.
Триггер – повреждение сосудистой стенки и обнажение волокон коллагена — запускает события, которые следуют одно за другим следующим образом.
Реакция кровеносного сосуда
1) Спазм – моментальное рефлекторное сужение сосуда.
Реакция тромбоцитов
2) Адгезия – тромбоциты, благодаря наличию рецепторов к коллагену, прилипают к внутренней стенке сосуда в месте повреждения. Такая стабилизация не дает току крови смывать сгусток тромбоцитов со стенки сосуда.
3) Активация — форма тромбоцитов изменяется, на их поверхности образуются отростки.
4) Агрегация — тромбоциты в большом количестве слипаются, набухают и образуют все более крупный агрегат — рыхлый тромбоцитарный сгусток.
Таким образом, место повреждения сосуда закрывается очень плотной многослойной пробкой (белый тромб), который формируется в течение 3-5 минут. Обычно этого достаточно, чтобы остановить кровотечение у здорового человека при повреждении мелких сосудов.
Обычно этого достаточно, чтобы остановить кровотечение у здорового человека при повреждении мелких сосудов.
Таким образом, первичный гемостаз обусловлен сужением сосудов и их механической закупоркой агрегатами тромбоцитов.
| Единичный тромбоцит в активированном состоянии с отростками | |
| Тромбоцитарный сгусток | |
2. Вторичный гемостаз (плазменный, коагуляция)
Первичный белый тромб — это только временное решение, так как достаточно резкого движения или даже незначительных колебаний артериального давления (например, при физическом напряжении), чтобы сорвать тромбоцитарную пробку с места повреждения сосудистой стенки. Необходим дополнительный механизм, который укрепит скопление тромбоцитов и плотно свяжет его с краями раны. Начинается

При свертывании крови запускается каскад реакций, превращающих растворенный в плазме белок фибриноген в нерастворимый фибрин, который формирует подобие тонкой сетки. Сеть фибрина захватывает находящиеся рядом лейкоциты и эритроциты, формирует красный тромб, уплотняет его и прикрепляет к краям раны. Коагуляция, или свертывание крови, останавливает кровотечение из крупных сосудов и предотвращает его возобновление. В коагуляционном каскаде реакций участвуют особые белки плазмы — факторы свертывания крови, которые обозначаются римскими цифрами в порядке их открытия (например, фактор II, VII и т.д.). Таким образом, в норме скорость свертывания крови зависит от взаимодействия целого ряда различных факторов.
3. Фибринолиз (растворение сгустка крови)
Система фибринолиза восстанавливает проходимость сосудов после ремонта места повреждения сосудистой стенки. Расщепление фибрина происходит под действием специального фермента — плазмина — с образованием продуктов деградации фибрина (ПДФ). Процесс фибринолиза длится от дней до недель, в зависимости от размера и выраженности повреждения сосуда.
Процесс фибринолиза длится от дней до недель, в зависимости от размера и выраженности повреждения сосуда.
II. Алгоритм диагностики нарушений системы гемостаза
Нарушения в системе свертываемости могут приводить к серьезным, с угрозой жизни, кровотечениям и тромбозам.
Которые, в конечном итоге, являются прямой или косвенной причиной наступления более половины всех летальных исходов: например, тромбозы при травме, сепсисе, онкологическом заболевании, хирургическом вмешательстве и др.
Поэтому своевременная точная оценка состояния системы гемостаза – одна из важнейших задач медицины.
Клиническая лабораторная диагностика решает эту задачу с помощью алгоритма:
от оценочных методов (скрининг) к специальным исследованиям (уточнение диагноза).
1. Скрининг
Первоначально выполняются исследования, отражающие состояние целых звеньев системы гемостаза.
Для этого существует стандартный набор тестов, традиционно называемых скрининговыми:
• время кровотечения
• протромбиновое время (ПВ)
• международное нормализованное отношение (МНО)
• активированное частичное тромбопластиновое время (АЧТВ)
• тромбиновое время (ТВ)
• концентрация фибриногена
• растворимые фибрин-мономерные комплексы (РФМК)
• Д-димер
Диагностическая значимость скрининга:
• нормальные результаты – нет значительных изменений в системе гемостаза
• аномальные результаты – показывают направленность нарушений в системе гемостаза
Протромбиновое время (ПВ) — один из основных базовых тестов в повседневной клинической практике; используется для определения времени свертывания и расчета МНО.
| |
Международное нормализованное отношение (МНО)
| |
Активированное частичное тромбопластиновое время (АЧТВ) – представляет собой время, за которое формируется сгусток в образце плазмы крови, после добавления к ней специальных активаторов этого процесса.
| |
| Фибриноген (по Клаусу) — количественный анализ фибриногена — базовый тест оценки системы гемостаза. Фибриноген (I фактор свертывания) в крови находится в растворенном состоянии и под воздействием тромбина превращается в нерастворимый фибрин (полимеризация). Наиболее распространенный метод определения фибриногена — определение по Клаусу. Он основан на измерении времени, необходимого для образования нерастворимого полимера фибрина в разведенной плазме после добавлении большого количества тромбина. Показатель превращения фибриногена в фибрин: чем фибриногена больше, тем интенсивнее свертывание; если его слишком мало, то сгусток (тромб) образуется недостаточного размера и не может остановить кровотечение. | |
Тромбиновое время (ТВ) – используется для определения времени превращения фибриногена в фибрин и оценки антикоагулянтной активности крови. Превращение фибриногена в фибрин происходит в цитратной плазме после добавления в неё тромбина и кальция. При этом скорость образования фибринового сгустка зависит, главным образом, от количества и функциональной полноценности фибриногена и присутствия в крови антикоагулянтов. Превращение фибриногена в фибрин происходит в цитратной плазме после добавления в неё тромбина и кальция. При этом скорость образования фибринового сгустка зависит, главным образом, от количества и функциональной полноценности фибриногена и присутствия в крови антикоагулянтов. | |
 Кровь собирают в пробирку с цитратом натрия, который действует как антикоагулянт: связывает ионы кальция, без которых кровь не свертывается. Избыток кальция возвращает цитратной плазме способность к свёртыванию. Далее к плазме с кальцием добавляется тканевой фактор (III фактор свертывания), и измеряется время образования сгустка.
Кровь собирают в пробирку с цитратом натрия, который действует как антикоагулянт: связывает ионы кальция, без которых кровь не свертывается. Избыток кальция возвращает цитратной плазме способность к свёртыванию. Далее к плазме с кальцием добавляется тканевой фактор (III фактор свертывания), и измеряется время образования сгустка. Таким образом, оценивается степень воздействия факторов свертывания крови на образование тромба.
Таким образом, оценивается степень воздействия факторов свертывания крови на образование тромба.III. Методы исследования системы свертывания
Клоттинговый (англ. «сlot» – сгусток) – в пробе запускается процесс | |
| Хромогенный – в пробу добавляются субстраты, специфичные к определенному белку системы гемостаза. Когда в процессе свертывания нарабатывается искомый белок, он отщепляет от субстрата хромогенную часть, и по интенсивности хромогенного излучения определяют концентрацию и кривую наработки искомого белка.  Доступно только для оптического способа измерения. | |
| Иммунотурбидиметрический – анализируемый белок связывается с взвешенными в пробе частицами, после чего по интенсивности светорассеяния взвеси определяют концентрацию искомого белка. Доступно только для оптического способа измерения. |
Прибор для оценки системы гемостаза — коагулометр
Работа любого коагулометра основывается на способе регистрации времени образования фибринового сгустка — механическом или оптическом.
IV. Способы исследования системы свертывания
1) Механический способ – время образование сгустка определяется по изменению вязкости реакционной смеси:
• в реакционную кювету помещается металлический шарик;
• вокруг кюветы создаётся вращающееся магнитное поле;
• шарик вращается в магнитном поле со строго фиксированной скоростью или совершает колебательные движения с фиксированной амплитудой;
• при формировании сгустка вязкость пробы растет, движение шарика замедляется;
• прибор регистрирует изменение скорости движения шарика;
• остановка шарика приводит к автоматической остановке секундомера коагулометра.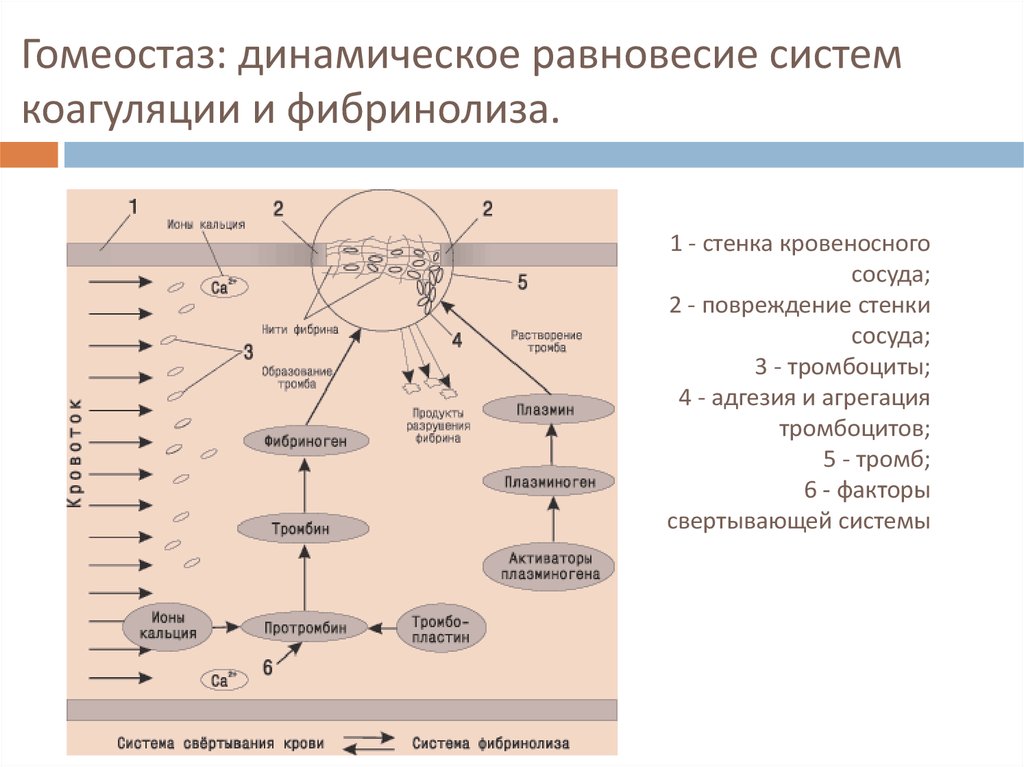
ВАЖНО! Результат механического измерения не зависит от оптической плотности пробы, поэтому в качестве образца можно использовать как цитратную плазму, так и цельную кровь, в том числе, капиллярную.
ВАЖНО! Благодаря тому, что оптическая плотность пробы не влияет на результат механического измерения, можно исследовать «сложные» пробы (иктеричные, липемичные и гемолизные), без их отбраковки и повторных назначений. Доступно только для механического способа измерения.
Механика прощает ошибки преаналитического этапа.
Что такое иктеричность?
Иктеричная проба окрашена в ярко-желтый цвет из-за высокой концентрации билирубина в крови, которая чаще всего обусловлена различными заболеваниями печени, а также приемом некоторых лекарственных препаратов. Высокая концентрация билирубина в пробе может исказить значение лабораторного показателя. Предсказать иктеричность образца, как правило, невозможно. При этом не всегда возможно и скорректировать повышенный уровень билирубина в крови пациента. Чтобы выполнить анализ иктеричной пробы и получить достоверный результат, необходимо использовать соответствующие способы и оборудование, которые позволяют исследовать иктеричную пробу без определения оптической плотности – на механических коагулометрах.
При этом не всегда возможно и скорректировать повышенный уровень билирубина в крови пациента. Чтобы выполнить анализ иктеричной пробы и получить достоверный результат, необходимо использовать соответствующие способы и оборудование, которые позволяют исследовать иктеричную пробу без определения оптической плотности – на механических коагулометрах.
Что такое липемия?
Липемичная проба имеет желтовато-белый цвет из-за высокой концентрации липидов (жиров) в крови. Чаще всего липемия обусловлена приемом жирной пищи незадолго до сдачи крови, а также некоторыми нарушениями обмена веществ, в частности, обмена жиров. Высокая концентрация жиров в крови может исказить значение лабораторного показателя. Как избежать влияния липемии на результат? Если нарушены правила подготовки к сдаче анализов, кровь можно пересдать. Но если липемия обусловлена нарушениями метаболизма, «улучшить» образец невозможно в принципе. Чтобы выполнить анализ такой пробы и получить корректный результат, необходимо использовать соответствующие способы и оборудование (без определения оптической плотности), которые позволяют исследовать мутную пробу.
ВАЖНО! Если аномальная окраска плазмы обусловлена, например, приемом лекарств, то новое взятие образца ситуацию с качеством пробы не улучшит. Для таких пациентов получение результата возможно только механическим способом. Таким образом, только механика даст корректный результат для «сложной» пробы, качество которой улучшить слишком затратно или вообще невозможно.
| ВАЖНО! Особенности механических коагулометров позволяют успешно применять их для оценки гемостаза как в рутинном скрининге, так и в педиатрической практике и при контроле лечения непрямыми антикоагулянтами. |
2) Оптический способ — детекция сгустка по изменению оптической плотности пробы:
• Исходная плазма прозрачна
• Формирование сгустка уменьшает
светопропускание через кювету
• Уменьшение светопропускания фиксируется
оптической системой прибора
ВАЖНО! Результат оптического измерения зависит от оптической плотности пробы, поэтому в качестве образца нельзя использовать цельную кровь, можно использовать только плазму. По этой же причине сложные пробы отбраковываются, а используемые реагенты должны быть прозрачными.
По этой же причине сложные пробы отбраковываются, а используемые реагенты должны быть прозрачными.
Оптический способ имеет лучшую чувствительность при замедленном образовании сгустка, при низком уровне фибриногена, на фоне антикоагулянтной терапии и в случае, если колебания шарика рвут слабые нити фибрина. С другой стороны, при скрининговых исследованиях (область использования полуавтоматических коагулометров) доля пациентов с такими особенностями свертывающей системы очень мала. Они обычно наблюдаются в специализированных клиниках, с другим парком оборудования и набором тестов. Поэтому вышеупомянутая особенность оптического метода не дает какого-либо ключевого преимущества в сравнении с механическим методом в области применения полуавтоматических коагулометров.
| Механика | Оптика |
Физические характеристики плазмы (мутность, желтушность) | Не влияют на результат | Влияют на результат |
Анализ цельной крови | Возможен | Не возможен |
Короткое время сворачивания | Всегда фиксируется | Не всегда фиксируется |
Методы исследования | Клоттинговый | Клоттинговый, хромогенный, турбидиметрический |
Определение сгустка при низком фибриногене | Затруднена | Возможна |
Графическое отображение хода реакции | Нет | Да |
II. Рациональный подход к анализатору гемостаза
Рациональный подход к анализатору гемостаза
Анализаторы для оценки гемостаза можно разделить две группы: полуавтоматические коагулометры и полностью автоматизированные аналитические системы.
Полуавтоматические коагулометры — наиболее востребованные и распространенные в практике лабораторной службы в современной России, особенно в условиях чрезвычайной ситуации с распространением новой коронавирусной инфекции.
Рассмотрим значимые характеристики полуавтоматического коагулометра на примере анализатора TS4000+ производства HTI.
Количество определяемых параметров
Современные полуавтоматические анализаторы гемостаза, в том числе TS4000+, имеют на борту уже предустановленные коагулологические методики (тесты). Как правило, это не менее 12 определяемых параметров гемостаза: основные скрининговые тесты и факторы свертывания.
В меню коагулометра TS4000+ запрограммированы 12 тестов: АЧТВ, ПВ, ТВ, Фибриноген и факторы свертывания II, V, VII, VIII, IX, X, XI, XII.
Метод исследования
Анализаторы гемостаза подразделяют, по способу регистрации момента образования сгустка, на оптические и механические. При этом надо помнить, что нет оптико-механического способа определения сгустка: выпускаются коагулометры оптико-механического типа, в которых технически осуществлена возможность проводить оценку гемостаза механическим или оптическим методом, который выбирает оператор.
Коагулометр TS4000+ использует механический способ регистрации сгустка и не зависит от оптической плотности пробы. Это делает TS4000+ универсальным – он может работать как с цельной кровью, в том числе капиллярной, так и с плазмой, в различных разбавлениях и с применением любых реагентов, даже непрозрачных. Благодаря тому, что оптическая плотность пробы не влияет на измерения TS4000+, на нем возможно тестировать «сложные» пробы (мутные и окрашенные), без их отбраковки и избыточных затрат на повторное выполнение анализа.
Механика TS4000+ прощает ошибки преаналитического этапа.
Напомним, что на рынке РФ представлены также оптические коагулометры, позволяющие проводить исследование гемостаза хромогенными методами. Однако, трудоемкость выполнения, низкая селективность хромогенных субстратов (которые могут реагировать с другими продуктами реакции), высокая стоимость анализа, а также небольшое количество выполняемых соответствующих тестов накладывают ограничения на массовое применение данного метода.
Производительность коагулометра
Анализатор гемостаза выбирают, ориентируясь на прогнозируемое количество коагулологических тестов. Предлагаемые на рынке полуавтоматические коагулометры можно разделить на две группы по производительности: 2-канальные (10–20 проб в день) и 4-канальные (20–40 проб в день). КДЛ неспециализированных лечебных учреждений обычно выполняет стандартную коагулограмму, в среднем, 4 скрининговых теста на пациента.
Соответственно, в таких лабораториях востребованы 4-канальные полуавтоматические коагулометры, такие как TS4000+, которые способны полностью закрыть потребность в рутинных коагулограммах.
Одноканальные коагулометры не следует даже рассматривать, так как их функционал ограничен, а производительность крайне мала.
Объем пробы
Объем пробы для анализа – важный критерий выбора анализатора. Экономичный объем образца позволяет использовать приборы в педиатрии или при скрининговых исследованиях, а также дает возможность выполнять повторные исследования. Малый объем пробы существенно — в 2 раза — снижает потребление реагентов, по сравнению с ручными способами. Например, TS4000+ использует для анализа не более 100 мкл плазмы и 50–200 мкл реагентов.
Стоимость эксплуатации, адаптация к реагентам
При выборе коагулометра большое значение имеет как его первоначальная стоимость и комплектация, так и последующие расходы в течение всего срока эксплуатации (регулярные закупки расходных материалов к нему — кювет, шариков-мешалок и реагентов).
Полуавтоматические коагулометры являются открытыми системами, что позволяет использовать реагенты любого производителя, без специальных адаптаций и аттестаций.
Однако на некоторых коагулометрах производители устанавливают специальные защитные системы, не позволяющие использовать реагенты и расходные материалы сторонних компаний. В таких приборах предусмотрено использование специальных информационных карт, которые поставляются с наборами реагентов. Цель введения таких карт – защитить анализатор от несанкционированного использования не предназначенных для данного прибора реагентов. Например, полуавтоматический коагулометр «КоаТест-4» (НПЦ Астра, Россия) имеет закрытую систему по кюветам и реагентам. На сегодняшний день он адаптирован под реагенты фирмы РЕНАМ. Такая «закрытость» неудобна для лаборатории как с точки зрения привязанности к одному производителю (задержки поставок реагентов), так и с экономической точки зрения (невозможно перейти на менее затратные материалы). В целом, цена закрытого коагулометра ниже, но стоимость «родных» реагентов обычно достаточно высокая.
В целом, цена закрытого коагулометра ниже, но стоимость «родных» реагентов обычно достаточно высокая.
В отличие от «закрытых» полуавтоматических коагулометров, TS4000+ является полностью открытым бюджетным прибором как по первоначальной цене и комплектации, так и по стоимости использования в течение всего срока эксплуатации. Он открыт по реагентам, работает с наборами любых производителей, а также характеризуется низким энергопотреблением.
Удобство эксплуатации
На выбор оборудования также влияет такой фактор как эргономичность. Простота и удобство работы с прибором могут стать решающими при прочих равных характеристиках.
Например, наличие у TS4000+ дозатора шариков, 7 позиций для реагентов с подогревом и без, 16 ячеек для предварительного прогрева образцов, возможность перепрограммирования тестовых протоколов, кнопочная клавиатура, встроенный термопринтер, интерфейс RS232 для передачи информации на отдельный компьютер или во внутреннюю лабораторную сеть – свидетельствуют о том, что данный прибор будет эффективен и удобен в работе.
При выборе коагулометра следует тщательно изучить все его особенности, рассчитать стоимость эксплуатации в соответствии с объемом коагулологических исследований, а также поинтересоваться мнением коллег.
И сделать правильный выбор!
Вернуться
Комплекс исследований системы гемостаза — статьи от специалистов клиники «Мать и дитя»
Система гемостаза — это биологическая система, обеспечивающая, с одной стороны, сохранение жидкого состояния циркулирующей крови (предотвращение образования тромбов), а с другой, — предупреждение и остановку кровотечений. Именно это свойство системы гемостаза обеспечивает нормальное функционирование организма.
Исследование системы гемостаза выполняется в следующих случаях:
- прием гормональных контрацептивов: перед началом использования (для выбора безопасного вида контрацепции) и во время приема КОК (для контроля состояния свертывающей системы) отягощенный акушерский анамнез:
- две и более остановки развития плода
- неудачные попытки ЭКО
- невынашивание беременности
- тяжелый гестоз в анамнезе
- бесплодие
- наличие сердечно-сосудистых заболеваний (особенно после перенесенных инсультов и инфарктов, после операций на сердце)
- наличие у близких родственников в молодом возрасте (младше 40 лет) инфарктов и инсультов
- признаки кровоточивости, как то: носовые кровотечения; наличие множества мелких синячков; длительная кровоточивость при бритье и порезах, из десен, из геморроидальных трещин
- обильные менструальные выделения
- прием непрямых антикоагулянтов (варфарин, дикумарол, фенилин), низкомолекулярных гепаринов (фраксипарин, дельтапарин, клексан, фрагмин, кальципарин) и антиагрегантов (ТромбоАсс, кардиомагнил, аспирин, плавикс)
- возраст пациента старше 50 лет (для определения необходимости применения антиагрегантов)
- оценка функции печени
- обследование перед операцией
- обследование перед программой ЭКО
- обследование перед планируемой беременностью
- обследование во время беременности
- обследование для постановки дифференциального диагноза тромбофилии, геморрагии, гемофилий, АФС
При исследовании системы гемостаза важно оценить работу всех компонентов.

Гемостаз осуществляется тремя взаимодействующими между собой функционально-структурными компонентами:
- Сосудистым (эндотелий сосудистой стенки)
- Клеточным (клетки крови)
- Плазменным (факторы свертывания, антикоагулянты, участники фибринолиза)
В нашей лаборатории мы исследуем каждый этап.
Давайте разберемся — какие исследования характеризуют тот или иной этап гемостаза.
Сосудисто-тромбоцитарный гемостаз:
- Количество тромбоцитов (в составе общего анализа крови или как отдельное исследование), дополнительно рассчитываются тромбоцитарные индексы — средний размер тромбоциты, тромбокрит, анизоцитоз тромбоцитов.
- Определение времени, за которое свертывается кровь при проколе пальца руки.
- Определение функциональной активности тромбоцитов по их способности к агрегации (слипанию). В процессе исследования важно оценить — как тромбоциты агрегируют при воздействии на них различных веществ, так называемых индукторов агрегации (синонимы «активатор», «стимулятор») и без добавления индукторов — спонтанная агрегация.

В случае спонтанной агрегации активизация тромбоцитов происходит за счет турбулентности потока крови (имитация процесса in vivo, т. е. «в организме»).
Также проводится оценка агрегации тромбоцитов под воздействием различных индукторов в разных концентрациях: агрегация с АДФ, коллагеном, ристомицином, арахидоновой кислотой, УИА (универсальным индуктором агрегации).
В нашей лаборатории проводится комплексная агрегатограмма, в которую входит спонтанная агрегация и агрегация с АДФ в трех концентрациях. Такое исследование позволяет более точно оценить функции тромбоцитов. Результат комплексной агрегатограммы выдается с распечаткой графиков и с заключением.
Для исследования агрегации мы используем агрегометр «Биола», который является уникальным прибором, сочетающим общепризнанный метод определения агрегации по Борну (турбидиметрический) и метод ФСП (с оценкой среднего размера агрегатов). Преимущество этого метода в том, что он позволяет оценить агрегацию с низкими физиологическими дозами индуктора, приближая лабораторный анализ, к процессам, протекающим в организме.
Исследование вторичного или плазменного гемостаза:
Свертывание крови протекает по двум путям — внешнему и внутреннему.
- Внешний путь активируется при повреждении кровеносного сосуда извне (например, при порезе) и его можно оценить при проведении протромбинового теста, включающего в себя активность по Квику, протромбиновый индекс и МНО. МНО (международное нормализованное отношение) — это стандартизованный показатель, используемый для контроля применения непрямых антикоагулянтов.
- Внутренний путь активируется при повреждении эндотелия (внутренней выстилки) сосудистой стенки и оценивается тестом АЧТВ (активированное частичное (парциальное) тромбопластиновое время).
- Конечный этап свертывания оценивают два теста — фибриноген и тромбиновое время, позволяя оценить количественные и качественные характеристики этого периода свертывания.
- Активизация свертывающей системы и образование тромбов могут происходить не только при повреждении сосуда, но и при воздействии многих веществ, находящихся в кровотоке.

Насколько активизирована эта система, можно оценить по результатам теста — РФМК (растворимые фибрин-мономерные комплексы — продукты деградации фибриногена). Повышение активности свертывающей системы может привести к повышенному тромбообразованию.
Образовавшиеся тромбы необходимо постепенно лизировать (разрушить), после того как они выполнят свою «функцию» и целостность поврежденного сосуда восстановиться. В дело вступает фибринолитическая система.
Ее «работоспособность» можно оценить при проведении теста: плазминоген, D-димер. Определяя активность плазминогена можно косвенно оценить «работу» плазмина — фермента «разрезающего» фибриновый сгусток. По концентрации D-димера (продукта деградации фибрина) можно сделать вывод, о том насколько активно идёт процесс расщепления образовавшихся сгустков.
- Для того, чтобы наша кровь поддерживалась в жидком состоянии, в плазме крови циркулируют т.н. физиологические (естественные) антикоагулянты. К ним относятся — антитромбин III, протеин С и его кофактор протеин S. Недостаток этих компонентов приводит к тромбозу.
- В некоторых случаях (например, для дифференциальной диагностики гемофилии, А и В, болезни (или синдрома) Виллебранда) необходимо дополнительные исследования для определения активности отдельных факторов: фактора VIII, фактора IX, фактора Виллебранда.
 физиологические (естественные) антикоагулянты. К ним относятся — антитромбин III, протеин С и его кофактор протеин S. Недостаток этих компонентов приводит к тромбозу.
физиологические (естественные) антикоагулянты. К ним относятся — антитромбин III, протеин С и его кофактор протеин S. Недостаток этих компонентов приводит к тромбозу.Говоря о патологии системы гемостаза и клинических состояниях, с ней связанных нельзя обойти тему диагностики антифосфолипидного синдрома (АФС), т.к. это один из видов тромбофилий, связанный с аутоиммунными процессами.
Основными маркерами АФС является наличие антител к кардиолипину и волчаночного антикоагулянта в крови (т.н. люпус-тест). Наличие антител к фосфолипидам в акушерской практике может быть причиной антенатальной гибели плода и невынашивания беременности. Повышенные титры антител были обнаружены при некоторых неврологических заболеваниях.
В последнее время наблюдаются ситуации, при которых оба вышеназванных теста могут оставаться отрицательными при наличии клинических проявлений АФС. Для диагностики таких состояний все чаще применяется определение антител к другим фосфолипидам (фосфатидилинозитолу, фосфатидилсерину), а также антител к бета-2 гликопротеину-1, аннексину V и протромбину.
Причиной повышенной свертываемости крови может быть и гипергомоцистеинемия — повышенное содержание гомоцистеина в крови.
Гомоцистеин является продуктом метаболизма метионина — одной из 8 незаменимых аминокислот организма. В норме он не накапливается. Обладает выраженным токсическим действием на клетку. Циркулируя в крови, повреждает сосуды, тем самым повышая свертываемость крови, способствует образованию микротромбов в сосудах (одна из причин невынашивания беременности, инсультов). Поэтому данное исследование показано лицам с сердечно-сосудистыми заболеваниями, женщинам с привычным невынашиванием, с замершими беременностями в анамнезе и бесплодием.
Все перечисленные выше исследования системы гемостаза можно пройти в Диагностической лаборатории ЗАО «Медицинская компания ИДК».
Подготовка к сдаче анализов: Кровь сдается натощак в утренние часы.
Для сдачи анализа «агрегация тромбоцитов» необходима особенная подготовка:
Кровь сдается натощак, допускается прием некрепкого чая и сухих хлебобулочных изделий.
Перед исследованием нельзя употреблять кофе, алкоголь (в течение суток), есть чеснок, курить, необходимо исключить любую физическую нагрузку. Также нельзя проводить исследование на фоне острого воспаления.
Следующие исследования сдаются в определенные дни по записи в процедурных кабинетах Медицинской компании ИДК на ул. Энтузиастов 29: фактор VIII, фактор IX, фактор Виллебранда, агрегация тромбоцитов, Протеин С и протеин S.
Запись на исследования по тел 8 (846) 215-22-03, (846) 927- 99- 91.
Система гемостаза в норме и при трансплантации печени (обзор) | Решетняк
1. Starzl T.E., Demetris A.J. Liver transplantation: a 31-year perspective. Chicago, London, Boca Raton, Littleton, Mass: Year book medical publishers, Inc.; 1990: 206. ISBN: 0-8151-7931-6
2. Oldenburg J., Marinova M., Müller-Reible C., Watzka M. The vitamin K cycle. Vitam. Horm. 2008; 78: 35-62. DOI: 10.1016/S00836729(07)00003-9. PMID: 18374189
3. Bandyopadhyay P.K., Garrett J.E., Shetty R.P., Keate T., Walker C.S., Olivera B.M. gamma-Glutamyl carboxylation: an extracellular posttranslational modification that antedates the divergence of molluscs, arthropods, and chordates. Proc. Natl. Acad. Sci. USA. 2002; 99 (3): 12641269. DOI: 10.1073/pnas.022637099. PMID: 11818531
4. Bandyopadhyay P.K. Vitamin K-dependent gamma-glutamylcarboxylation: an ancient posttranslational modification. Vitam. Horm. 2008; 78: 157-184. DOI: 10.1016/S0083-6729(07)00008-8. PMID: 18374194
5. Morrissey J. H. Tissue factor: a key molecule in hemostatic and nonhemostatic systems. Int. J. Hematol. 2004; 79 (2): 103-108. DOI: 10.1532/IJH97.03167. PMID: 15005335
H. Tissue factor: a key molecule in hemostatic and nonhemostatic systems. Int. J. Hematol. 2004; 79 (2): 103-108. DOI: 10.1532/IJH97.03167. PMID: 15005335
6. Hanson S.R., Griffin J.H., Harker L.A., Kelly A.B., Esmon C.T., Gruber A. Antithrombotic effects of thrombin-induced activation of endogenous protein C in primates. J. Clin. Invest. 1993; 92 (4): 2003-2012. DOI: 10.1172/JCI116795. PMID: 8408654
7. Shaw A.W., Pureza V.S., Sligar S.G., Morrissey J.H. The local phospholipid environment modulates the activation of blood clotting. J. Biol. Chem. 2007; 282 (9): 6556-6563. DOI: 1074/jbc.M607973200. PMID: 17200119
8. Morrissey J.H., Pureza V., Davis-Harrison R.L., Sligar S.G., Ohkubo Y.Z., Tajkhorshid E. Blood clotting reactions on nanoscale phospholipid bilayers. Thromb. Res. 2008; 122 (Suppl 1): S23-S26. DOI: 10.1016/S00493848(08)70014-8. PMID: 18691494
9. Smith S.A., Morrissey J.H. Polyphosphate: a new player in the field of hemostasis. Curr. Opin. Hematol. 2014; 21 (5): 388-394. DOI: 10.1097/MOH.0000000000000069. PMID: 25010799
DOI: 10.1097/MOH.0000000000000069. PMID: 25010799
10. Morrissey J.H., Davis-Harrison R.L., Tavoosi N., Ke K., Pureza V., Boettcher J.M., Clay M.C., Rienstra C.M., Ohkubo Y.Z., Pogorelov T.V., Tajkhorshid E. Protein-phospholipid interactions in blood clotting. Thromb. Res. 2010; 125 (Suppl 1): S23-S25. DOI: 10.1016/j.thromres.2010.01.027. PMID: 20129649
11. Ke K., Yuan J., Morrissey J.H. Tissue factor residues that putatively interact with membrane phospholipids. PLoS One. 2014; 9 (2): e88675. DOI: 10.1371/journal.pone.0088675. PMID: 24516673
12. Tavoosi N., Smith S.A., Davis-Harrison R.L., Morrissey J.H. Factor VII and protein C arephosphatidic acid-binding proteins. Biochemistry. 2013; 52 (33): 5545-5552. DOI: 10.1021/bi4006368. PMID: 23879866
13. Morawitz P. Beiträge zur Kenntnis der Blutgerinnung. Beitr. chem. Physiol. Pathol. 1904; 5: 133-141.
14. Morawitz P. Beiträge zur Kenntnis der Blutgerinnung. 2. Mitteilung. Arch. Klin. Med. 1904; 79: 215-233.
15. Nahas L., Macfarlane R.G., Denson K.W. A study of the coagulant action of eight snake venoms. Thromb. Diath. Haemorrh. 1964; 12: 355-367. PMID: 14254694
Nahas L., Macfarlane R.G., Denson K.W. A study of the coagulant action of eight snake venoms. Thromb. Diath. Haemorrh. 1964; 12: 355-367. PMID: 14254694
16. Davie E.W., Ratnoff O.D. Waterfall sequence for intrinsic blood clotting. Science. 1964; 145 (3638): 1310-1312. DOI: 10.1126/science.145.3638.1310. PMID: 14173416
17. Broze G.J.Jr. Tissue factor pathway inhibitor and the current concept of blood coagulation. Blood Coagul. Fibrinolysis. 1995; 6 (Suppl 1): S7-S13. DOI: 10.1097/00001721-199506001-00002. PMID: 7647225
18. Hoffman M.A., Monroe D.M.3rd. Сell-based model of hemostasis. Thromb. Haemost. 2001; 85 (6): 958-965. PMID: 11434702
19. Stout R.W., Crawford V. Seasonal variations in fibrinogen concentrations among elderly people. Lancet. 1991; 338 (8758): 9-13. DOI: 10.1016/0140-6736(91)90004-9. PMID: 1676131
20. Stout R.W., Crawford V.L., McDermott M.J., Rocks M.J., Morris T.C. Seasonal changes in haemostatic factors in young and elderly subjects. Age Ageing. 1996; 25 (3): 256-258. DOI: 10.1093/ageing/25.3.256. PMID: 8670564
1996; 25 (3): 256-258. DOI: 10.1093/ageing/25.3.256. PMID: 8670564
21. Hermida R.C., Calvo C., Ayala D.E., López J.E., Fernández J.R., Mojón A., Domínguez M.J., Covelo M. Seasonal variation of fibrinogen in dipper and nondipper hypertensive patients. Circulation. 2003; 108 (9): 1101-1106. DOI: 10.1161/01.CIR.0000085992.67380.7B. PMID: 12912809
22. Hall T.H., Dhir A. Anesthesia for liver transplantation. Semin. Cardiothorac. Vasc. Anesth. 2013; 17 (3): 180-194. DOI: 10.1177/108925321 3481115. PMID: 23482506
23. Palomo Sanchez J.C., Jimenez C., Moreno Gonzalez E., Garcia I., Palma F., Loinaz C., Gonzalez Ghamorro A. Effects of intraoperative blood transfusion on postoperative complications and survival after orthotopic liver transplantation. Hepatogastroenterology. 1998; 45 (22): 1026–1033. PMID: 9756002
24. Cleland S., Corredor C., Ye J.J., Srinivas C., McCluskey S.A. Massive haemorrhage inliver transplantation: сonsequences, prediction and management. World J. Transplant. 2016; 6 (2): 291-305. DOI: 10.5500/wjt.v6.i2.291. PMID: 27358774
2016; 6 (2): 291-305. DOI: 10.5500/wjt.v6.i2.291. PMID: 27358774
25. Kang Y.G., Martin D.J., Marquez J., Lewis J.H., Bontempo F.A., Shaw B.W.Jr., Starzl T.E., Winter P.M. Intraoperative changes in blood coagulation and thrombelastographic monitoring in liver transplantation. Anesth. Analg. 1985; 64 (9): 888–896. DOI: 10.1213/00000539198509000-00008. PMID: 3896028
26. Kriz N., Rinder C.S., Rinder H.M. Physiology of hemostasis: with relevance to current and future laboratory testing. Clin. Lab. Med. 2009; 29 (2): 159-174. DOI: 10.1016/j.cll.2009.06.004. PMID: 19665673
27. Rana A., Petrowsky H., Hong J.C., Agopian V.G., Kaldas F.M., Farmer D., Yersiz H., Hiatt J.R., Busuttil R.W. Blood transfusion requirement during liver transplantation is an important risk factor for mortality. J. Am. Coll. Surg. 2013; 216 (5): 902–907. DOI: 10.1016/j.jamcollsurg.2012.12.047. PMID: 23478547
28. Pereboom I.T., de Boer M.T., Haagsma E.B., Hendriks H.G., Lisman T., Porte R.J. Platelet transfusion during liver transplantation is associated with increased postoperative mortality due to acute lung injury.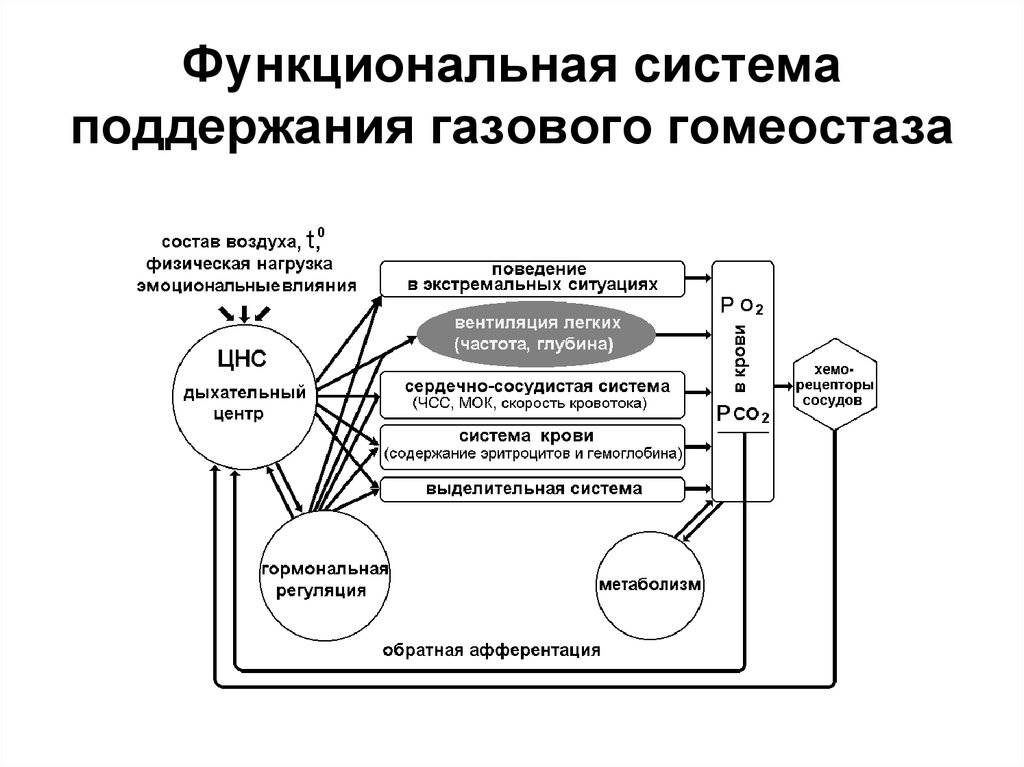 Anesth. Analg. 2009; 108 (4): 1083–1091. DOI: 10.1213/ane.0b013e3181948a59. PMID: 19299765
Anesth. Analg. 2009; 108 (4): 1083–1091. DOI: 10.1213/ane.0b013e3181948a59. PMID: 19299765
29. Steib A., Freys G., Lehmann C., Meyer C., Mahoudeau G. Intraoperative blood losses and transfusion requirements during adult liver transplantation remain difficult to predict. Can. J. Anaesth. 2001; 48 (11): 1075– 1079. DOI: 10.1007/BF03020372. PMID: 11744582
30. Massicotte L., Beaulieu D., Roy J.D., Marleau D., Vandenbroucke F., Dagenais M., Lapointe R., Roy A. MELD score and blood product requirements during liver transplantation: no link. Transplantation. 2009; 87 (11): 16891694. DOI: 10.1097/TP.0b013e3181a5e5f1. PMID: 19502961
31. Mangus R.S., Kinsella S.B., Nobari M.M., Fridell J.A., Vianna R.M., Ward E.S., Nobari R., Tector A.J. Predictors of blood product use in orthotopic liver transplantation using the piggyback hepatectomy technique. Transplant. Proc. 2007; 39 (10): 3207–3213. DOI: 10.1016/j.transproceed.2007.09.029. PMID: 18089355
32. Feng Z.Y., Jin X.D., Chen Y. Z. Predictors of massive blood transfusion in liver transplantation for patients with benign end-stage liver disease. Zhonghua Yi Xue Za Zhi. 2008; 88: 3040–3044. PMID: 19192401
Z. Predictors of massive blood transfusion in liver transplantation for patients with benign end-stage liver disease. Zhonghua Yi Xue Za Zhi. 2008; 88: 3040–3044. PMID: 19192401
33. Li C., Mi K., Wen T.F., Yan L.N., Li B., Wei Y.G., Yang J.Y., Xu M.Q., Wang W.T. Risk factors and outcomes of massive red blood cell transfusion following living donor liver transplantation. J. Dig. Dis. 2012; 13 (3): 161– 167. DOI: 10.1111/j.1751-2980.2011.00570.x. PMID: 22356311
34. Tao Y.F., Teng F., Wang Z.X., Guo W.Y., Shi X.M., Wang G.H., Ding G.S., Fu Z.R. Liver transplant recipients with portal vein thrombosis: a single center retrospective study. Hepatobiliary Pancreat. Dis. Int. 2009; 8 (1): 34–39. PMID: 19208512
35. Esmat Gamil M., Pirenne J., Van Malenstein H., Verhaegen M., Desschans B., Monbaliu D., Aerts R., Laleman W., Cassiman D., Verslype C., Van Steenbergen W., Van Pelt J., Nevens F. Risk factors for bleeding and clinical implications in patients undergoing liver transplantation. Transplant. Proc. 2012; 44 (9): 2857–2860. DOI: 10.1016/j.transproceed.2012.09.085. PMID: 23146542
Transplant. Proc. 2012; 44 (9): 2857–2860. DOI: 10.1016/j.transproceed.2012.09.085. PMID: 23146542
36. Varotti G., Santori G., Andorno E., Morelli N., Ertreo M., Strada P., Porcile E., Casaccia M., Centanaro M., Valente U. Impact of Model for End-Stage Liver Disease score on transfusion rates in liver transplantation. Transplant. Proc. 2013; 45 (7): 2684–2688. DOI: 10.1016/j.transproceed.2013.07.006. PMID: 24034024
37. De Santis G.C., Brunetta D.M., Nardo M., Oliveira L.C., Souza F.F., Cagnolati D., Mente К.D., Sankarankutty A.K., Covas D.T., de Castro e Silva O. Preoperative variables associated with transfusion requirements in orthotopic liver transplantation. Transfus. Apher. Sci. 2014; 50 (1): 99–105. DOI: 10.1016/j.transci.2013.10.006. PMID: 24291115
38. Cywinski J.B., Alster J.M., Miller C., Vogt D.P., Parker B.M. Prediction of intraoperative transfusion requirements during orthotopic liver transplantation and the influence on postoperative patient survival. Anesth. Analg. 2014; 118 (2): 428–437. DOI: 10.1213/ANE.0b013e3182a76f19. PMID: 24445640
Analg. 2014; 118 (2): 428–437. DOI: 10.1213/ANE.0b013e3182a76f19. PMID: 24445640
39. Ramos E., Dalmau A., Sabate A., Lama C., Llado L., Figueras J., Jaurrieta E. Intraoperative red blood cell transfusion in liver transplantation: influence on patient outcome, prediction of requirements, and measures to reduce them. Liver Transpl. 2003; 9 (12): 1320–1327. DOI: 10.1016/jlts.2003.50204. PMID: 14625833
40. Fan S.T., Yong B.H., Lo C.M., Liu C.L., Wong J. Right lobe living donor liver transplantation with or without venovenous bypass. Br. J. Surg. 2003; 90 (1): 48–56. DOI: 10.1002/bjs.4026. PMID: 12520574
41. Busque S., Esquivel C.O., Concepcion W., So S.K. Experience with the piggyback technique without caval occlusion in adult orthotopic liver transplantation. Transplantation. 1998; 65 (1): 77–82. DOI: 10.1097/00007890-199801150-00015. PMID: 9448148
42. Hosein Shokouh-Amiri M., Osama Gaber A., Bagous W.A., Grewal H.P., Hathaway D.K., Vera S.R., Stratta R.J., Bagous T. N., Kizilisik T. Choice of surgical technique influences perioperative outcomes in liver transplantation. Ann. Surg. 2000; 231 (6): 814–823. DOI: 10.1097/00000658200006000-00005. PMID: 10816624
N., Kizilisik T. Choice of surgical technique influences perioperative outcomes in liver transplantation. Ann. Surg. 2000; 231 (6): 814–823. DOI: 10.1097/00000658200006000-00005. PMID: 10816624
43. Nishida S., Nakamura N., Vaidya A., Levi D.M., Kato T., Nery J.R., Madariaga J.R., Molina E., Ruiz P., Gyamfi A., Tzakis A.G. Piggyback technique in adult orthotopic liver transplantation: an analysis of 1067 liver transplants at a single center. HPB (Oxford). 2006; 8 (3): 182–188. DOI: 10.1080/13651820500542135. PMID: 18333273
44. Senzolo M., Burra P., Cholongitas E., Burroughs A.K. New insights into the coagulopathy of liver disease and liver transplantation. World J. Gas troenterol. 2006; 12 (48): 7725–7736. DOI: 10.3748/wjg.v12.i48.7725. PMID: 17203512
45. Журавель С.В., Андрейцева О.И., Донова Л.В., Джаграев К.Р., Чжао А.В. Профилактика и лечение сосудистых осложнений после трансплантации печени. Трансплантология. 2012; 1-2: 33-37.
46. Bakker C.M., Metselaar H.J., Groenland T. N., Terpstra O.T., Stibbe J. Increased fibrinolysis in orthotopic but not in heterotopic liver transplantation: the role of the anhepatic phase. Transpl. Int. 1992; 5 (Suppl 1): S173-S174. DOI: 10.1007/978-3-642-77423-2_54. PMID: 14621767
N., Terpstra O.T., Stibbe J. Increased fibrinolysis in orthotopic but not in heterotopic liver transplantation: the role of the anhepatic phase. Transpl. Int. 1992; 5 (Suppl 1): S173-S174. DOI: 10.1007/978-3-642-77423-2_54. PMID: 14621767
47. Bakker C.M., Metselaar H.J., Groenland T.N., Gomes M.J., Knot E.A., Hesselink E.J., Schalm S.W., Stibbe J., Terpstra O.T. Increased tissue-type plasminogen activator activity in orthotopic but not heterotopic liver transplantation: the role of the anhepatic period. Hepatology. 1992; 16 (2): 404-408. DOI: 10.1002/hep.1840160219. PMID: 1639350
48. Azad S.C., Kratzer M.A., Groh J., Welte M., Haller M., Pratschke E. Intraoperative monitoring and postoperative reevaluation of hemostasis in orthotopic liver transplantation. Semin. Thromb. Hemost. 1993; 19 (3): 233-237. DOI: 10.1055/s-2007-994031. PMID: 8362253
49. Paulsen A.W., Whitten C.W., Ramsay M.A., Klintmalm G.B. Considerations for anesthetic management during veno-venous bypassin adult hepatic transplantation. Anesth. Analg. 1989; 68 (4): 489-496. DOI: 10.1213/00000539-198904000-00012. PMID: 2648895
Anesth. Analg. 1989; 68 (4): 489-496. DOI: 10.1213/00000539-198904000-00012. PMID: 2648895
50. Scherer R., Giebler R., Erhard J., Lange R., Günnicker M., Schmutzler M., Paar D., Kox W.J. A new method of veno-venous bypass during human orthotopic liver transplantation. Anaesthesia. 1994; 49 (5): 398-402. DOI: 10.1111/j.1365-2044.1994.tb03471.x. PMID: 8209979
51. Shimauchi T., Yamaura K., Higashi M., Abe K., Yoshizumi T., Hoka S. Fibrinolysis in living donor liver transplantation recipients evaluated using thromboelastometry: impact on mortality. Transplant. Proc. 2017; 49 (9): 2117-2121. DOI: 10.1016/j.transproceed.2017.09.025. PMID: 29149971
52. Bakker C.M., Blankensteijn J.D., Schlejen P., Porte R.J., Gomes M.J., Lampe H.I., Stibbe J., Terpstra O.T. The effects of long-term graft preservation on intraoperative hemostatic changes in liver transplantation. A comparison between orthotopic and heterotopic transplantation in the pig. HPB Surg. 1994; 7 (4): 265-280. DOI: 10. 1155/1994/27915. PMID: 8204546
1155/1994/27915. PMID: 8204546
53. Wolberg A.S., Meng Z.H., Monroe D.M.3rd, Hoffman M. A systematic evaluation of the effect of temperature on coagulation enzyme activity and platelet function. J. Trauma. 2004; 56 (6): 1221–1228. DOI: 10.1097/01.TA.0000064328.97941.FC. PMID: 15211129
54. Martini W.Z. Coagulopathy by hypothermia and acidosis: mechanisms of thrombin generation and fibrinogen availability. J. Trauma 2009; 67 (1): 202–208. DOI: 10.1097/TA.0b013e3181a602a7. PMID: 19590336
55. Han S.B., Gwak M.S., Choi S.J., Kim M.H., Ko J.S., Kim G.S., Joo H.S. Effect of active airway warming on body core temperature during adult liver transplantation. Transplant. Proc. 2013; 45(1): 251-254. DOI: 10.1016/j.transproceed.2012.05.088. PMID: 23375310
56. Ghadimi K., Levy J.H., Welsby I.J. Perioperative management of the bleeding patient. Br. J. Anaesth. 2016; 117 (Suppl 3): iii18-iii30. DOI: 10.1093/bja/aew358. PMID: 27940453
57. Reynolds L., Beckmann J., Kurz A. Perioperative complications of hypothermia. Best Pract. Res. Clin. Anaesthesiol. 2008; 22 (4): 645-657. DOI: 10.1016/j.bpa.2008.07.005. PMID: 19137808
58. Pandey C.K., Singh A., Kajal K., Dhankhar M., Tandon M., Pandey V.K., Karna S.T. Intraoperative blood loss in orthotopic liver transplantation: the predictive factors. World J. Gastrointest. Surg. 2015; 7 (6): 86-93. DOI: 10.4240/wjgs.v7.i6.86. PMID: 26131330
59. Senzolo M., Cholongitas E., Thalheimer U., Riddell A., Agarwal S., Mallett S., Ferronato C., Burroughs A.K. Heparin-like effect in liver disease and liver transplantation. Clin. Liver Dis. 2009; 13 (1): 43-53. DOI: 10.1016/j.cld.2008.09.004. PMID: 19150308
60. Tomescu D., Popescu M., Jipa L., Fota R., Ungureanu D., Zamfir R., Orban C., Dima S.O., Popescu I. The impact of donor liver graft quality on postoperative outcome in liver transplant recipients. A single centre experience. Rom. J. Anaesth. Intensive Care. 2016; 23 (1): 19-26. DOI: 10.21454/rjaic.7518.231.gft. PMID: 28913473
61. Moriau M. , Kestens P.J., Masure R. Heparin and antifibrinolytic agents during experimental hepatectomy and liver transplantation. Pathol. Eur. 1969; 4 (2): 172–182. PMID: 4896202
62. Porte R.J., Bontempo F.A., Knot E.A., Lewis J.H., Kang Y.G., Starzl T.E. Systemic effects of tissue plasminogen activator-associated fibrinolysis and its relation to thrombin generation in orthotopic liver transplantation. Transplantation. 1989; 47 (6): 978-984. DOI: 10.1097/00007890198906000-00012. PMID: 2499962
63. Hutchison D.E., Genton E., Porter K.A., Daloze P.M., Huguet C., Brettschneider L., Groth C.G., Starzl T.E. Platelet changes following clinical and experimental hepatic homotransplantation. Arch. Surg. 1968; 97 (1): 27–33. DOI: 10.1001/archsurg.1968.01340010057003. PMID: 4232038
64. Reuvers C.B., Terpstra O.T., Groenland T.H., Boks A.L., Faithfull N.S., ten Kate F.W. Hemodynamics and coagulation in experimental auxiliary liver transplantation during fulminant hepatic failure. Ann. Surg. 1986; 204 (5): 552-558. DOI: 10.1097/00000658-198611000-00008. PMID: 3532973
65. Kang Y., Audu P. Coagulation and liver transplantation. Int. Anesthesiol. Clin. 2006; 44 (4): 17-36. DOI: 10.1097/01.aia.0000210811.77663.1e. PMID: 17033477
66. Шумаков В.И. (ред.). Трансплантология. М.: МИА; 2006: 310-330. ISBN: 5-89481-453-7
67. Schrem H., Klußmann A., Focken M., Emmanouilidis N., Oldhafer F., Klempnauer J., Kaltenborn A. Post-operative hemorrhage after liver transplantation: risk factors and long-term outcome. Ann. Transplant. 2016; 21: 46-55. DOI: 10.12659/AOT.895605. PMID: 26818716
68. Yuki K., Bu W., Shimaoka M., Eckenhoff R. Volatileanesthetics, not intravenous anesthetic propofol bind to and attenuate the activation of platelet receptor integrin αIIbβ3. PLoSOne. 2013; 8 (4): e60415. DOI: 10.1371/journal.pone.0060415. PMID: 23573252
69. Faraday N. Platelets, perioperative hemostasis, and anesthesia. Anesthesiology. 2002; 96 (5): 1042-1043. DOI: 10.1097/00000542-20020500000003. PMID: 11981140
70. Ueda I. The effects of volatile general anesthetics on adenosine diphosphateinduced platelet aggregation. Anesthesiology. 1971; 34 (5): 405408. DOI: 10.1097/00000542-197105000-00006. PMID: 5554584
71. Журавель С.В., Чугунов А.О., Салиенко А.А., Донова Л.В. Трансплантация печени, осложненная острым тромбозом воротной вены. Вестн. интенс. терапии. 2009; 3: 70-72.
72. Abuelkasem E., Lu S., Tanaka K., Planinsic R., Sakai T. Comparison between thrombelastography and thromboelastometry in hyperfibrinolysis detection during adult liver transplantation..Br. J. Anaesth. 2016; 116 (4): 507-512. DOI: 10.1093/bja/aew023. PMID: 26934942
73. Ozier Y., Pessione F., Samain E., Courtois F.; French Study Group on Blood Transfusion in Liver Transplantation. Institutional variability in transfusion practice for liver transplantation. Anesth. Analg. 2003; 97 (3): 671– 679. DOI: 10.1213/01.ANE.0000073354.38695.7C. PMID: 12933381
74. Hutten B.A., Lensing A.W., Kraaijenhagen R. A., Prins M.H. Safety of treatment with oral anticoagulants in the elderly. A systematic review. Drugs Aging. 1999; 14 (4): 303-312. DOI: 10.2165/00002512-19991404000005. PMID: 10319243
75. Anand S.S., Bates S., Ginsberg J.S., Levine M., Buller H., Prins M., Haley S., Kearon C., Hirsh J., Gent M. Reccurent venous thrombsis and heparin therapy: an evaluation of the importance of early activated partial tromboplastin times. Arch. Intern. Med. 1999; 159 (17): 2029-2032. DOI: 10.1001/archinte.159.17.2029. PMID: 10510988
76. Algarni A.A., Mourad M.M., Bramhall S.R. Anticoagulation and antiplatelets as prophylaxis for hepatic artery thrombosis after liver transplantation. World J. Hepatol. 2015; 7 (9): 1238-1243. DOI: 10.4254/wjh.v7.i9.1238. PMID: 26019738
77. Wall W.J., Grant D.R., Duff J.H., Kutt J.L., Ghent C.N. Blood transfusion requirements and renal function in patient undergoing liver transplantation without venous bypass. Transplant. Proc. 1987; 19 (4 Suppl 3): 1720. PMID: 3303529
78. de Boer M.T., Molenaar I.Q., Hendriks H.G., Slooff M.J., Porte R.J. Minimizing blood loss in liver transplantation: progress through research and evolution of techniques. Dig. Surg. 2005; 22 (4): 265-275. DOI: 10.1159/000088056. PMID: 16174983
79. de Boer M.T., Christensen M.C., Asmussen M., van der Hilst C.S., Hendriks H.G., Slooff M.J., Porte R.J. The impact of intraoperative transfusion of platelets and red blood cells on survival after liver transplantation. Anesth. Analg. 2008; 106 (1): 32–44. DOI: 10.1213/01.ane.0000289638. 26666.ed. PMID: 18165548
80. Cap A.P., Spinella P.C., Borgman M.A., Blackbourne L.H., Perkins J.G. Timing and location of blood product transfusion and outcomes in massively transfused combat casualties. J. Trauma Acute Care Surg. 2012; 73 (2 Suppl 1): S89–S94. DOI: 10.1097/TA.0b013e318260625a. PMID: 22847102
81. Phillips S.D., Maguire D., Deshpande R., Muiesan P., Bowles M.J., Rela M., Heaton N.D. A prospective study investigating the cost effectiveness of intraoperative blood salvage during liver transplantation. Transplantation. 2006; 81 (4): 536–540. DOI: 10.1097/01.tp.0000199318.17013.c5. PMID: 16495800
82. Massicotte L., Thibeault L., Beaulieu D., Roy J.D., Roy A. Evaluation of cell salvage autotransfusion utility during liver transplantation. HPB (Oxford). 2007; 9 (1): 52–57. DOI: 10.1080/13651820601090596. PMID: 18333113
83. Massicotte L., Thibeault L., Roy A. Classical notions of coagulation revisited in relation with blood losses, transfusion rate for 700 consecutive liver transplantations. Semin. Thromb. Hemost. 2015; 41 (5): 538-546. DOI: 10.1055/s-0035-1550428. PMID: 26080304
84. Kırnap M., Tezcaner T., Ayvazogl˘u Soy H.E., Akdur A., Yildirim S., Torgay A., Moray G., Haberal M. Efficacy of cell saver use in living-donor liver transplant. Exp. Clin. Transplant. 2015; 13 (Suppl 1): 315–317. DOI: 10.6002/ect.mesot2014.P153. PMID: 25894181
85. Liang T.B., Li D.L., Liang L., Li J.J., Bai X.L., Yu W., Wang W.L., Shen Y., Zhang M., Zheng S.S. Intraoperative blood salvage during liver transplan tation in patients with hepatocellular carcinoma: efficiency of leukocyte depletion filters in the removal of tumor cells. Transplantation. 2008; 85 (6): 863-869. DOI: 10.1097/TP.0b013e3181671f2e. PMID: 18360269
86. Araujo R.L., Pantanali C.A., Haddad L., Rocha Filho J.A., D’Albuquerque L.A., Andraus W. Does autologous blood transfusion during liver transplantation for hepatocellular carcinoma increase risk of recurrence? World J. Gastrointest. Surg. 2016; 8 (2): 161-168. DOI: 10.4240/wjgs.v8.i2.161. PMID: 26981190
87. Freire M.P., Soares Oshiro I.C., Bonazzi P.R., Guimarães T., Ramos Figueira E.R., Bacchella T., Costa S.F., Carneiro D’Albuquerque L.A., Abdala E. Surgical site infections in liver transplant recipients in the model for endstage liver disease era: an analysis of the epidemiology, risk factors, and outcomes. Liver Transpl. 2013; 19 (9): 1011–1019. DOI: 10.1002/lt.23682. PMID: 23744748
88. Reichert B., Kaltenborn A., Becker T., Schiffer M., Klempnauer J., Schrem H. Massive blood transfusion after the first cut in liver transplantation predicts renal outcome and survival. Langenbecks Arch. Surg. 2014; 399 (4): 429–440. DOI: 10.1007/s00423-014-1181-y. PMID: 24682384
89. Мороз В.В., Черныш А.М., Козлова Е.К., Сергунова В.А., Гудкова О.Е., Хорошилов С.Е., Онуфриевич А.Д., Костин А.И. Нарушения морфологии и наноструктуры мембран эритроцитов при длительном хранении эритроцитарной взвеси (исследование при помощи атомной силовой микроскопии). Бюл. эксперим. биологии и медицины. 2015; 159 (3): 390-394. DOI: 10.1007/s10517-015-2975-9. PMID: 26212816
90. Kozlova E., Chernysh A., Moroz V., Sergunova V., Gudkova O., Manchenko E. Morphology, membrane nanostructure and stiffness for quality assessment of packed red blood cells. Sci. Rep. 2017; 7 (1): 7846. DOI: 1038/s41598-017-08255-9. PMID: 28798476
91. Сергунова В.А., Гудкова О.Е., Манченко Е.А., Козлова Е.К., Бобринская И.Г., Черныш А.М., Козлов А.П. Влияние температуры эритроцитарной взвеси на морфологию и наноструктуру мембран клеток. Общая реаниматология. 2017; 13 (4): 30-37. DOI: 10.15360/1813-9779-20174-30-37
92. Wang S.C., Shieh J.F., Chang K.Y., Chu Y.C., Liu C.S., Loong C.C., Chan K.H., Mandell S., Tsou M.Y. Thromboelastography-guided transfusion decreases intraoperative blood transfusion during orthotopic liver transplantation: randomized clinical trial. Transplant. Proc. 2010; 42 (7): 2590–2593. DOI: 10.1016/j.transproceed.2010.05.144. PMID: 20832550
93. Hendriks H.G., Meijer K., de Wolf J.T., Klompmaker I.J., Porte R.J., de Kam P.J., Hagenaars A.J., Melsen T., Slooff M.J., van der Meer J. Reduced transfusion requirements by recombinant factor VIIa in orthotopic liver transplantation: a pilot study. Transplantation. 2001; 71 (3): 402-405. DOI: 10.1097/00007890-200102150-00011. PMID: 11233901
94. Pugliese F., Ruberto F., Summonti D., Perrella S., Cappannoli A., Tosi A., D’alio A., Bruno K., Martelli S., Celli P., Morabito V., Rossi M., Berloco P.B, Pietropaoli P. Activated recombinant factor VII in orthotopic liver transplantation. Transplant. Proc. 2007; 39 (6): 1883-1885. DOI: 10. 1016/j.transproceed.2007.05.062. PMID: 17692642
95. Chavez-Tapia N.C., Alfaro-Lara R., Tellez-Avila F., Barrientos-Gutiérrez T., González-Chon O., Mendez-Sanchez N., Uribe M. Prophylactic activated recombinant factor VII in liver resection and liver transplantation: systematic review and meta-analysis. PLoS One. 2011; 6 (7): e22581. DOI: 10.1371/journal.pone.0022581. PMID: 21818342
96. Scheffert J.L., Taber D.J., Pilch N.A., McGillicuddy J.W., Baliga P.K., Chavin K.D. Timing of factor VIIa in liver transplantation impacts cost and clinical outcomes. Pharmacotherapy. 2013; 33 (5): 483-488. DOI: 10.1002/phar.1230. PMID: 23468226
97. Helfritz F.A., Stiefel P., Manns M.P., Klempnauer J., Lehner F., Lenzen H., Ciesek S. Use of recombinant factor VIIA as a risk factor for graft loss after orthotopic liver transplantation due to hepatic artery thrombosis. J. Bioequiv. Availab. 2016; 8: 8-12. DOI: 10.4172/jbb.1000259
98. Trzebicki J., Flakiewicz E., Kosieradzki M., Blaszczyk B. , Kołacz M., Jureczko L., Pacholczyk M., Chmura A., Lagiewska B., Lisik W., Wasiak D., Kosson D., Kwiatkowski A., Lazowski T. The use of thromboelastometry in the assessment of hemostasis during orthotopic liver transplantation reduces the demand for blood products. Ann. Transplant. 2010; 15 (3): 19–24. PMID: 20877262
99. Molenaar I.Q., Warnaar N., Groen H., Tenvergert E.M., Slooff M.J., Porte R.J. Efficacy and safety of antifibrinolytic drugs in liver transplantation: a systematic review and meta-analysis. Am. J. Transplant. 2007; 7 (1): 185–194. DOI: 10.1111/j.1600-6143.2006.01591.x. PMID: 17227567
100. Kozek-Langenecker S.A., Afshari A., Albaladejo P., Santullano C.A., De Robertis E., Filipescu D.C., Fries D., Görlinger K., Haas T., Imberger G., Jacob M., Lancé M., Llau J., Mallett S., Meier J., Rahe-Meyer N., Samama C.M., Smith A., Solomon C., Van der Linden P., Wikkelsø A.J., Wouters P., Wyffels P. Management of severe perioperative bleeding: guidelines from the European Society of Anaesthesiology. Eur. J. Anaesthesiol. 2013; 30 (6): 270–382. DOI: 10.1097/EJA.0b013e32835f4d5b. PMID: 23656742
101. Ogren M., Bergqvist D., Björck M., Acosta S., Eriksson H., Sternby N.H. Portal vein thrombosis: prevalence, patient characteristics and lifetime risk: a population study based on 23,796 consecutive autopsies. World J. Gastroenterol. 2006; 12 (13): 2115-2119. DOI: 10.3748/wjg.v12.i13.2115. PMID: 16610067
102. Хорошилов С.Е., Никулин А.В. Детоксикация при критических состояниях: понимание научной проблемы в XXI веке (обзор). Общая реаниматология. 2017; 13 (5): 85-108. DOI: 10.15360/18139779-2017-5-85-108
103. Trzebicki J., Kosieradzki M., Flakiewicz E., Kuzminska G., Wasiak D., Pacholczyk M., Lagiewska B., Lisik W., Kosson D., Kulik A., Chmura A., Lazowski T. Detrimental effect of aprotinin ban on amount of blood loss during liver transplantation: single-center experience. Transplant. Proc. 2011; 43 (5): 1725–1727. DOI: 10.1016/j.transproceed.2011.01.182. PMID: 21693266
104. Schofield N., Sugavanam A., Thompson K., Mallett S.V. No increase in blood transfusions during liver transplantation since the withdrawal of aprotinin. Liver Transpl. 2014; 20 (5): 584–590. DOI: 10.1002/lt.23839. PMID: 24481770
105. Massicotte L., Lenis S., Thibeault L., Sassine M.P., Seal R.F., Roy A. Effect of low central venous pressure and phlebotomy on blood product transfusion requirements during liver transplantations. Liver Transpl. 2006; 12 (1): 117-123. DOI: 10.1002/lt.20559. PMID: 16382461
106. Massicotte L., Beaulieu D., Thibeault L., Roy J.D., Marleau D., Lapointe R., Roy A. Coagulation defects do not predict blood product requirements during liver transplantation. Transplantation 2008; 85 (7): 956962. DOI: 10.1097/TP.0b013e318168fcd4. PMID: 18408574
107. Schroeder R.A., Collins B.H., Tuttle-Newhall E., Robertson K., Plotkin J., Johnson L.B., Kuo P.C. Intraoperative fluid management during orthotopic liver transplantation. J. Cardiothorac. Vasc. Anesth. 2004; 18 (4): 438–441. DOI: 10.1053/j.jvca.2004.05.020. PMID: 15365923
108. Colomina M.J., Díez Lobo A., Garutti I., Gómez-Luque A., Llau J.V., Pita E. Perioperative use of prothrombin complex concentrates. Minerva Anestesiol. 2012; 78: 358-368. PMID: 22357373
109. Arshad F., Ickx B., van Beem R.T., Polak W., Grüne F., Nevens F., Ilmakunnas M., Koivusalo A.M., Isoniemi H., Strengers P.F., Groen H., Hendriks H.G., Lisman T., Pirenne J., Porte R.J. Prothrombin complex concentrate in the reduction of blood loss during orthotopicliver transplantation: PROTON-trial. BMC Surg. 2013; 13: 22. DOI: 0.1186/1471-2482-1322. PMID: 23815798
110. Quintero J., Ortega J., Miserachs M., Bueno J., Bilbao I., Charco R. Low plasma levels of antithrombin III in the early post-operative period following pediatric liver transplantation: should they be replaced? A singlecenter pilot study. Pediatr. Transplant. 2014; 18 (2): 185-189. DOI: 10.1111/petr.12217. PMID: 24438318
111. Allingstrup M., Wetterslev J., Ravn F. B., Møller A.M., Afshari A. Antithrombin III for critically ill patients. Cochrane Database Syst. Rev. 2016; 2: CD005370. DOI: 10.1002/14651858.CD005370.pub3. PMID: 26858174
Анализ крови на гемостаз, гемостазиограмма — коагулограмма крови, цена исследования
Версия для печати
В состав данного комплекса входят следующие исследования:
Гемостазиологические исследования
- Протромбин (Prothrombin), Международное нормализованное отношение, МНО (International Normalized Ratio, INR)
- Фибриноген (Fibrinogen)
- Антитромбин III (Antithrombin III)
- Активированное частичное тромбопластиновое время (Activated Partial Thromboplastin Time)
- Тромбиновое время (Thrombin Time)
Биоматериал
Для данного исследования лаборатория принимает следующий биоматериал:
- Замороженная плазма (цитрат натрия)
Подготовка к исследованию
Взятие крови проводится натощак (не менее 8 и не более 14 ч голодания). Можно пить воду без газа. Если исследование назначается на фоне приема препаратов, влияющих на свертывание крови, необходимо отметить это в направлении.
Исследование системы гемостаза — один из самых широко применяемых тестов в современной медицине. Исследование показателей, характеризующих гемостаз (свёртывание крови), имеет большое значение в диагностике различных заболеваний (сердечнососудистая, акушерская, генетическая патология, системные заболевания, заболевания системы крови и др.), профилактике осложнений после оперативных вмешательств и др.
В программу входят основные показатели свертывающей и противосвертывающей (антикоагулянтной) системы крови.
Обращаем Ваше внимание на то, что интерпретация результатов исследований, установление диагноза, а также назначение лечения, в соответствии с Федеральным законом № 323-ФЗ «Об основах охраны здоровья граждан в Российской Федерации» от 21 ноября 2011 года, должны производиться врачом соответствующей специализации.
- Код:
-
пример
результата300006
можно сдать на дому
- Стоимость:
- + 190 р. Взятие крови
При единовременном заказе нескольких услуг, услуга по сбору биоматериала оплачивается только один раз.
1 877 р.
- Срок выполнения:
-
Указанный срок не включает день взятия биоматериала.
1 к.д.
результаты за 3-6 часов (CITO)
Серологическая диагностика для госпитализации
- результаты за 3-6 часов (CITO)
- Код:
- 300015
- Срок:
- 1 к. д.
Цена: 1784 р.
Общий анализ крови + СОЭ с лейкоцитарной формулой (с микроскопией мазка крови при наличии патологических сдвигов), венозная кровь
- результаты за 3-6 часов (CITO)
- Код:
- 110006
- Срок:
- 1 к.д.
Цена: 743 р.
Общий анализ мочи (Urine test) с микроскопией осадка
- результаты за 3-6 часов (CITO)
- Код:
- 110101
- Срок:
- 1 к. д.
Цена: 352 р.
Креатинин (Сreatinine)
- результаты за 3-6 часов (CITO)
- Код:
- 090004
- Срок:
- 1 к.д.
Цена: 286 р.
Мочевина (Urea)
- результаты за 3-6 часов (CITO)
- Код:
- 090005
- Срок:
- 1 к. д.
Цена: 286 р.
Показать еще
О возможных противопоказаниях необходимо проконсультироваться со специалистом
Расширенное исследование генов системы гемостаза (без описания результатов врачом-генетиком)
Метод определения Real-time-PCR.
Исследуемый материал Цельная кровь (с ЭДТА)
Доступен выезд на дом
Расширенное исследование генов системы гемостаза: F2, F5, MTHFR, MTR, MTRR, F13, FGB, ITGA2, ITGВ3, F7, PAI-1
Комплексное исследование генетических факторов риска развития нарушений в системе свертывания крови и фолатном цикле (без заключения врача-генетика).
Различные изменения в генах системы гемостаза и цикла обмена фолатов предрасполагают к развитию большого числа патологических состояний: инфаркты, инсульты, тромбоэмболии, кровотечения, патология беременности и родов, осложнения послеоперационного периода и т. д. Профиль включает в себя исследование основных полиморфизмов в генах системы гемостаза и фолатного цикла:
- F2 c.*97G>A (20210 G>A; rs1799963),
- F5 c.1601G>A (Arg534Gln; 1691 G>A; rs6025),
- MTHFR c.665C>T (Ala222Val; 677 C>T; rs1801133),
- MTHFR c.1286A>C (Glu429Ala; 1298 A>C; rs1801131),
- MTR c.2756A>G (Asp919Gly; rs1805087),
- MTRR c.66A>G (Ile22Met; rs1801394),
- F13 с.103G>T (I63Т; rs5985),
- FGB c.-467G>A (-455 G>А; rs1800790),
- ITGA2 c.759C>T (Phe253Phe, 807 C>T; rs1126643),
- ITGB3 c.176T>C (Leu59Pro; 1565 T>C; rs5918),
- F7 c.1238G>A (Arg353Gln; 10976 G>A; rs6046),
- PAI-1 (SERPINE1) –675 5G>4G (rs1799889).
Ген F2 кодирует аминокислотную последовательность белка протромбина. Полиморфизм F2 c.*97G>A приводит к повышенной экспрессии гена. Клинически неблагоприятный вариант полиморфизма (c. *97A) наследуется по аутосомно-доминантному типу. Наличие полиморфизма F2 c.*97G>A в гомозиготной или гетерозиготной форме значительно (в 3 и более раз, а на фоне курения — в 40 и более раз) увеличивает риск возникновения венозных тромбозов, в том числе тромбозов сосудов мозга и сердца, особенно в молодом возрасте. У пациентов-носителей данного полиморфизма повышен риск развития тромбоэмболий после хирургических вмешательств. Приём оральных контрацептивов у данной группы лиц также увеличивает риск тромбозов (относительный риск развития тромбофилии и венозной тромбоэмболии у гетерозиготных носительниц полиморфизма c.*97G>A возрастает в 16 раз).
Ген F5 кодирует аминокислотную последовательность белка проакцелерина — коагуляционного фактора 5. Нуклеотидная замена c.1601G>A («мутация Лейден») приводит к аминокислотной замене аргинина на глутамин в позиции 534, что придает устойчивость активной форме проакцелерина. Клинически это проявляется рецидивирующими венозными тромбозами и тромбоэмболиями. Наличие полиморфизма в гомозиготной или гетерозиготной форме значительно (в 3 и более раз, а на фоне заместительной гормонотерапии или приема оральных контрацептивов — в 30 и более раз) увеличивает риск венозных тромбозов. Риск инфаркта миокарда увеличивается в 2 и более раз, риск развития патологии беременности (прерывание беременности, преэклампсия, хроническая плацентарная недостаточность и синдром задержки роста плода) увеличивается в 3 и более раз.
Также, пациенты, являющиеся одновременно носителями полиморфизма c.*97G>A гена протромбина и «мутации Лейден», еще в большей степени подвержены риску развития тромбозов и тромбоэмболий.
Ген MTHFR кодирует аминокислотную последовательность фермента метилентетрагидрофолатредуктазы, играющего ключевую роль в метаболизме фолиевой кислоты. Полиморфизм c.665C>T гена MTHFR связан с заменой нуклеотида цитозина (С) на тимин (Т), что приводит к аминокислотной замене аланина на валин в позиции 222. Вариант c.665Т связан с четырьмя группами мультифакториальных заболеваний: сердечно-сосудистыми, дефектами развития плода, колоректальной аденомой и раком молочной железы и яичников. У женщин с генотипом c.665Т/Т дефицит фолиевой кислоты во время беременности может приводить к порокам развития плода, в том числе незаращению нервной трубки. Неблагоприятное воздействие варианта c.665Т- зависит от внешних факторов: низкого содержания в пище фолатов, курения, приема алкоголя. Сочетание генотипа c.665Т/Т и папилломавирусной инфекции увеличивает риск цервикальной дисплазии. Назначение препаратов фолиевой кислоты может значительно снизить негативное влияние данного варианта полиморфизма.
Полиморфизм MTHFR c.1286A>C связан с точечной заменой нуклеотида аденина (А) на цитозин (С), что приводит к замене аминокислотного остатка глутаминовой кислоты на аланин в позиции 429, относящейся к регулирующей области молекулы фермента. При наличии данного полиморфизма отмечается снижение активности фермента MTHFR. Это снижение обычно не сопровождается изменением уровня гомоцистеина в плазме крови у носителей дикого варианта полиморфизма c.665C>T, однако сочетание аллельного варианта* c.1286C с аллелем c.665T приводит к снижению уровня фолиевой кислоты и соответствует по своему эффекту гомозиготному состоянию MTHFR c.665Т/T. При этом риск развития дефектов нервной трубки повышается в 2 раза. Жизнеспособность плодов, имеющих одновременно оба полиморфных варианта, также снижена.
Ген MTR кодирует аминокислотную последовательность фермента метионин синтазы. Полиморфизм c.2756A>G связан с аминокислотной заменой (аспарагиновой кислоты на глицин) в молекуле фермента. В результате этой замены функциональная активность фермента изменяется, что приводит к повышению риска формирования пороков развития у плода. Влияние полиморфизма усугубляется повышенным уровнем гомоцистеина.
Ген MTRR кодирует аминокислотную последовательность фермента редуктазы метионинсинтазы. Полиморфизм c.66A>G связан с аминокислотной заменой в молекуле фермента. В результате этой замены функциональная активность фермента снижается, что приводит к повышению риска развития дефектов нервной трубки у плода. Влияние полиморфизма усугубляется дефицитом витамина В12. При сочетании полиморфизма c.66A>G гена MTRR с полиморфизмом c.665C>T в гене MTHFR риск spina bifida увеличивается. Полиморфизм c.66A>G гена MTRR усиливает гипергомоцистеинемию, вызываемую полиморфизмом c.665C>T в гене MTHFR.
Ген фибриназы (F13) кодирует синтез трансглютаминазы, участвующей в стабилизации фибринового сгустка и в формировании соединительной ткани. Аллельные варианты с.103G/Т и с.103Т/Т приводят к снижению уровня трансглютаминазы с образованием сетчатой структуры фибрина с более тонкими волокнами, меньшими порами, и изменением характеристик проникновения, которое в сочетании с другими факторами риска ассоциируется с возможным риском внутричерепных кровоизлияний и кровотечений из внутренних органов, а также привычным невынашиванием беременности. При этом аллельный вариант с.103Т может выступать в роли протективного фактора в отношении инфаркта миокарда и венозных тромбозов.
Ген FGB кодирует β-цепь фибриногена, являющегося предшественником фибрина. Аллельный вариант c.-467А обусловливает усиленную транскрипцию гена и может приводить к увеличению уровня фибриногена в крови и повышению вероятности образования тромбов при наличии дополнительных факторов риска. Гетерозиготный вариант c.-467G/А связывают с повышенным риском ишемического инсульта и лакунарными инфарктами церебральных сосудов. Гомозиготный вариант c.-467A/А связывают с повышенным риском инфаркта миокарда.
Ген гликопротеина Gp1a (ITGA2) кодирует синтез альфа-2-субъединицы интегринов – специализированных рецепторов тромбоцитов. Аллельный вариант c.759Т вызывает изменение первичной структуры субъединицы и свойств рецепторов. При гетерозиготном (c.759C/T) варианте отмечается увеличение скорости адгезии тромбоцитов к коллагену I типа, что может приводить к повышенному риску тромбофилии, инфаркта миокарда и других сердечно-сосудистых заболеваний. Аллельный вариант c.759Т связывают со случаями резистентности к аспирину. Помимо этого, при гомозиготном (c.759Т/T) варианте значительно увеличивается количество рецепторов на поверхности тромбоцитов. В совокупности, при гомозиготном варианте данного полиморфизма значительно повышен риск тромбофилии, инфаркта миокарда и развития других острых эпизодов тромбообразования в возрасте до 50 лет, даже по сравнению с гетерозиготным вариантом.
Ген гликопротеина Gp3a (ITGB3) кодирует синтез бета-3 цепи интегринового комплекса GP2b\3a, участвующего в разнообразных межклеточных взаимодействиях (адгезии и сигнализации). Аллельный вариант c.176С (гетерозигота c.176T/C) обусловливает повышенную адгезию тромбоцитов и может приводить к увеличению риска развития острого коронарного синдрома, а также связан с синдромом привычного невынашивания беременности. Гомозиготный вариант c.176С/C обусловливает повышенную адгезию тромбоцитов и может приводить к значительному увеличению риска развития острого коронарного синдрома в возрасте до 50 лет. У лиц с полиморфными аллельными вариантами часто отмечается пониженная эффективность аспирина.
Аллельный вариант c.1238A (гетерозигота c.1238G/A и гомозигота c.1238А/A) гена F7 приводит к понижению экспрессии гена и снижению уровня фактора 7 в крови, рассматривается как протективный маркёр в отношении развития тромбозов и инфаркта миокарда.
Ген ингибитора активатора плазминогена (PAI-1) кодирует белок-антагонист тканевого и урокиназного активатора плазминогена. Преобладающим в популяции вариантом исследуемого полиморфизма является гетерозиготный вариант -675 5G/4G. В связи с этим данный полиморфизм самостоятельного диагностического значения не имеет, эффект возможно оценить в сочетании с другими факторами предрасполагающими к развитию патологии (например в сочетании с FGB c.-467A). Аллельный вариант -675 4G сопровождается большей активностью гена, чем -675 5G, что обусловливает более высокую концентрацию PAI-1 и уменьшение активности противосвёртывающей системы. Гомозигота -675 4G/4G ассоциирована с повышением риска тромбообразования, преэклампсии, нарушением функции плаценты и самопроизвольного прерывания беременности.
*Примечание: иногда в научной литературе при описании однонуклеотидных замен, характерных для генных полиморфизмов, встречается термин «мутантный аллель». Это терминологическая неточность, так как в классической генетике термин «мутантный аллель» традиционно рассматривается как синоним термина «мутация». При мутациях, как известно, изменение структуры гена приводит к образованию (экспрессии) нефункциональных белков и к неизбежному развитию наследственного заболевания. При полиморфизмах изменение в структуре гена приводит лишь к появлению белков с немного изменёнными физико-химическими свойствами. Такие изменения, как известно, проявляют себя при воздействии на организм различных факторов внешней среды или при изменении функционального состояния организма человека. И только в таких ситуациях функционирование белков со структурными особенностями может, либо способствовать ускорению развития заболевания, либо, напротив, тормозить формирование патологических процессов. Поэтому, на наш взгляд, для разграничения изменений в генах столь очень похожих структурно, но приводящих к несоизмеримо разным последствиям для организма, корректнее в отношении генных полиморфизмов применять понятие «аллельный вариант гена», а не «мутантный аллель».
Регуляция центральной нервной системой энергии организма и гомеостаза глюкозы
Бек, Б. и Ричи, С. Диетическая модуляция грелина и лептина и поведение при переедании после потери веса у крыс Цукера с ожирением. Дж. Эндокринол. 202 , 29–34 (2009).
КАС Статья пабмед Google ученый
Dulloo, A.G. & Calokatisa, R. Адаптация к низкому потреблению калорий у мышей с ожирением: вклад метаболического компонента в снижение расхода энергии во время и после потери веса. Междунар. Дж. Обес. 15 , 7–16 (1991).
КАС пабмед Google ученый
MacLean, P. S. et al. Периферические метаболические реакции на продолжительное снижение веса, которые способствуют быстрому и эффективному восстановлению у склонных к ожирению крыс. утра. Дж. Физиол. Регул. интегр. Комп. Физиол. 290 , R1577–R1588 (2006 г.).
КАС Статья пабмед Google ученый
Мелби, С. Л., Пэрис, Х. Л., Форайт, Р. М. и Пет, Дж. Ослабление биологического стремления к набору веса после потери веса: должно ли то, что снижается, всегда возвращаться обратно? Питательные вещества 9 , 468 (2017).
Центральный пабмед Статья КАС Google ученый
Мортон Г.Дж., Каммингс Д.Е., Баскин Д.Г., Барш Г.С. и Шварц М.В. Центральная нервная система контролирует потребление пищи и массу тела. Природа 443 , 289–295 (2006).
КАС Статья пабмед Google ученый
Williams, G. et al. Гипоталамус и контроль энергетического гомеостаза разные цепи, разные цели. Физиол. Поведение 74 , 683–701 (2001).
КАС Статья пабмед Google ученый
Аонге, К. М., Д’Алессио, Д. А. и Шварц, М. В. Мозговой контроль уровня глюкозы в крови: последствия для патогенеза диабета 2 типа. Диабетология https://doi.org/10.1007/s00125-020-05293-3 (2020).
Рикелс, М. Р. и Робертсон, Р. П. Трансплантация островков поджелудочной железы у людей: недавний прогресс и направления на будущее. Эндокр. Ред. 40 , 631–668 (2019).
Артикул пабмед Google ученый
Браун, Дж. М., Скарлетт, Дж. М. и Шварц, М. В. Переосмысление роли мозга в гомеостазе глюкозы и патогенезе диабета. Дж. Клин. Инвестировать . 129 , 3035–3037 (2019).
Faber, C.L., Deem, J.D., Campos, C.A., Taborsky, G.J. & Morton, G.J. Контроль ЦНС эндокринной поджелудочной железы. Диабетология 63 , 2086–2094 (2020).
ПабМед ПабМед Центральный Статья Google ученый
Смит, С. и Херон, А. Диабет и ожирение: две эпидемии. Нац. Мед. 12 , 75–80 (2006).
КАС Статья пабмед Google ученый
Considine, R. V. et al. Концентрации иммунореактивного лептина в сыворотке у людей с нормальным весом и ожирением. Н. англ. Дж. Мед. 334 , 292–295 (1996).
КАС Статья пабмед Google ученый
Frederich, R.C. et al. Уровни лептина отражают содержание липидов в организме у мышей: свидетельство индуцированной диетой устойчивости к действию лептина. Нац. Мед. 1 , 1311–1314 (1995).
КАС Статья пабмед Google ученый
Hummel, K.P., Dickie, M.M. & Coleman, D.L. Диабет, новая мутация у мышей. Наука 153 , 1127–1128 (1966).
КАС Статья пабмед Google ученый
Tartaglia, L. A. et al. Идентификация и клонирование экспрессии рецептора лептина, OB-R. Cell 83 , 1263–1271 (1995).
КАС Статья пабмед Google ученый
Ингаллс, А. М., Дики, М. М. и Снелл, Г. Д. Ожирение, новая мутация у домовой мыши. Дж. Херед. 41 , 317–318 (1950).
КАС Статья пабмед Google ученый
Kennedy, G.C. Роль депо жира в гипоталамическом контроле потребления пищи у крыс. Проц. Р. Соц. Лонд. Б биол. науч. 140 , 578–596 (1953).
КАС Статья пабмед Google ученый
Батлер, П. К. и др. Влияние приема пищи на концентрацию амилина в плазме у людей с NIDDM и без диабета. Диабет 39 , 752–756 (1990).
КАС Статья пабмед Google ученый
Elliott, R. M. et al. Секреция глюкагоноподобного пептида-1(7–36)амида и глюкозозависимого инсулинотропного полипептида в ответ на прием питательных веществ у человека: острая постпрандиальная и 24-часовая секреция. Дж. Эндокринол. 138 , 159–166 (1993).
КАС Статья пабмед Google ученый
GIBBS, J., YOUNG, R.C. & SMITH, G.P. Холецистокинин вызывает чувство сытости у крыс с открытыми желудочными фистулами. Природа 245 , 323–325 (1973).
КАС пабмед Статья Google ученый
Strubbe, J. & Steffens, A. Быстрое высвобождение инсулина после приема пищи у неанестезированной крысы. утра. Дж. Физиол. 229 , 1019–1022 (1975).
КАС пабмед Статья Google ученый
Berthoud, H.-R., Lynn, P.A. & Blackshaw, L.A. Блуждающие и спинальные механосенсоры в желудке и толстой кишке крысы имеют множественные рецептивные поля. утра. Дж. Физиол. 280 , R1371–R1381 (2001 г.).
КАС Google ученый
Филлипс, Р. Дж. и Паули, Т. Л. Рецепторы напряжения и растяжения в гладких мышцах желудочно-кишечного тракта: переоценка электрофизиологии механорецепторов блуждающего нерва. Мозг Res. Ред. 34 , 1–26 (2000).
КАС Статья пабмед Google ученый
Фрека, Дж. М. и Маттес, Р. Д. Возможное увлечение грелина обычными схемами приема пищи у людей. утра. Дж. Физиол. 294 , Г699–G707 (2008 г.).
КАС Google ученый
Рен, А. М. и др. Грелин усиливает аппетит и увеличивает потребление пищи у людей. Дж. Клин. Эндокринол. Метаб. 86 , 5992 (2001).
КАС Статья пабмед Google ученый
Johnen, H. et al. Индуцированная опухолью анорексия и потеря веса опосредованы цитокином MIC-1 суперсемейства TGF-β. Нац. Мед. 13 , 1333–1340 (2007).
КАС Статья пабмед Google ученый
Цай, В. В. В., Хусаини, Ю., Сейнсбери, А. , Браун, Д. А. и Брейт, С. Н. Путь MIC-1/GDF15-GFRAL в энергетическом гомеостазе: значение для ожирения, кахексии и других сопутствующих заболеваний. Сотовый метаб. 28 , 353–368 (2018).
КАС Статья пабмед Google ученый
Борнер, Т. и др. GDF15 вызывает анорексию через тошноту и рвоту. Сотовый метаб. https://doi.org/10.1016/j.cmet.2019.12.004 (2020 г.).
Кляйнерт, М. и др. Упражнения увеличивают циркулирующий GDF15 в организме человека. Мол. Метаб. 9 , 187–191 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Патель, С. и др. GDF15 обеспечивает эндокринный сигнал пищевого стресса у мышей и людей. Метаб. клеток 29 , 707–718 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Emmerson, P. J. et al. Метаболические эффекты GDF15 опосредованы орфанным рецептором GFRAL. Нац. Мед. 23 , 1215–1219 (2017).
КАС Статья пабмед Google ученый
Хсу, Дж.-Ю. и другие. Негомеостатическая регуляция массы тела через рецептор GDF15, ограниченный стволом мозга. Природа 550 , 255–259 (2017).
Артикул КАС пабмед Google ученый
Mullican, S.E. et al. GFRAL является рецептором для GDF15, и этот лиганд способствует снижению веса у мышей и нечеловеческих приматов. Нац. Мед. 23 , 1150–1157 (2017).
КАС Статья пабмед Google ученый
Ян, Л. и др. GFRAL является рецептором для GDF15 и необходим для действия лиганда против ожирения. Нац. Мед. 23 , 1158–1166 (2017).
КАС Статья пабмед Google ученый
Wischhusen, J., Melero, I. & Fridman, W. H. Фактор роста/дифференциации-15 (GDF-15): от биомаркера до нового целевого иммунного контрольного пункта. Фронт. Иммунол. 11 , 951 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Celi, F. S. et al. Минимальные изменения температуры окружающей среды приводят к значительному увеличению расхода энергии и изменению гормонального гомеостаза у здоровых взрослых людей. евро. Дж. Эндокринол. 163 , 863–872 (2010).
КАС пабмед ПабМед Центральный Статья Google ученый
Langeveld, M. et al. Мягкое воздействие холода на чувство голода, потребление пищи, чувство сытости и температуру кожи у людей. Эндокр. Соединять. 5 , 65–73 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Макари, М., Даунси, М.Дж. и Инграм, Д.Л. Изменения в потреблении пищи в ответ на изменения температуры окружающей среды: модификации предыдущего теплового и пищевого опыта. Арка Пфлюгера. 396 , 231–237 (1983).
КАС Статья пабмед Google ученый
Seeley, R. J. et al. Поведенческие, эндокринные и гипоталамические реакции на непроизвольное переедание. утра. Дж. Физиол. 271 , R819–R823 (1996).
КАС Статья пабмед Google ученый
Ravussin, Y. et al. Доказательства нелептиновой системы, которая защищает от увеличения веса при переедании. Сотовый метаб. 28 , 289–299 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Уайт, К. Л., Пурпера, М. Н., Баллард, К. и Моррисон, К. Д. Снижение потребления пищи после переедания включает лептин-зависимые и лептин-независимые механизмы. Физиол. Поведение 100 , 408–416 (2010).
КАС пабмед ПабМед Центральный Статья Google ученый
Коулман, Д. Л. Эффекты парабиоза тучных мышей с диабетом и нормальных. Диабетология 9 , 294–298 (1973).
КАС Статья пабмед Google ученый
Коулман, Д. Л. Ожирение и диабет: два мутантных гена, вызывающие синдромы диабета и ожирения у мышей. Диабетология 14 , 141–148 (1978).
КАС Статья пабмед Google ученый
Д’Алессио, Д. А., Киффер, Т. Дж., Таборски, Г. Дж. и Хавел, П. Дж. Активация парасимпатической нервной системы необходима для нормальной секреции инсулина, вызванной приемом пищи, у макак-резусов. Дж. Клин. Эндокринол. Метаб. 86 , 1253–1259 (2001).
Артикул пабмед Google ученый
Havel, P.J., Veith, R.C., Dunning, B.E. & Taborsky, G.J. Норадренергические нервы поджелудочной железы активируются нейроглюкопенией, но не гипотензией или гипоксией у собак. Доказательства специфичной для стресса и региональной избирательной активации симпатической нервной системы. Дж. Клин. Инвестировать. 82 , 1538–1545 (1988).
КАС пабмед ПабМед Центральный Статья Google ученый
Лернер, Р. Л. и Порт, Д. Исследования секретин-стимулированных реакций инсулина у человека. Дж. Клин. Инвестировать. 51 , 2205–2210 (1972).
КАС пабмед ПабМед Центральный Статья Google ученый
Росарио, В. и др. Карта нейронов островков головного мозга и поджелудочной железы показывает дифференциальную регуляцию глюкозы из разных областей гипоталамуса. Диабет 65 , 2711–2723 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Thorens, B. Нейронная регуляция массы и функции островковых клеток поджелудочной железы. Диабет Ожирение. Метаб. 16 , 87–95 (2014).
КАС Статья пабмед Google ученый
Берту, Х. Р., Берейтер, Д. А., Тримбл, Э. Р., Сигель, Э. Г. и Жанрено, Б. Головная фаза, рефлекторная секреция инсулина, нейроанатомическая и физиологическая характеристика. Diabetologia 20 , 393–401 (1981).
КАС Статья пабмед Google ученый
Тефф, К. Л. Как нервное опосредование упреждающего и компенсаторного высвобождения инсулина помогает нам переносить пищу. Физиол. Поведение 103 , 44–50 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Ahrén, B. Вегетативная регуляция секреции островковых гормонов – последствия для здоровья и болезней. Diabetologia 43 , 393–410 (2000).
Артикул пабмед Google ученый
Brochu, M. et al. Висцеральная жировая ткань является независимым коррелятом утилизации глюкозы у пожилых женщин с ожирением в постменопаузе. Дж. Клин. Эндокринол. Метаб. 85 , 2378–2384 (2000).
КАС пабмед Google ученый
Ким, К. и Парк, С. М. Связь мышечной массы и жировой массы с резистентностью к инсулину и распространенностью метаболического синдрома у взрослых корейцев: перекрестное исследование. Науч. Респ. 8 , 2703 (2018).
ПабМед ПабМед Центральный Статья КАС Google ученый
Faerch, K. et al. Естественная история чувствительности к инсулину и секреции инсулина при переходе от нормальной толерантности к глюкозе к нарушенной гликемии натощак и нарушенной толерантности к глюкозе: исследование Inter99. Diabetes Care 32 , 439–444 (2009 г.).
КАС пабмед ПабМед Центральный Статья Google ученый
Ретнакаран, Р. и др. Гиперболическая зависимость между секрецией инсулина и чувствительностью при пероральном тесте на толерантность к глюкозе. Ожирение 16 , 1901–1907 (2008).
КАС Статья пабмед Google ученый
Уолтон С., Годсланд И. Ф., Праудлер А. Дж., Фелтон С. В. и Винн В. Влияние индекса массы тела и распределения жира на чувствительность, секрецию и клиренс инсулина у здоровых мужчин без ожирения. Дж. Клин. Эндокринол. Метаб. 75 , 170–175 (1992).
КАС пабмед Google ученый
Morton, G.J. et al. Доказательства того, что симпатическая нервная система вызывает быструю, скоординированную и взаимную регулировку секреции инсулина и чувствительности к инсулину во время воздействия холода. Диабет 66 , 823–834 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Мута, К. и др. Глюкорегуляторные реакции на охлаждение преоптической области гипоталамуса. Мозг Res. 1710 , 136–145 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Диллон, Х. и др. Лептин напрямую активирует нейроны SF1 в VMH, и это действие лептина необходимо для нормального гомеостаза массы тела. Нейрон 49 , 191–203 (2006).
КАС Статья пабмед Google ученый
Flak, J. N. et al. Подмножество нейронов вентромедиального ядра гипоталамуса регулирует уровень глюкозы в крови независимо от инсулина. Дж. Клин. Инвестировать. 130 , 2943–2952 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
German, J.P. et al. Лептин активирует новый механизм ЦНС для независимой от инсулина нормализации тяжелой диабетической гипергликемии. Эндокринология 152 , 394–404 (2011).
КАС Статья пабмед Google ученый
Мик, Т. Х. и др. Действие лептина в вентромедиальном ядре гипоталамуса достаточно, но не обязательно для нормализации диабетической гипергликемии. Эндокринология 154 , 3067–3076 (2013).
КАС пабмед ПабМед Центральный Статья Google ученый
Балланд, Э. и др. Передача сигналов лептина в аркуатном ядре снижает способность инсулина подавлять выработку глюкозы в печени у мышей с ожирением. Cell Rep. 26 , 346–355 (2019).
КАС Статья пабмед Google ученый
Хашикава, К. и др. Клетки Esr1 + в вентромедиальном гипоталамусе контролируют агрессию самок. Нац. Неврологи. 20 , 1580–1590 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Krause, WC & Ingraham, HA Происхождение и функции вентролатерального VMH: сложный кластер нейронов, определяющий половые различия в метаболизме и поведении. в Половые и гендерные факторы, влияющие на метаболический гомеостаз, диабет и ожирение (изд. Mauvais-Jarvis, F.) 1043, 199–213 (Springer International Publishing, 2017).
Вандер Туиг, Дж. Г., Кнеханс, А. В. и Ромсос, Д. Р. Снижение активности симпатической нервной системы у крыс с поражениями вентромедиального гипоталамуса. Науки о жизни. 30 , 913–920 (1982).
Артикул Google ученый
Kunwar, P. S. et al. Вентромедиальные нейроны гипоталамуса контролируют защитное эмоциональное состояние. eLife 4 , e06633 (2015).
Центральный пабмед Статья Google ученый
Minokoshi, Y., Haque, M.S. & Shimazu, T. Микроинъекция лептина в вентромедиальный гипоталамус увеличивает поглощение глюкозы в периферических тканях у крыс. Диабет 48 , 287–291 (1999).
КАС Статья пабмед Google ученый
Toda, C. et al. Киназа, регулируемая внеклеточным сигналом, в вентромедиальном гипоталамусе опосредует индуцированное лептином поглощение глюкозы в скелетных мышцах красного типа. Диабет 62 , 2295–2307 (2013).
КАС пабмед ПабМед Центральный Статья Google ученый
Noble, E.E., Billington, C.J., Kotz, C.M. & Wang, C. Окситоцин в вентромедиальном ядре гипоталамуса снижает прием пищи и резко увеличивает расход энергии. утра. Дж. Физиол. 307 , R737–R745 (2014 г.).
КАС Google ученый
Гавини С.К., Джонс У.К. и Новак С.М. Активация вентромедиального гипоталамического меланокортинового рецептора: регуляция расхода энергии и термогенеза скелетных мышц: VMH и термогенез скелетных мышц. J. Physiol. 594 , 5285–5301 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Мик, Т. Х. и др. Функциональная идентификация нейроцепи, регулирующей уровень глюкозы в крови. Проц. Натл акад. науч. США 113 , E2073–E2082 (2016 г.).
КАС пабмед ПабМед Центральный Статья Google ученый
Борг, В.П., Шервин, Р.С., Воин, М.Дж., Борг, М.А. и Шульман, Г.И. Локальная вентромедиальная глюкозопения гипоталамуса вызывает выброс контррегуляторного гормона. Диабет 44 , 180–184 (1995).
КАС Статья пабмед Google ученый
Furigo, I.C. et al. Гормон роста усиливает восстановление гипогликемии через вентромедиальные гипоталамические нейроны. FASEB J . https://doi.org/10.1096/fj.2015R (2019 г.).
Garfield, A.S. et al. Парабрахиально-гипоталамический холецистокининовый нейроконтур контролирует контррегуляторные реакции на гипогликемию. Сотовый метаб. 20 , 1030–1037 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Sabatini, P.V. et al. tTARGIT AAV опосредуют чувствительные и гибкие манипуляции с популяциями интерсекционных нейронов у мышей. eLife 10 , e66835 (2021).
ПабМед ПабМед Центральный Статья КАС Google ученый
Бурдаков Д. и Гонсалес Дж. А. Физиологические функции нейронов, ингибируемых глюкозой. Acta Physiol. 195 , 71–78 (2009 г.)).
КАС Статья Google ученый
Симадзу, Т. и Минокоши, Ю. Системная регуляция глюкозы чувствительными к глюкозе нейронами в вентромедиальном ядре гипоталамуса (VMH). J. Endocr. соц. 1 , 449–459 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Hirschberg, P.R., Sarkar, P., Teegala, S.B. & Routh, VH. Нейроны, ингибированные глюкозой в вентромедиальном гипоталамусе: роль в гомеостазе глюкозы и энергии? J. Нейроэндокринол. 32 , e12773 (2020).
КАС Статья пабмед Google ученый
Flak, J. N. et al. Ингибированные лептином нейроны PBN усиливают реакцию на гипогликемию при отрицательном энергетическом балансе. Нац. Неврологи. 17 , 1744–1750 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Dunn-Meynell, A. A. et al. Взаимосвязь между уровнями глюкозы в мозге и крови и спонтанным и глюкопривным питанием. J. Neurosci. 29 , 7015–7022 (2009 г.).
КАС пабмед ПабМед Центральный Статья Google ученый
Fioramonti, X., Chrétien, C., Leloup, C. & Pénicaud, L. Недавние достижения в области клеточных и молекулярных механизмов обнаружения глюкозы в гипоталамических нейронах. Фронт. Физиол. 8 , 875 (2017).
ПабМед ПабМед Центральный Статья Google ученый
Hwang, J.J. et al. Притупление повышения уровня глюкозы в мозге при гипергликемии у взрослых с ожирением и СД2. JCI Insight 2 , e95913 (2017).
Центральный пабмед Статья Google ученый
Williams, E.K. et al. Сенсорные нейроны, которые обнаруживают растяжение и питательные вещества в пищеварительной системе. Cell 166 , 209–221 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Донован, С. М. Определение уровня глюкозы в воротной вене. Диабет Нутр. Метаб. 15 , 308–312 (2002).
КАС пабмед Google ученый
Чан, О. и Шервин, Р. С. Существуют ли перекрестные помехи между портальными и гипоталамическими цепями, чувствительными к глюкозе? Диабет 63 , 2617–2619 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Jokiaho, A.J., Donovan, C.M. & Watts, A.G. Скорость падения уровня глюкозы в крови определяет необходимость проецирования катехоламинергических нейронов переднего мозга для симпатоадреналовых реакций самцов крыс. Диабет 63 , 2854–2865 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Campfield, L.A., Smith, F.J., Guisez, Y., Devos, R. & Burn, P. Рекомбинантный OB-белок мыши: свидетельство периферического сигнала, связывающего ожирение и центральные нейронные сети. Наука 269 , 546–549 (1995).
КАС Статья пабмед Google ученый
Schwartz, M.W. et al. Специфичность действия лептина на повышенный уровень глюкозы в крови и экспрессию гена гипоталамического нейропептида Y у ob/ob мышей. Диабет 45 , 531–535 (1996).
КАС Статья пабмед Google ученый
ван ден Хук, А. М. и др. Дефицит лептина сам по себе определяет состав тела и действие инсулина у мышей ob/ob . J. Нейроэндокринол. 20 , 120–127 (2008).
ПабМед Статья КАС Google ученый
Ахима, Р. С. и др. Роль лептина в нейроэндокринной реакции на голодание. Природа 382 , 250–252 (1996).
КАС пабмед Статья Google ученый
Шварц М.В., Сили Р.Дж., Кэмпфилд Л.А., Берн П. и Баскин Д.Г. Идентификация мишеней действия лептина в гипоталамусе крыс. Дж. Клин. Инвестировать. 98 , 1101–1106 (1996).
КАС пабмед ПабМед Центральный Статья Google ученый
Ahrén, B., Månsson, S., Gingerich, R.L. & Havel, PJ. Регуляция лептина в плазме у мышей: влияние возраста, диеты с высоким содержанием жиров и голодания. утра. Дж. Физиол. 273 , R113–R120 (1997 г.).
ПабМед Google ученый
Клеман, К. и др. Мутация в гене рецептора лептина человека вызывает ожирение и дисфункцию гипофиза. Природа 392 , 398–401 (1998).
ПабМед Статья Google ученый
Montague, C. T. et al. Врожденный дефицит лептина связан с тяжелым ранним началом ожирения у людей. Природа 387 , 903–908 (1997).
КАС пабмед Статья Google ученый
Zhang, Y. et al. Позиционное клонирование гена ожирения мыши и его человеческого гомолога. Природа 372 , 425–432 (1994).
КАС пабмед Статья Google ученый
Д’соуза, А. М., Асади, А., Джонсон, Дж. Д., Кови, С. Д. и Киффер, Т. Дж. Дефицит лептина у крыс приводит к гиперинсулинемии и нарушению гомеостаза глюкозы. Эндокринология 155 , 1268–1279 (2014).
ПабМед Статья КАС Google ученый
Pelleymounter, MA et al. Влияние продукта гена ожирения на регуляцию массы тела у мышей ob/ob . Наука 269 , 540–543 (1995).
КАС пабмед Статья Google ученый
Орал, Э. А. и др. Лептино-заместительная терапия липодистрофии. Н. англ. Дж. Мед. 346 , 570–578 (2002).
КАС пабмед Статья Google ученый
Petersen, K. F. et al. Лептин устраняет резистентность к инсулину и стеатоз печени у пациентов с тяжелой липодистрофией. Дж. Клин. Инвестировать. 109 , 1345–1350 (2002).
КАС пабмед ПабМед Центральный Статья Google ученый
Шимомура И., Хаммер Р. Э., Икемото С., Браун М. С. и Гольдштейн Дж. Л. Лептин устраняет резистентность к инсулину и сахарный диабет у мышей с врожденной липодистрофией. Природа 401 , 73–76 (1999).
КАС пабмед Статья Google ученый
Havel, P. J. et al. Заметное и быстрое снижение циркулирующего лептина у крыс со стрептозотоциновым диабетом: реверсирование инсулином. утра. Дж. Физиол. 274 , R1482–R1491 (1998).
КАС пабмед Google ученый
Чинооковонг, Н., Ван, Дж. Л. и Ши, З. К. Лептин восстанавливает эугликемию и нормализует обмен глюкозы при инсулинодефицитном диабете у крыс. Диабет 48 , 1487–1492 (1999).
КАС пабмед Статья Google ученый
German, J.P. et al. Дефицит лептина вызывает резистентность к инсулину, вызванную неконтролируемым диабетом. Диабет 59 , 1626–1634 (2010).
КАС пабмед ПабМед Центральный Статья Google ученый
Hidaka, S. et al. Хроническая центральная инфузия лептина восстанавливает гипергликемию независимо от приема пищи и уровня инсулина у крыс с диабетом, индуцированным стрептозотоцином. FASEB J. 16 , 509–518 (2002).
КАС пабмед Статья Google ученый
Fujikawa, T. et al. Лептин задействует гипоталамическую нейросхему, чтобы обеспечить выживание в отсутствие инсулина. Сотовый метаб. 18 , 431–444 (2013).
КАС пабмед ПабМед Центральный Статья Google ученый
Fujikawa, T., Chuang, J.-C., Sakata, I., Ramadori, G. & Coppari, R. Терапия лептином улучшает инсулин-дефицитный диабет 1 типа с помощью механизмов, зависящих от ЦНС, у мышей. Проц. Натл акад. науч. США 107 , 17391–17396 (2010 г.).
КАС пабмед ПабМед Центральный Статья Google ученый
Мортон, Г. Дж., Мик, Т. Х., Матсен, М. Э. и Шварц, М. В. Доказательства против подавления гипоталамо-гипофизарно-надпочечниковой оси при антидиабетическом действии лептина. Дж. Клин. Инвестировать. 125 , 4587–4591 (2015).
ПабМед ПабМед Центральный Статья Google ученый
Perry, R. J. et al. Лептин излечивает диабет путем подавления гипоталамо-гипофизарно-надпочечниковой оси. Нац. Мед. 20 , 759–763 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Аликс, П. М., Гебре-Эгзиабхер, Ф. и Сулаж, К. О. Лептин как уремический токсин: пагубная роль лептина при хроническом заболевании почек. Биохимия 105 , 12–21 (2014).
КАС Статья пабмед Google ученый
Mittendorfer, B. et al. Лечение рекомбинантным человеческим лептином не улучшает действие инсулина у пациентов с ожирением и диабетом 2 типа. Диабет 60 , 1474–1477 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Мун, Х.-С. и другие. Эффективность метрелептина у пациентов с ожирением и диабетом 2 типа: клеточные и молекулярные пути, лежащие в основе толерантности к лептину. Диабет 60 , 1647–1656 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Ханаи, К. и др. Центральное действие кислого фактора роста фибробластов в регуляции питания. утра. Дж. Физиол. 256 , R217–R223 (1989).
КАС пабмед Google ученый
Оомура Ю. и др. Новый глюкозосенсор мозга и его физиологическое значение. утра. Дж. Клин. Нутр. 55 , 278С–282С (1992).
КАС Статья пабмед Google ученый
Сасаки, К. и др. Влияние факторов роста фибробластов и родственных пептидов на потребление пищи крысами. Физиол. Поведение 56 , 211–218 (1994).
КАС Статья пабмед Google ученый
Suh, J.M. et al. Эндокринизация FGF1 продуцирует неоморфный и мощный сенсибилизатор инсулина. Природа 513 , 436–439 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Теннант, К. Г., Линдсли, С. Р., Киригити, М. А., Тру, К. и Киевит, П. Центральное и периферическое введение фактора роста фибробластов 1 улучшает секрецию инсулина островками поджелудочной железы на моделях мышей с диабетом. Диабет 68 , 1462–1472 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Скарлетт, Дж. М. и др. Центральная инъекция фактора роста фибробластов 1 вызывает устойчивую ремиссию диабетической гипергликемии у грызунов. Нац. Мед. 22 , 800–806 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Браун, Дж. М. и др. Срединное возвышение дугообразного ядра гипоталамуса является мишенью для устойчивой ремиссии диабета, вызванной фактором роста фибробластов 1. Диабет 68 , 1054–1061 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Скарлетт, Дж. М. и др. Периферические механизмы, опосредующие устойчивое антидиабетическое действие FGF1 в головном мозге. Диабет 68 , 654–664 (2019).
КАС Статья пабмед Google ученый
Коппари, Р. и др. Аркуатное ядро гипоталамуса: ключевое место для опосредования эффектов лептина на гомеостаз глюкозы и двигательную активность. Метаб. клеток 1 , 63–72 (2005).
КАС Статья пабмед Google ученый
Berglund, E.D. et al. Прямое действие лептина на нейроны РОМС регулирует гомеостаз глюкозы и чувствительность печени к инсулину у мышей. Дж. Клин. Инвестировать. 122 , 1000–1009 (2012).
КАС пабмед ПабМед Центральный Статья Google ученый
Goncalves, G. H.M., Li, W., Garcia, A.V.C.-G., Figueiredo, M.S. & Bjørbæk, C. Пептидные нейроны гипоталамуса, связанные с агути, и центральная меланокортиновая система являются важнейшими медиаторами противодиабетического действия лептина. Cell Rep. 7 , 1093–1103 (2014).
ПабМед ПабМед Центральный Статья КАС Google ученый
Huo, L. et al. Лептин-зависимый контроль баланса глюкозы и двигательной активности нейронами РОМС. Метаб. клеток 9 , 537–547 (2009).
КАС пабмед ПабМед Центральный Статья Google ученый
Бентсен, М. А. и др. Транскриптомный анализ связывает различные типы клеток гипоталамуса с устойчивой ремиссией диабета, вызванной фактором роста фибробластов 1. Нац. коммун. 11 , 4458 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Аонге, К. М. и др. Сборка гипоталамической перинейрональной сети необходима для устойчивой ремиссии диабета, вызванного фактором роста фибробластов 1 у крыс. Нац. Метаб. 2 , 1025–1033 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Кларет, М. и др. Делеция Lkb1 в проопиомеланокортиновых нейронах нарушает периферический гомеостаз глюкозы у мышей. Диабет 60 , 735–745 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Smith, M. A. et al. Рибосомы S6K1 в нейронах POMC и AgRP регулируют гомеостаз глюкозы, но не пищевое поведение у мышей. Cell Rep. 11 , 335–343 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Чабра, К. Х. и др. Дефицит гипоталамической ПОМС улучшает толерантность к глюкозе, несмотря на резистентность к инсулину, за счет увеличения глюкозурии. Диабет 65 , 660–672 (2016).
КАС Статья пабмед Google ученый
Чабра, К. Х. и др. Снижение активности почечных симпатических нервов способствует повышению глюкозурии и улучшению толерантности к глюкозе в гипоталамус-специфических Pomc 9Нокаутные мыши 0005. Мол. Метаб. 6 , 1274–1285 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Shin, A.C. et al. Передача сигналов инсулинового рецептора в POMC, но не в AgRP, нейронах контролирует действие инсулина в жировой ткани. Диабет 66 , 1560–1571 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Рен Х. и др. Gpr17-мишень FoxO1 активирует нейроны AgRP для регулирования потребления пищи. Cell 149 , 1314–1326 (2012).
КАС пабмед ПабМед Центральный Статья Google ученый
Сюй, А. В. и др. PI3K объединяет действие инсулина и лептина на нейроны гипоталамуса. Дж. Клин. Инвестировать. 115 , 951–958 (2005).
КАС пабмед ПабМед Центральный Статья Google ученый
Könner, A.C. et al. Действие инсулина в нейронах, экспрессирующих AgRP, необходимо для подавления продукции глюкозы в печени. Сотовый метаб. 5 , 438–449 (2007).
Артикул КАС пабмед Google ученый
Aponte, Y., Atasoy, D. & Sternson, S.M. Нейронов AGRP достаточно для быстрой организации пищевого поведения без обучения. Нац. Неврологи. 14 , 351–355 (2011).
КАС Статья пабмед Google ученый
Krashes, M.J. et al. Быстрая обратимая активация нейронов AgRP управляет пищевым поведением у мышей. Дж. Клин. Инвестировать. 121 , 1424–1428 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Engström Ruud, L., Pereira, M.M.A., de Solis, A.J., Fenselau, H. & Brüning, JC. NPY опосредует быстрое питание и регуляторные функции метаболизма глюкозы нейронов AgRP. Нац. коммун. 11 , 442 (2020).
ПабМед ПабМед Центральный Статья КАС Google ученый
ван де Валл, Э. и др. Коллективные и индивидуальные функции лептиновых рецепторов модулируют нейроны, контролирующие метаболизм и прием пищи. Эндокринология 149 , 1773–1785 (2008).
Артикул КАС пабмед Google ученый
Deem, J.D. et al. Индуцированная холодом гиперфагия требует активации нейронов AgRP у мышей. eLife 9 , e58764 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Лю, Т. и др. Активация нейронов AgRP натощак требует рецепторов NMDA и включает спиногенез и повышенный возбуждающий тонус. Нейрон 73 , 511–522 (2012).
ПабМед ПабМед Центральный Статья КАС Google ученый
Mizuno, T.M. et al. Гипоталамическая мРНК проопиомеланокортина снижается натощак у мышей ob/ob и db/db , но стимулируется лептином. Диабет 47 , 294–297 (1998).
КАС Статья пабмед Google ученый
Takahashi, K. A. & Cone, R. D. Голодание вызывает значительное, лептин-зависимое увеличение частоты собственного потенциала действия орексигенного нейропептида дугообразного ядра Y/агути-родственных белковых нейронов. Эндокринология 146 , 1043–1047 (2005).
КАС Статья пабмед Google ученый
Колачински, Дж. В., Оганнесян, Дж. П., Консидайн, Р. В., Марко, К. С. и Каро, Дж. Ф. Реакция лептина на кратковременное и длительное переедание у людей. Дж. Клин. Эндокринол. Метаб. 81 , 4162–4165 (1996).
КАС пабмед Google ученый
Колао, А. и др. Лечение гипергликемии у пациентов с болезнью Кушинга, получающих пасиреотид: рекомендации медицинского эксперта. Гипофиз 17 , 180–186 (2014).
КАС Статья пабмед Google ученый
Hsia, D. S., Grove, O. & Cefalu, W. T. Обновленная информация об ингибиторах натрий-глюкозного котранспортера-2 для лечения сахарного диабета. Курс. мнение Эндокринол. Диабет Ожирение. 24 , 73–79 (2017).
КАС пабмед ПабМед Центральный Google ученый
Скарони, К., Зилио, М., Фоти, М. и Боскаро, М. Нарушения метаболизма глюкозы при синдроме Кушинга: от молекулярной основы до клинического лечения. Эндокр. 38 , 189–219 (2017).
Артикул пабмед Google ученый
Ши, Дж. и др. Обзор: черепно-мозговая травма и гипергликемия, потенциально модифицируемый фактор риска. Oncotarget 7 , 71052–71061 (2016).
ПабМед ПабМед Центральный Статья Google ученый
Farooqi, I. S. et al. Частичный дефицит лептина и ожирение человека. Природа 414 , 34–35 (2001).
КАС Статья пабмед Google ученый
Флак, Дж. Н. и Майерс, М. Г. Миниобзор: Механизмы действия лептина на ЦНС. Мол. Эндокринол. 30 , 3–12 (2016).
КАС Статья пабмед Google ученый
Bal, N.C. et al. Как бурая жировая ткань, так и процессы термогенеза скелетных мышц активируются во время легкой и тяжелой адаптации мышей к холоду. Дж. Биол. хим. 292 , 16616–16625 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Моррисон С.Ф., Мэдден С.Дж. и Тупоун Д. Центральная нервная регуляция термогенеза бурой жировой ткани и расхода энергии. Сотовый метаб. 19 , 741–756 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Carey, M. et al. Центральные K-АТФ-каналы модулируют эффективность глюкозы у людей и грызунов. Диабет 69 , 1140–1148 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Тимпер К. и Брюнинг Дж. К. Гипоталамические цепи, регулирующие аппетит и энергетический гомеостаз: пути к ожирению. Дис. Модель. мех. 10 , 679–689 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Барретт П., Мерсер Дж. Г. и Морган П. Дж. Доклинические модели исследования ожирения. Дис. Модель. мех. 9 , 1245–1255 (2016).
ПабМед ПабМед Центральный Статья Google ученый
Дим, Дж. Д., Мута, К., Скарлетт, Дж. М., Мортон, Г. Дж. и Шварц, М. В. Что мы должны думать о роли мозга в гомеостазе глюкозы и диабете? Диабет 66 , 1758–1765 (2017).
КАС пабмед ПабМед Центральный Статья Google ученый
Сюй, Дж. и др. Генетическая идентификация нейронных цепей лептина в гомеостазе энергии и глюкозы. Природа 556 , 505–509 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Halban, P. A. et al. Отказ β-клеток при диабете 2 типа: предполагаемые механизмы и перспективы профилактики и лечения. Diabetes Care 37 , 1751–1758 (2014).
ПабМед ПабМед Центральный Статья Google ученый
Hermann, G.M. et al. Неонатальный догоняющий рост увеличивает восприимчивость к диабету, но улучшает поведенческие и сердечно-сосудистые исходы у самцов мышей с низкой массой тела при рождении. Педиатр. Рез 66 , 53–58 (2009).
ПабМед ПабМед Центральный Статья Google ученый
Perng, W., Oken, E. & Dabelea, D. Переедание в процессе развития, ожирение и диабет 2 типа у потомства. Diabetologia 62 , 1779–1788 (2019).
Артикул пабмед Google ученый
Шеллонг, К. и др. Специфические для пола эпигенетические изменения гипоталамической системы Agrp-Pomc не объясняют «диабетизм» у потомства перекормленных матерей крыс с высоким содержанием жиров (HFD). Дж. Нутр. Биохим. 75 , 108257 (2020).
КАС Статья пабмед Google ученый
de Kloet, A.D. & Herman, J.P. Связи между жиром и мозгом: глюкокортикоидный контроль адипоцитов над стрессом и метаболизмом. Фронт. Нейроэндокринол. 48 , 50–57 (2018).
Артикул КАС пабмед Google ученый
Магомедова Л. и Камминс С. Л. Глюкокортикоиды и метаболический контроль. Справочник. Эксп. Фармакол. 233 , 73–93 (2016).
КАС Статья пабмед Google ученый
Warner, A. & Mittag, J. Гормон щитовидной железы и центральный контроль гомеостаза. Дж. Мол. Эндокринол. 49 , Р29–Р35 (2012 г.).
КАС пабмед Статья Google ученый
Каррагета Д. Ф., Оливейра П. Ф., Алвес М. Г. и Монтейро М. П. Ожирение и мужской гипогонадизм: рассказы о порочном круге. Обес. Ред. 20 , 1148–1158 (2019).
ПабМед Google ученый
López, M. & Tena-Sempere, M. Влияние эстрадиола на гипоталамический термогенез AMPK и BAT: путь к лечению ожирения? Фармакол. тер. 178 , 109–122 (2017).
Артикул КАС пабмед Google ученый
Мове-Джарвис Ф., Клегг Д. Дж. и Хевенер А. Л. Роль эстрогенов в контроле энергетического баланса и гомеостаза глюкозы. Эндокр. 34 , 309–338 (2013).
КАС пабмед ПабМед Центральный Статья Google ученый
Койман С., ван ден Хеувел Дж. К. и Ренсен П. К. Н. Нейронный контроль активности бурого жира. Тенденции Эндокринол. Метаб. 26 , 657–668 (2015).
КАС Статья пабмед Google ученый
Бартнесс Т.Дж., Воган С.Х. и Сонг С.К. Симпатическая и сенсорная иннервация бурой жировой ткани. Междунар. Дж. Обес. 34 , С36–С42 (2010).
Артикул Google ученый
McNeill, B. T., Morton, N. M. & Stimson, R. H. Утилизация субстрата бурой жировой тканью: что горячо, а что нет? Фронт. Эндокринол. 11 , 571659 (2020).
Артикул Google ученый
Моррисон С. Ф. и Накамура К. Центральные механизмы терморегуляции. год. Преподобный Физиол. 81 , 285–308 (2019).
КАС Статья пабмед Google ученый
Росси, М. А. и Стубер, Г. Д. Перекрывающиеся мозговые цепи для гомеостатического и гедонистического питания. Сотовый метаб. 27 , 42–56 (2018).
КАС Статья пабмед Google ученый
Конус, Р. Д. Анатомия и регуляция центральной меланокортиновой системы. Нац. Неврологи. 8 , 571–578 (2005).
КАС Статья пабмед Google ученый
Саттон А.К., Майерс М.Г. и Олсон Д.П. Роль цепей PVH в действии лептина и энергетическом балансе. Annu Rev. Physiol. 78 , 207–221 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Фрай, М. и Фергюсон, А. В. Органы чувств, окружающие желудочек: мозговые мишени для циркулирующих сигналов, контролирующих пищевое поведение. Физиол. Поведение 91 , 413–423 (2007).
КАС Статья пабмед Google ученый
Ollmann, M.M. et al. Антагонизм центральных рецепторов меланокортина in vitro и in vivo с помощью белка, родственного агути. Наука 278 , 135–138 (1997).
КАС пабмед Статья Google ученый
Schwartz, M.W. et al. Лептин увеличивает экспрессию мРНК гипоталамуса проопиомеланокортина в ростральном дугообразном ядре. Диабет 46 , 2119–2123 (1997).
КАС пабмед Статья Google ученый
Zhan, C. et al. Острое и длительное подавление пищевого поведения нейронами РОМС в стволе мозга и гипоталамусе соответственно. Дж. Неврологи. 33 , 3624–3632 (2013).
КАС пабмед ПабМед Центральный Статья Google ученый
Baskin, D.G., Breininger, J.F. & Schwartz, M.W. мРНК рецептора лептина идентифицирует субпопуляцию нейронов нейропептида Y, активируемых натощак в гипоталамусе крысы. Диабет 48 , 828–833 (1999).
КАС пабмед Статья Google ученый
Эриксон Дж. К., Холлопетер Г. и Палмитер Р. Д. Ослабление синдрома ожирения у мышей ob/ob путем потери нейропептида Y. Science 274 , 1704–1707 (1996).
КАС пабмед Статья Google ученый
Winkler, Z. et al. Активируемая гипогликемией гипоталамическая микроглия ослабляет контррегуляторные реакции глюкозы. Науч. 9 , 6224 (2019).
ПабМед ПабМед Центральный Статья КАС Google ученый
Luquet, S. Нейроны NPY/AgRP необходимы для питания взрослых мышей, но могут быть удалены у новорожденных. Наука 310 , 683–685 (2005).
КАС пабмед Статья Google ученый
Gropp, E. et al. Нейроны, экспрессирующие родственные агути пептиды, обязательны для питания. Нац. Неврологи. 8 , 1289–1291 (2005).
КАС пабмед Статья Google ученый
Zhang, X. & van den Pol, A. N. Дофамин/тирозингидроксилаза нейронов дугообразного ядра гипоталамуса высвобождает ГАМК, взаимодействует с дофаминергическими и другими дугообразными нейронами и реагирует на динорфин, мет-энкефалин и окситоцин. J. Neurosci. 35 , 14966–14982 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Zhang, X. & van den Pol, A.N. Нейроны тирозингидроксилазы дугообразного ядра гипоталамуса играют орексигенную роль в энергетическом гомеостазе. Нац. Неврологи. 19 , 1341–1347 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Атасой, Д., Бетли, Дж. Н., Су, Х. Х. и Стернсон, С. М. Деконструкция нейронной цепи для голода. Природа 488 , 172–177 (2012).
КАС пабмед ПабМед Центральный Статья Google ученый
Garfield, A.S. et al. Нейронная основа аппетита, регулируемого рецептором меланокортина-4. Нац. Неврологи. 18 , 863–871 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Li, M. M. et al. Паравентрикулярный гипоталамус регулирует насыщение и предотвращает ожирение посредством двух генетически различных цепей. Нейрон 102 , 653–667 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Leinninger, G.M. et al. Действие лептина через нейроны нейротензина контролирует орексин, мезолимбическую дофаминовую систему и энергетический баланс. Сотовый метаб. 14 , 313–323 (2011).
КАС пабмед ПабМед Центральный Статья Google ученый
Hawke, Z. et al. Нейроны PACAP в вентромедиальном ядре гипоталамуса являются мишенями центральной передачи сигналов лептина. J. Neurosci. 29 , 14828–14835 (2009).
КАС пабмед ПабМед Центральный Статья Google ученый
Паттерсон, С. М., Лешан, Р. Л., Джонс, Дж. К. и Майерс, М. Г. Молекулярное картирование областей мозга мыши, иннервируемых клетками, экспрессирующими рецептор лептина. Мозг Res. 1378 , 18–28 (2011).
КАС пабмед Статья Google ученый
Резай-Заде, К. и др. Нейроны рецепторов лептина в дорсомедиальном гипоталамусе являются ключевыми регуляторами расхода энергии и массы тела, но не потребления пищи. Мол. Метаб. 3 , 681–693 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Chou, T.C. et al. Критическая роль дорсомедиального ядра гипоталамуса в широком диапазоне поведенческих циркадианных ритмов. J. Neurosci. 23 , 10691–10702 (2003).
КАС пабмед ПабМед Центральный Статья Google ученый
Grill, H. J. & Hayes, M. R. Нейроны заднего мозга как важный узел нейроанатомически распределенного контроля энергетического баланса. Сотовый метаб. 16 , 296–309 (2012).
КАС пабмед ПабМед Центральный Статья Google ученый
Zhang, C. et al. Типы клеток Area postrema, которые опосредуют поведение, связанное с тошнотой. Нейрон https://doi. org/10.1016/j.neuron.2020.11.010 (2020).
Людвиг, М. К. и др. Генетическая карта дорсального вагусного комплекса мыши и его роль в ожирении. Нац. Метаб . https://doi.org/10.1038/s42255-021-00363-1 (2021 г.).
Sabatini, P.V. et al. Нейроны, экспрессирующие GFRAL, подавляют прием пищи через пути отвращения. Проц. Натл акад. науч. США 118 , e2021357118 (2021).
ПабМед ПабМед Центральный Статья КАС Google ученый
Бай, Л. и др. Генетическая идентификация сенсорных нейронов блуждающего нерва, контролирующих питание. Сотовый 179 , 1129–1143 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Кельберер, М. М., Рупрехт, Л. Э., Лю, В. В., Венг, П. и Бохоркес, Д. В. Клетки нейроподов: новая биология сенсорной трансдукции кишечника и мозга. Annu Rev. Neurosci. 43 , 337–353 (2020).
КАС Статья пабмед ПабМед Центральный Google ученый
Huo, L., Maeng, L., Bjørbaek, C. & Grill, H.J. Лептин и контроль потребления пищи: нейроны в ядре солитарного тракта активируются как растяжением желудка, так и лептином. Эндокринология 148 , 2189–2197 (2007).
КАС Статья пабмед Google ученый
Huo, L., Gamber, KM, Grill, HJ & Bjørbaek, C. Дивергентная передача сигналов лептина в проглюкагоновых нейронах ядра солитарного тракта у мышей и крыс. Эндокринология 149 , 492–497 (2008).
КАС Статья пабмед Google ученый
Цай, В. В.-В. и другие. GDF15 опосредует устойчивость к ожирению посредством воздействия на нейроны GFRAL в AP/NTS заднего мозга. Международный Дж. Обес. 43 , 2370–2380 (2019).
Артикул КАС Google ученый
Alhadeff, A.L. et al. Эндогенная передача сигналов рецептора глюкагоноподобного пептида-1 в ядре одиночного пути необходима для контроля потребления пищи. Нейропсихофармакология 42 , 1471–1479 (2017).
КАС Статья пабмед Google ученый
Hayes, M. R. et al. Эндогенная передача сигналов лептина в хвостовом ядре tractus solitarius и area postrema необходима для регуляции энергетического баланса. Сотовый метаб. 23 , 744 (2016).
КАС Статья пабмед Google ученый
Cheng, W. et al. Нейроны рецептора кальцитонина в ядре солитарного тракта мыши контролируют энергетический баланс посредством неаверсивного подавления кормления. Метаб. клеток 31 , 301–312 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Cheng, W. et al. Нейроны ядра одиночного пути, экспрессирующие рецептор лептина, подавляют потребление пищи независимо от GLP1 у мышей. JCI Insight 5 , e134359 (2020).
Центральный пабмед Статья Google ученый
Wadden, T. A. et al. Влияние семаглутида подкожно по сравнению с плацебо в качестве дополнения к интенсивной поведенческой терапии на массу тела у взрослых с избыточной массой тела или ожирением: рандомизированное клиническое исследование STEP 3. JAMA https://doi.org/10.1001/jama.2021.1831 (2021).
Палмитер, Р. Д. Парабрахиальное ядро: нейроны CGRP функционируют как общий сигнал тревоги. Trends Neurosci. 41 , 280–293 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Чен, Дж. Ю., Кампос, К. А., Джарви, Б. С. и Палмитер, Р. Д. Парабрахиальные нейроны CGRP создают и поддерживают неприятные вкусовые воспоминания. Нейрон 100 , 891–899 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Кампос, К. А., Боуэн, А. Дж., Шварц, М. В. и Палмитер, Р. Д. Парабрахиальные нейроны CGRP контролируют прекращение приема пищи. Сотовый метаб. 23 , 811–820 (2016).
КАС пабмед ПабМед Центральный Статья Google ученый
Д’Агостино, Г. и др. Аппетит контролируется холецистокининовым ядром солитарного пути к нейроцепи гипоталамуса. eLife 5 , e12225 (2016).
ПабМед ПабМед Центральный Статья КАС Google ученый
Ву, К., Бойл, М.П. и Пальмитер, Р.Д. Потеря ГАМКергической передачи сигналов нейронами AgRP к парабрахиальному ядру приводит к голоданию. Cell 137 , 1225–1234 (2009).
ПабМед ПабМед Центральный Статья Google ученый
Шах, Б.П. и др. Экспрессирующие MC4R глутаматергические нейроны в паравентрикулярном гипоталамусе регулируют питание и синаптически связаны с парабрахиальным ядром. Проц. Натл акад. науч. США 111 , 13193–13198 (2014).
КАС пабмед ПабМед Центральный Статья Google ученый
Li, A.-J., Wang, Q. & Ritter, S. Селективная фармакогенетическая активация подгрупп катехоламинов в вентролатеральном мозговом веществе вызывает ключевые глюкорегуляторные реакции. Эндокринология 159 , 341–355 (2018).
КАС Статья пабмед Google ученый
Hudson, B. & Ritter, S. Катехоламиновые нейроны заднего мозга опосредуют завершающие реакции на глюкопривацию. Физиол. Поведение 82 , 241–250 (2004).
КАС Статья пабмед Google ученый
Cryer, P. E. Контррегуляция глюкозы: профилактика и коррекция гипогликемии у людей. утра. Дж. Физиол. 264 , E149–E155 (1993).
КАС пабмед Google ученый
Li, A.-J., Wang, Q., Elsarelli, M.M., Brown, R.L. & Ritter, S. Катехоламиновые нейроны заднего мозга активируют орексиновые нейроны во время системной глюкопривации у самцов крыс. Эндокринология 156 , 2807–2820 (2015).
КАС пабмед ПабМед Центральный Статья Google ученый
Аклан, И. и др. Катехоламиновые нейроны NTS опосредуют гипогликемический голод через медиальные пути питания гипоталамуса. Метаб. клеток 31 , 313–326 (2020).
КАС Статья пабмед Google ученый
Хенш, Т. К. Пластичность критического периода в локальных корковых цепях. Нац. Преподобный Нейроски. 6 , 877–888 (2005).
КАС Статья пабмед Google ученый
Bouret, S.G. Трофическое действие лептина на нейроны гипоталамуса, регулирующие питание. Наука 304 , 108–110 (2004).
КАС Статья пабмед Google ученый
Камитакахара, А., Буйе, К., Ван, К.-Х. & Симерли, Р. Критический период трофического действия лептина на нейроны AgRP в аркуатном ядре гипоталамуса. Дж. Комп. Нейрол. 526 , 133–145 (2018).
КАС пабмед Статья Google ученый
Буре, С. Г., Дрейпер, С. Дж. и Симерли, Р. Б. Формирование проекционных путей от аркуатного ядра гипоталамуса к гипоталамическим областям, участвующим в нервном контроле пищевого поведения у мышей. J. Neurosci. 24 , 2797–2805 (2004).
КАС пабмед ПабМед Центральный Статья Google ученый
Rupp, A.C. et al. Специфические субпопуляции гипоталамических нейронов, экспрессирующих рецептор лептина, опосредуют эффекты ранней делеции рецептора лептина в развитии на энергетический баланс. Мол. Метаб. 14 , 130–138 (2018).
КАС пабмед ПабМед Центральный Статья Google ученый
Ахима, Р. С., Прабакаран, Д. и Флиер, Дж. С. Постнатальный всплеск лептина и регуляция циркадного ритма лептина при кормлении. Влияние на энергетический гомеостаз и нейроэндокринную функцию. Дж. Клин. Инвестировать. 101 , 1020–1027 (1998).
КАС пабмед ПабМед Центральный Статья Google ученый
Биддингер, Дж. Э., Лазаренко, Р. М., Скотт, М. М. и Симерли, Р. Лептин подавляет развитие входов ГПП-1 в паравентрикулярное ядро гипоталамуса. eLife 9 , e59857 (2020).
КАС пабмед ПабМед Центральный Статья Google ученый
Ринаман, Л. Постнатальное развитие входов катехоламинов в паравентрикулярное ядро гипоталамуса у крыс. Дж. Комп. Нейрол. 438 , 411–422 (2001).
КАС пабмед Статья Google ученый
Мирзаде З. и др. Формирование перинейрональной сети в критический период созревания нейронов в дугообразном ядре гипоталамуса. Нац. Метаб. 1 , 212–221 (2019).
КАС пабмед ПабМед Центральный Статья Google ученый
Карулли, Д. и др. Животные, у которых отсутствует link белок, имеют ослабленные перинейрональные сети и постоянную пластичность. Мозг 133 , 2331–2347 (2010).
Артикул пабмед Google ученый
Пиццоруссо, Т. Реактивация пластичности глазного доминирования во взрослой зрительной коре. Наука 298 , 1248–1251 (2002).
КАС Статья пабмед Google ученый
Гоголла, Н., Карони, П., Люти, А. и Херри, К. Перинейронные сети защищают воспоминания о страхе от стирания. Наука 325 , 1258–1261 (2009).
КАС пабмед Статья Google ученый
Tamashiro, K.L.K., Terrillion, C.E., Hyun, J., Koenig, J.I. & Moran, T.H. Пренатальный стресс или диета с высоким содержанием жиров повышают восприимчивость к ожирению, вызванному диетой, у потомства крыс. Диабет 58 , 1116–1125 (2009).
КАС пабмед ПабМед Центральный Статья Google ученый
Главас М.М. и др. Раннее переедание приводит к ранней резистентности дугообразного лептина и повышенной чувствительности к диете с высоким содержанием жиров. Эндокринология 151 , 1598–1610 (2010).
КАС пабмед ПабМед Центральный Статья Google ученый
1.3: Гомеостаз и системы контроля
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 11099
цели обучения
- Определение гомеостаза
- Дайте определение системе управления и опишите ее компоненты
- Дайте определение отрицательной обратной связи и приведите один пример с использованием температуры тела
- Дайте определение положительным отзывам и приведите один пример; также сравните и сопоставьте положительные и отрицательные отзывы с точки зрения взаимосвязи между ответом и результатом
Гомеостаз (гомео- = «подобный, похожий, такого же рода»; stasis = «стоять на месте») означает поддерживать функции организма в определенных приемлемых пределах, приспосабливаясь к внутренним и внешним изменениям. Температура, концентрация питательных веществ, кислотность, вода, натрий, кальций, кислород, а также кровяное давление, частота сердечных сокращений и частота дыхания — вот некоторые из внутренних переменных организма, которые должны оставаться в определенном диапазоне. Когда организму не удается поддерживать внутренние параметры тела в определенном диапазоне, нормальная функция нарушается, что может привести к заболеванию или заболеванию.
проверка понятий, терминов и фактов
Учебные вопросы Напишите свой ответ в виде предложения (не используйте в ответе расплывчатые слова)
1. Какие внутренние переменные тела должны оставаться в определенном диапазоне?
2. Что происходит, когда внутренние переменные тела выходят за пределы определенного диапазона?
3. Что такое гомеостаз?
Все, что должно поддерживаться в организме в пределах нормы, должно иметь систему управления . Система управления состоит из четырех компонентов:
Стимул или физиологическая переменная, которая изменяет , является предметом регулирования. Переменная в широком смысле — это значение, которое варьируется или меняется. Двумя примерами изменяющихся переменных являются температура тела и уровень глюкозы в крови. Все, что можно измерить и изменить, является переменной.
Сенсор или сенсорный рецептор — это клетка, ткань или орган, воспринимающие изменение стимула или физиологической переменной. Например, окончания чувствительных нервных клеток в коже ощущают повышение температуры тела, а специализированные клетки поджелудочной железы ощущают падение уровня глюкозы в крови. Сенсорный рецептор или датчик обеспечивает вход в центр управления.
Центр управления — это структура тела, которая определяет нормальный диапазон переменной или заданное значение . Например, область мозга, называемая гипоталамусом, определяет уставку температуры тела (около 37 °C, или 98,6 °F), а специализированные клетки поджелудочной железы определяют уставку глюкозы в крови (около 70–100 мг/дл). ). Для поддержания гомеостаза центр управления реагирует на изменения стимула, полученного от сенсора, посылая сигналы эффекторам.
Эффектор — это клетка, ткань или орган, которые реагируют на сигналы из центра управления, тем самым обеспечивая реакцию на стимул (физиологическая переменная, которая изменилась) для поддержания гомеостаза. Например, потовые железы (эффекторы) по всему телу выделяют пот для снижения температуры тела; а клетки печени (эффекторы) выделяют глюкозу для повышения уровня глюкозы в крови. См. четыре компонента системы управления на рисунке \(\PageIndex{1}\) ниже.
Рисунок \(\PageIndex{1}\) Компоненты системы управления: изменяющийся стимул или физиологическая переменная, датчик или сенсорный рецептор, центр управления и эффектор
Проверка понятий, терминов и фактов
Учебные вопросы Напишите свой ответ в виде предложения (не используйте в ответе нестандартные слова)
1. Какие четыре компонента входят в систему управления?
2. Дайте определение стимулу на двух примерах
3. Что такое сенсор (или сенсорный рецептор)?
4. Что такое центр управления?
5. Что такое уставка?
6. Что такое эффектор?
Большинство систем управления поддерживают гомеостаз с помощью процесса, называемого отрицательной обратной связью . Отрицательная обратная связь предотвращает выход физиологической переменной или функции организма за пределы нормального диапазона. Он делает это, обращая изменение физиологической переменной (стимула) после превышения нормального диапазона. Компонентами отрицательной обратной связи являются датчик (или сенсорный рецептор), центр управления (где находится заданная точка) и эффектор. См. рисунок \(\PageIndex{2}\) ниже.
Рисунок \(\PageIndex{2}\) В петле отрицательной обратной связи стимул — отклонение от заданного значения — сопротивляется физиологическому процессу, возвращающему организм к гомеостазу. (а) Петля отрицательной обратной связи состоит из четырех основных частей. (б) Температура тела регулируется отрицательной обратной связью.
проверки понятий, терминов и фактов
Учебные вопросы Напишите свой ответ в виде предложения (не используйте неразборчивые слова)
1. Что такое негативная обратная связь?
Положительная обратная связь — это механизм, который усиливает изменение
физиологического состояния организма, а не обращает его вспять (как это делает механизм отрицательной обратной связи). Отклонение от нормального диапазона приводит к большим изменениям, и система отдаляется от нормального диапазона. Положительная обратная связь в организме нормальна только тогда, когда есть определенная конечная точка.
Роды — это пример петли положительной обратной связи, которая является нормальной, но активируется только при необходимости. Первые схватки (стимул) подталкивают ребенка к шейке (самой нижней части матки). Шейка матки содержит чувствительные к растяжению клетки (сенсоры), которые отслеживают степень растяжения. Эти нервные клетки посылают сообщения в мозг (центр управления), который, в свою очередь, заставляет гипофиз в основании мозга выделять гормон окситоцин в кровоток. Окситоцин вызывает более сильные сокращения гладких мышц (эффекторов) в матке, продвигая ребенка дальше по родовым путям. Это вызывает еще большее растяжение шейки матки. Цикл растяжения, выброса окситоцина и все более сильных сокращений прекращается только с рождением ребенка. В этот момент растяжение шейки матки прекращается, прекращая выброс окситоцина. Конец 9Результатом 2344 в цикле положительной обратной связи является достижение конечной точки (доставка), а не достижение заданной точки , как при отрицательной обратной связи. См. рисунок \(\PageIndex{3}\) ниже в качестве примера и таблицу \(\PageIndex{1}\) для сравнения отрицательных и положительных отзывов.
Рисунок \(\PageIndex{3}\) Нормальные роды управляются положительной обратной связью. Петля положительной обратной связи приводит к изменению состояния организма, а не к возврату к заданной точке.
| Отрицательный отзыв | Положительный отзыв | |
|---|---|---|
| Пример | Регуляция температуры тела или уровня глюкозы в крови | Нормальные роды |
| Ответ | Обращает вспять изменение физиологического состояния | Усиливает изменение физиологического состояния |
| Результат | Возврат к заданному значению | Достичь конечной точки |
| Комбинезон | Обеспечение стабильности | Ускоряет процесс до завершения |
Проверка понятий, терминов и фактов. 0008
1. Что такое положительный отзыв?
2. Чем отличаются отрицательные и положительные отзывы (Сравните ответ, результат и общий результат)
- Наверх
- Была ли эта статья полезной?
- Тип изделия
- Раздел или страница
- Лицензия
- CC BY-NC-SA
- Показать оглавление
- нет
- Теги
- гомеостаз
Что означает гомеостаз?
Написано Никлией Харрис-Рэй
Медицинская экспертиза Пунам Сачдев 28 апреля 2022 г.
В этой статье
- Дайте определение гомеостазу
- Пример гомеостаза
- Как работает регуляция с обратной связью?
- Почему важен гомеостаз?
- Как организм поддерживает гомеостаз?
- Где еще применяется гомеостаз?
Любой процесс саморегуляции, используемый биологической системой для поддержания стабильности при постоянной адаптации к окружающим условиям, является частью гомеостаза. Эти приспособления, сделанные телом, необходимы для выживания. Когда процесс гомеостаза успешен, жизнь продолжается. В случае неудачи может возникнуть дисбаланс, который может привести к смерти.
Определение гомеостаза
Состояние баланса во всех физических системах, необходимое для правильного функционирования и выживания организма, называется гомеостазом. В состоянии гомеостаза уровни тела постоянно регулируются в ответ на изменения снаружи и внутри тела. Некоторые из систем, которые постоянно приспосабливаются, чтобы оставаться на нормальном уровне:
- сахар в крови
- кровяное давление
- энергия
- кислотность
- кислород
- белки
- температура
- гормоны
- электролиты
Пример гомеостаза
Любая телесная система в динамическом равновесии достигает устойчивого состояния или баланса, который может противостоять внешним силам изменения. Когда эта система нарушается, регулирующие устройства, встроенные в тело, реагируют на создание нового баланса.
Один процесс называется управлением с обратной связью. Все процессы реализации и координации функций являются примерами гомеостаза. Это происходит при содействии нервной системы, гормональной системы или электрических токов.
Простым примером регулирования посредством гомеостаза является механическая система действия, которая контролирует температуру в помещении, т. е. термостат. Центр термостата представляет собой металлическую полосу, которая может воспринимать изменения температуры и реагировать, управляя электрической цепью.
Когда в помещении холодно, печь активируется термостатом, после чего температура повышается. В момент достижения заданного уровня контура термостата топка останавливается, а температура остается прежней или снижается.
Биологические системы намного сложнее, но могут достигать аналогичных результатов. Эти системы имеют регуляторы, сравнимые с механическими устройствами, и у них одинаковые цели: то есть регулировать систему и поддерживать условия в нормальном диапазоне. Это цель, будь то регулирование комнатной температуры или выравнивание давления в системе кровообращения.
Как работает регулирование обратной связи?
Важнейшей особенностью гомеостаза является то, что внутренняя среда организма поддерживается за счет узкого диапазона значений саморегулирующейся системы. И обратная связь, и прямая связь — это способы поддержания гомеостаза . Система обратной связи определяется как замкнутая структура, которая управляет будущими действиями, передавая в систему прошлые изменения во внутренней среде. Затем система меняет свое поведение, чтобы приспособиться к внешним условиям.
Существует два типа систем обратной связи. Они бывают отрицательными и положительными. Отрицательная обратная связь ищет цель и реагирует на любые неудачи в достижении этой цели. Он поддерживает постоянный диапазон значений.
Положительная обратная связь порождает процессы роста, когда действия извлекают выгоду из результатов, которые, в свою очередь, приводят к более масштабным действиям. Эти системы обратной связи подчиняются более высоким уровням контроля, и им может противодействовать отрицательная обратная связь 9.2354 .
Рабочий диапазон охватывает переменные, которые регулируются и могут реагировать на стимулы из окружающей среды. Результатом этих сложных взаимодействий и конкурирующих систем отрицательной и положительной обратной связи является гомеостаз, который является основой физиологической регуляции.
Почему важен гомеостаз?
Гомеостаз — центральная объединяющая концепция физиологии и саморегуляции, осуществляемая для поддержания внутренней стабильности. Гомеостаз непостоянен. Это динамический процесс, который изменяет внутренние условия, чтобы способствовать выживанию .
Регулирование посредством гомеостаза не является единичным циклом обратной связи. Он отражает сложное взаимодействие многих систем обратной связи, контролируемых нервами и гормонами. Эта обратная связь приводит к точному уровню контроля и гибкости, что позволяет организму приспосабливаться к изменениям условий окружающей среды .
Жизнь и здоровье организма являются конечным результатом регуляции посредством гомеостаза. Нарушение гомеостаза является причиной болезни.
Надлежащее лечение и терапия должны быть направлены на восстановление условий гомеостаза. В противном случае тяжелая болезнь или смерть могут быть неизбежны.
Как организм поддерживает гомеостаз?
Контроль температуры тела является хорошим общим примером того, как тело поддерживает гомеостаз в биологической системе. У людей средняя температура тела 98,6 градусов по Фаренгейту. Однако на это число могут повлиять различные факторы, в том числе:
- воздействие
- скорость метаболизма
- гормоны
- болезни
Все эти факторы могут привести к экстремально низкой или высокой температуре. Затем температура тела регулируется мозгом в области, называемой гипоталамусом. Обратная связь температуры тела передается через нервную систему в мозг. Это приводит к корректировке уровня сахара в крови, частоты дыхания и скорости метаболизма в попытке вернуться к гомеостазу.
Потеря тепла в организме зависит от потоотделения, снижения активности и функций теплообмена, которые позволяют большому количеству крови циркулировать на поверхности кожи. Потери тепла уменьшаются за счет уменьшения кожного кровообращения, изоляции и внешних приспособлений. Примеры включают поиск убежища, одежды и внешних источников тепла.
Разница между низкой и высокой температурой тела представляет собой гомеостатическое плато. Это нормальный диапазон, который позволяет продолжать жизнь. По мере того как уровни температуры приближаются к экстремальным значениям, коррекция с помощью отрицательной обратной связи помогает системе вернуться к нормальному диапазону.
Организм также поддерживает гомеостаз через систему кровообращения и барорецепторы. Барорецепторы — это чувствительные к давлению области кровеносных сосудов, которые реагируют на растяжение. Они посылают информацию о артериальном давлении в мозг, который посылает гормоны, вырабатываемые щитовидной железой и гипоталамусом, для регулирования обмена веществ в организме.
Где еще применяется гомеостаз?
Гомеостаз также использовался в экологических условиях. Впервые представленное экологом Робертом Макартуром, понимание гомеостаза в экосистемах достигается путем объединения биоразнообразия и взаимодействия между различными видами. Считалось, что эта концепция может объяснить стабильность в экосистеме. С годами он эволюционировал, включив в него неживые части, и некоторые считают его спорным.
Системы регуляции гомеостаза
Системы регуляции гомеостаза д-ра Джозефа Дж. Коллинза, RN, ND
Хотя в предыдущей статье обсуждалась концепция «усталости надпочечников», в ней также указывалось, что генерализованный адаптационный синдром является результатом воздействия стресса на многие системы регуляции гомеостаза. Мы понимаем, что существует несколько систем, вовлеченных в клинические состояния, которые проявляются в виде расстройств, связанных с утомляемостью. Различные системы регуляции гомеостаза, на которые влияет стресс и неспособность адаптироваться к стрессу, включают:
Ось гипоталамо-гипофизарно-надпочечниковая (ГГН) участвует в первичной реакции на стресс посредством взаимодействия между средним мозгом и корой надпочечников . При остром стрессе могут повышаться все гормоны надпочечников, включая кортизол, ДГЭА, а также прогестерон, альдостерон и андостендион. При хроническом стрессе уровень кортизола продолжает оставаться повышенным, в то время как другие гормоны снижаются. Высокий уровень кортизола может фактически вызвать резистентность кортикоидных рецепторов. Если стресс продолжается, происходит истощение надпочечников, что приводит к падению всех гормонов надпочечников. Изменения в гормонах надпочечников влияют на все другие регуляторные системы.
Симпатоадреналовая система (САС) участвует в первичном ответе на стресс посредством взаимодействия между симпатической нервной системой и мозговым веществом надпочечников . Стресс приводит к тому, что САС вызывает повышение уровня адреналина и норадреналина (адреналина и норадреналина), возбуждающих катехоламинов, которые также обладают свойствами нейротрансмиттера. Эти молекулы стимулируют различные системы организма и влияют на все остальные регуляторные системы.
гипоталамо-гипофизарно-гондальная (ГГГ) 9Ось 2335 контролирует рост, развитие, размножение и старение. Гипоталамус вырабатывает гонадотропин-рилизинг-гормон (ГнРГ), после чего передняя часть гипофиза вырабатывает лютеинизирующий гормон (ЛГ) и фолликулостимулирующий гормон (ФСГ), в результате чего гонады вырабатывают половые гормоны, такие как эстроген и/или тестостерон. Длительный или чрезмерный стресс может нарушить функцию HPG, что приведет к снижению выработки половых гормонов. Снижение половых гормонов делает организм более восприимчивым к стрессу.
Ось гипоталамо-гипофизарно-щитовидная железа (ГГТ) отвечает за регуляцию метаболизма. Гипоталамус выделяет тиреотропин-рилизинг-гормон (ТРГ), когда уровень гормонов щитовидной железы низкий. Это стимулирует гипофиз к выработке тиреотропного гормона (ТТГ), который стимулирует щитовидную железу вырабатывать больше гормонов щитовидной железы. Длительный или чрезмерный стресс может нарушить функцию ГГТ, что приведет к снижению выработки гормонов щитовидной железы. Снижение гормонов щитовидной железы делает организм более восприимчивым к стрессу.
Нейроэндокринно-иммунная (НЭИ) система представляет собой взаимодействие между нервной системой, эндокринной системой и иммунной системой в ответ на стресс. Усиление реакции иммунной системы может быть полезным при травматических и инфекционных стрессовых событиях. Однако длительная или неуместная активация NEI отрицательно повлияет на каждую систему организма, каждую регуляторную систему.
Глюкозо-инсулиновая система (ГИС) отвечает за поддержание соответствующего уровня глюкозы в крови. Во время аллостаза ГИС может допускать более высокие уровни глюкозы в ответ на острый стресс. Если аллостаз сохраняется, то неадекватное повышение уровня глюкозы будет иметь неблагоприятные последствия для всего организма. Повышенный уровень глюкозы может вызвать чрезмерную выработку инсулина, что может вызвать проблемы с гипогликемией (низкий уровень сахара в крови) в краткосрочной перспективе, но привести к резистентности к инсулину (преддиабету), если он продолжится. Ненормальный уровень глюкозы делает организм более восприимчивым к стрессу. Например, стресс от единичного эпизода гипогликемии повышает вероятность того, что у человека будут более частые и более интенсивные приступы гипогликемии. Это хорошо задокументированный пример дисфункции регуляторной системы, ухудшающейся с течением времени.
Моноаминергическая система мозга (BMS) отвечает за балансировку возбуждающих и тормозных нейротрансмиттеров в головном мозге. BMS может допускать более высокие уровни возбуждающих нейротрансмиттеров, таких как адреналин и норадреналин, в ответ на острый стресс. Однако длительное или неадекватное повышение уровня возбуждающих нейротрансмиттеров будет иметь неблагоприятное воздействие на мозг, а также на другие системы организма.
Ренин-ангиотензин-альдостероновая система (РААС) представляет собой гормональную систему, которая регулирует кровяное давление, электролитный и водный баланс. Аллостатическая реакция РААС способствует задержке жидкости и натрия и вазоконстрикции в ответ на острый стресс. Если гомеостаз не восстанавливается, эта длительная задержка жидкости и натрия, а также вазоконстрикция могут привести к высокому кровяному давлению, которое оказывает неблагоприятное воздействие на все системы организма. Задержка жидкости и высокое кровяное давление делают организм более восприимчивым к стрессу.
Система детоксикации печени (HDS) отвечает за снижение токсичности молекул из эндогенных и экзогенных источников посредством процессов детоксикации фазы 1 и фазы 2. Повышение уровня катехоламинов, вызванное стрессом, снижает эффективность HDS, что может повлиять на здоровье каждой системы организма и каждой регуляторной системы и сделать организм более восприимчивым к стрессу.
Система сотовой сигнализации (CCS) представляет собой очень сложную систему межклеточной связи, которая поддерживает правильное функционирование клеток во всем организме. Стресс может нарушать клеточную сигнализацию, изменяя количество и/или тип сигнальных белков или снижая функцию клеточных рецепторов. Наиболее частым отклонением от нормальной передачи клеточных сигналов является развитие резистентности рецепторов, при которой клетки организма становятся устойчивыми — и игнорируют — посылаемые им сообщения. Инсулинорезистентность является наиболее известным примером резистентности клеточных рецепторов. Другим примером является резистентность глюкокортикоидных рецепторов, которая отрицательно влияет на то, как клетки реагируют на кортизол 9.0008
Система восстановления-окисления (REDOX) — это системы, в которых окислительные и восстановительные биологические процессы происходят во всем организме. Окисление и восстановление являются важными биологическими процессами, происходящими в каждой клетке. Одним из наиболее важных примеров активности РЕДОКС-активности в организме является РЕДОКС-активность, происходящая в митохондриях каждой клетки. Митохондрии — это органеллы, вырабатывающие энергию, которые создают энергию, необходимую каждой клетке для функционирования. Стресс может нарушить баланс между окислением и восстановлением и снизить эффективность биологических процессов.
Система грелин-лептин-адипонектин (GLAS) представляет собой сложную систему, которая регулирует аппетит, метаболизм, энергетический гомеостаз и рост. GLAS динамически взаимодействует с гормоном роста, ИФР-1 и ИФР-2, а также с инсулином. Подавляющее аппетит действие лептина и адипонектина снижается при стрессе. Грелин, усиливающий чувство голода, повышается при стрессе. Острый стресс может ингибировать лептин и адипонектин, в то время как хронический стресс и воспаление при ожирении могут вызывать повышение лептина и резистентности к лептину, что приводит к избыточному потреблению энергии. Психический стресс может увеличить уровень грелина.
Гомеостаз и аллостаз
Эти системы регуляции гомеостаза предназначены для поддержания стабильного, относительно постоянного равновесия во внутренней среде. Одна система может контролировать уровень сахара в крови; другой может контролировать жидкость и электролиты. Другие системы могут контролировать уровни гормонов или нейротрансмиттеров, балансировать антиоксиданты или контролировать детоксикацию. Когда система поддерживает баланс, говорят, что она находится в гомеостазе. Гомеостаз буквально означает «похожее состояние» — так что все примерно одинаково, что идеально. Когда система выходит из равновесия, она находится в аллостазе, что означает «другое состояние» — все не идеально. Тело может перейти в другое состояние, когда оно должно ответить на запрос. Частота сердечных сокращений, дыхание и температура тела увеличиваются при физической нагрузке, что является здоровым аллостазом. Тело обычно возвращается к гомеостазу после завершения упражнения. Но если частота сердечных сокращений, дыхания и температура тела повышены из-за болезни, например сильной лихорадки, то это нездоровый аллостаз.
AdrenoMend™ — это препарат, специально предназначенный для гормонов, который был разработан для поддержки функции каждой системы регуляции гомеостаза. Узнайте больше об AdrenoMend™.
Знакомство с признаками и симптомами
Когда тело застревает в хроническом состоянии аллостаза с усталостью, болями, сыпью, нарушением мышления и другими хроническими признаками и симптомами, оно застревает в состоянии аллостаза. Важно отметить, что большинство признаков и симптомов связаны с аллостазом различных систем регуляции гомеостаза. Пример симптомов усталости, которые могут быть вызваны аллостазом гипоталамо-гипофизарно-надпочечниковой (ГГН) оси, гипоталамо-гипофизарно-тиреоидной (ГГТ) оси или моноаминергической системы головного мозга (ММС), и это лишь некоторые из них. Другим примером признака может быть высокое кровяное давление, которое может быть связано с аллостазом ренин-ангиотензин-альдостероновой системы (РААС) или симпато-адреномедуллярной системы (САС).
Далее: Узнайте больше о признаках и симптомах «усталости надпочечников»
Биологические системы: гомеостаз | Техасский шлюз
Введение в гомеостаз и механизмы обратной связи Примеры гомеостаза и механизмов обратной связи Обзор гомеостаза и механизмов обратной связи
Подумайте, как вы обогреваете и охлаждаете свой дом. У вас есть системы отопления и кондиционирования. Когда на улице жарко, кондиционер охлаждает ваш дом. И наоборот, когда на улице холодно, печь обогревает ваш дом.
Целью этих систем является поддержание внутри вашего дома постоянной комфортной температуры, несмотря на изменения температуры снаружи.
Это состояние постоянных условий можно назвать равновесным , или устойчивым состоянием . В данном примере состоянием равновесия является постоянная температура внутри дома.
Точно так же в вашем организме есть системы, поддерживающие стабильное состояние внутренней среды для многих различных параметров, таких как температура тела или уровень глюкозы в крови. Способность организма поддерживать постоянство внутренней среды в ответ на изменения внешней среды называется гомеостаз .
Подобно системам отопления и кондиционирования воздуха в доме, ваше тело имеет системы, которые реагируют на изменения в окружающей среде для поддержания гомеостаза. Эти механизмы называются механизмами обратной связи .
Процитировать источник
Изображения из Microsoft Word 2010.
Сосредоточьтесь на механизмах обратной связи
1. Отрицательный отзыв
Теперь рассмотрим, что происходит, когда жаркая погода согревает дом. Температура внутри выходит из равновесия. Он поднимается выше уставки термостата. Это повышение температуры вызывает срабатывание термостата, который включает кондиционер. Кондиционер охлаждает дом до достижения заданной температуры, а затем отключается. Теперь в доме вернулась прежняя температура.
Это пример отрицательного отзыва . При отрицательной обратной связи механизм действует в направлении, противоположном первоначальному изменению, так что установившееся состояние восстанавливается. Большинство гомеостатических механизмов представляют собой отрицательную обратную связь.
2. Положительная обратная связь
Теперь предположим, что техник HVAC пересек провода в системе. Провода были перекрещены, чтобы при повышении температуры в доме вместо кондиционера срабатывала печь. Печь обогревала дом, что еще больше повышало температуру. Повышенная температура продолжала поддерживать работу печи, что еще больше нагревало дом. Из этого сценария видно, что в доме становится все жарче, а не возвращается к заданному значению термостата.
Это пример положительных отзывов . При положительной обратной связи механизм действует в том же направлении, что и первоначальное изменение. Стационарное состояние никогда не восстанавливается, и благоприятны экстремальные условия. Лишь в нескольких случаях гомеостатические механизмы представляют собой петли положительной обратной связи. Одним из примеров являются роды и родоразрешение во время родов. Во время родов выделяется гормон окситоцин, который усиливает и ускоряет схватки. Усиление схваток приводит к высвобождению большего количества окситоцина, и цикл продолжается до тех пор, пока ребенок не родится. Рождение прекращает выброс окситоцина и завершает механизм положительной обратной связи.
Процитировать источник
Изображения из Microsoft Word 2010.
Части механизма обратной связи
Давайте продолжим думать о системах отопления и охлаждения дома. Датчик, такой как термопара, определяет температуру внутри дома и отправляет информацию на термостат. Термостат сравнивает температуру с заданной. Если температура выше заданной, термостат включает кондиционер. Если температура ниже заданной, термостат включает печь. Если температура соответствует заданной, ничего не делается.
Механизмы обратной связи состоят из датчиков, интеграторов и эффекторов:
- Датчик: Контролирует окружающую среду (внешнюю или внутреннюю) и отправляет информацию интегратору.
- Интегратор: Обрабатывает информацию от датчиков и отправляет сигналы или команды эффекторам.
- Эффектор: Принимает меры для восстановления стабильного состояния среды.
Примечание: Датчики называются рецептора в биологических системах.
Процитировать источник
Изображения из Microsoft Word 2010.
Регулирование температуры тела
Учтите, что вы тренируетесь под палящим солнцем. Когда температура вашего тела повышается, у вас есть гомеостатический механизм для ее снижения:
- Рецепторы кожи обнаруживают повышенную температуру. Они посылают информацию нервными импульсами к температурным центрам в гипоталамусе.
- Гипоталамус интегрирует информацию и посылает нервные импульсы в капилляры кожи и в потовые железы.
- Кровеносные сосуды в коже становятся шире, что увеличивает приток крови к коже. Увеличенный кровоток передает тепло, которое теряется в воздухе через кожу. Потовые железы выделяют пот, который испаряется и охлаждает кожу.
- Температура тела снижается почти до нормы. Это продолжается до тех пор, пока вы не покинете солнечную зону и не прекратите тренироваться. Затем температура тела возвращается к норме, и вы перестаете потеть.
Обратите внимание, что это механизм отрицательной обратной связи.
И наоборот, если вы выходите на улицу в холодный день, температура вашего тела снижается.
- Рецепторы кожи обнаруживают понижение температуры. Они посылают информацию нервными импульсами к температурным центрам в гипоталамусе.
- Гипоталамус интегрирует информацию и посылает нервные импульсы в капилляры кожи и скелетные мышцы.
- Кровеносные сосуды в коже сужаются, что снижает приток крови к коже. Уменьшение кровотока уменьшает потери тепла в воздух через кожу. Мышцы быстро сокращаются (дрожь), в результате чего выделяется тепло.
- Эти действия повышают температуру тела.
Газообмен в листьях
Гомеостатические механизмы не ограничиваются животными. Они есть и у растений.
Вот один пример. В листьях есть отверстия, называемые устьицами (множественное число: устьица). Вокруг стомы расположены замыкающие клетки. Газы, такие как углекислый газ, кислород и водяной пар, входят и выходят из листа через эти отверстия. Вот как работает механизм:
В течение дня достаточно солнечного света для фотосинтеза. Фотосинтез в замыкающих клетках вызывает накопление в них ионов калия и воды (путем осмоса). Замыкающие клетки набухают и открывают стому. В результате газы могут проникать в лист и выходить из него. И наоборот, ночью фотосинтез прекращается. Замыкающие клетки удаляют калий и воду (путем осмоса). Замыкающие клетки сокращаются и закрывают стому. Впоследствии никакие газы не могут попасть внутрь листа или выйти из него.
Процитировать источник
Изображения из Microsoft Word 2010.
Свойства гомеостатических механизмов
При идентификации или оценке гомеостатического механизма ищите следующие свойства: либо внутреннюю среду, либо внешнюю среду.
Давайте потренируемся. Ниже приведены некоторые механизмы обратной связи. Определите рецепторы, интеграторы и эффекторы. Определите, является ли это механизмом отрицательной или положительной обратной связи.
- Печать
- Поделиться
Гомеостаз – анатомия и физиология
Введение в человеческое тело
OpenStaxCollege
Цели обучения
К концу этого раздела вы сможете:
- Обсудить роль гомеостаза в здоровом функционировании
- Сравните отрицательную и положительную обратную связь, предоставив по одному физиологическому примеру каждого механизма
Поддержание гомеостаза требует, чтобы организм постоянно следил за своим внутренним состоянием. От температуры тела до артериального давления и уровней определенных питательных веществ, каждое физиологическое состояние имеет определенную заданную точку. Уставка – это физиологическое значение, вокруг которого колеблется нормальный диапазон. Нормальный диапазон — это ограниченный набор значений, который является оптимально здоровым и стабильным. Например, уставка нормальной температуры тела человека составляет приблизительно 37°C (98,6°F) Физиологические параметры, такие как температура тела и артериальное давление, имеют тенденцию колебаться в пределах нормы на несколько градусов выше и ниже этой точки. Центры управления в мозгу и других частях тела отслеживают отклонения от гомеостаза и реагируют на них с помощью отрицательной обратной связи. Отрицательная обратная связь — это механизм, который устраняет отклонение от заданного значения. Следовательно, отрицательная обратная связь поддерживает параметры тела в пределах их нормального диапазона. Поддержание гомеостаза с помощью отрицательной обратной связи постоянно происходит во всем теле, и поэтому понимание отрицательной обратной связи имеет фундаментальное значение для понимания физиологии человека.
Система отрицательной обратной связи состоит из трех основных компонентов ([ссылка] и ). Датчик, также называемый рецептором, является компонентом системы обратной связи, которая отслеживает физиологическое значение. Это значение сообщается в центр управления. Центр управления — это компонент системы обратной связи, который сравнивает значение с нормальным диапазоном. Если значение слишком сильно отклоняется от заданного значения, центр управления активирует эффектор. Эффектор — это компонент системы обратной связи, который вызывает изменение, направленное на изменение ситуации и возвращение значения к нормальному диапазону.
Петля отрицательной обратной связи
В петле отрицательной обратной связи стимул — отклонение от заданного значения — сопротивляется физиологическому процессу, возвращающему организм к гомеостазу. (а) Петля отрицательной обратной связи состоит из четырех основных частей. (б) Температура тела регулируется отрицательной обратной связью.
Чтобы привести систему в движение, стимул должен вывести физиологический параметр за пределы его нормального диапазона (то есть за пределы гомеостаза). Этот стимул «услышан» определенным датчиком. Например, при контроле уровня глюкозы в крови специфические эндокринные клетки поджелудочной железы обнаруживают избыток глюкозы (стимул) в кровотоке. Эти бета-клетки поджелудочной железы реагируют на повышенный уровень глюкозы в крови, высвобождая гормон инсулин в кровоток. Инсулин дает сигнал скелетным мышечным волокнам, жировым клеткам (адипоцитам) и клеткам печени поглощать избыток глюкозы, удаляя ее из кровотока. По мере того, как концентрация глюкозы в кровотоке падает, уменьшение концентрации — фактическая отрицательная обратная связь — обнаруживается альфа-клетками поджелудочной железы, и выделение инсулина прекращается. Это предотвращает дальнейшее падение уровня сахара в крови ниже нормального диапазона.
Люди имеют аналогичную систему обратной связи регулирования температуры, которая работает, способствуя либо потере тепла, либо увеличению тепла ([ссылка] b ). Когда центр регуляции температуры мозга получает данные от датчиков, указывающие на то, что температура тела превышает нормальный диапазон, он стимулирует группу клеток мозга, называемую «центром потери тепла». Эта стимуляция имеет три основных эффекта:
- Кровеносные сосуды в коже начинают расширяться, позволяя большему количеству крови из ядра тела течь к поверхности кожи, позволяя теплу излучаться в окружающую среду.
- Когда приток крови к коже увеличивается, потовые железы активизируются, увеличивая их выработку. Когда пот испаряется с поверхности кожи в окружающий воздух, он уносит с собой тепло.
- Увеличивается глубина дыхания, и человек может дышать через открытый рот, а не через носовые ходы. Это еще больше увеличивает потерю тепла из легких.
Напротив, активация центра накопления тепла в мозге под воздействием холода снижает приток крови к коже, а кровь, возвращающаяся из конечностей, направляется в сеть глубоких вен. Такое расположение улавливает тепло ближе к ядру тела и ограничивает потери тепла. Если потеря тепла серьезна, мозг запускает усиление случайных сигналов к скелетным мышцам, заставляя их сокращаться и вызывая дрожь. Сокращения мышц при дрожи выделяют тепло при расходовании АТФ. Мозг запускает щитовидную железу в эндокринной системе для высвобождения гормона щитовидной железы, который увеличивает метаболическую активность и выработку тепла в клетках по всему телу. Мозг также сигнализирует надпочечникам о высвобождении эпинефрина (адреналина), гормона, вызывающего расщепление гликогена на глюкозу, которую можно использовать в качестве источника энергии. Распад гликогена на глюкозу также приводит к усилению метаболизма и выработке тепла.
Концентрация воды в организме имеет решающее значение для нормального функционирования. Тело человека сохраняет очень жесткий контроль над уровнем воды без сознательного контроля со стороны человека. Посмотрите это видео, чтобы узнать больше о концентрации воды в организме. Какой орган в первую очередь контролирует количество воды в организме?
Положительная обратная связь усиливает изменение физиологического состояния организма, а не обращает его вспять. Отклонение от нормального диапазона приводит к большим изменениям, и система отдаляется от нормального диапазона. Положительная обратная связь в организме нормальна только тогда, когда есть определенная конечная точка. Роды и реакция организма на кровопотерю — два примера положительной обратной связи, которая является нормальной, но активируется только при необходимости.
Роды в срок являются примером ситуации, в которой сохранение существующего состояния организма нежелательно. Для изгнания ребенка в конце беременности требуются огромные изменения в организме матери. И события родов, однажды начавшись, должны быстро развиваться до завершения, иначе жизни матери и ребенка угрожает опасность. Экстремальная мышечная работа во время родов является результатом системы положительной обратной связи ([ссылка]).
Петля положительной обратной связи
Нормальные роды управляются положительной обратной связью. Петля положительной обратной связи приводит к изменению состояния организма, а не к возврату к гомеостазу.
Первые схватки (стимул) подталкивают ребенка к шейке матки (самая нижняя часть матки). Шейка матки содержит чувствительные к растяжению нервные клетки, которые контролируют степень растяжения (сенсоры). Эти нервные клетки посылают сообщения в мозг, который, в свою очередь, заставляет гипофиз в основании мозга выделять гормон окситоцин в кровоток. Окситоцин вызывает более сильные сокращения гладких мышц матки (эффекторов), проталкивая ребенка дальше по родовым путям. Это вызывает еще большее растяжение шейки матки. Цикл растяжения, выброса окситоцина и все более сильных сокращений прекращается только с рождением ребенка. В этот момент растяжение шейки матки прекращается, прекращая выброс окситоцина.
Второй пример положительной обратной связи заключается в устранении экстремальных повреждений тела. После проникающего ранения самой непосредственной угрозой является чрезмерная кровопотеря. Меньшая циркуляция крови означает снижение артериального давления и снижение перфузии (проникновения крови) в мозг и другие жизненно важные органы. Если перфузия сильно снижена, жизненно важные органы отключаются, и человек умирает. Организм реагирует на эту возможную катастрофу выбросом в поврежденную стенку кровеносного сосуда веществ, запускающих процесс свертывания крови. По мере того, как происходит каждый этап свертывания крови, он стимулирует высвобождение большего количества свертывающих веществ. Это ускоряет процессы свертывания крови и запечатывания поврежденного участка. Свертывание происходит в локальной области благодаря строго контролируемой доступности белков свертывания крови. Это адаптивный, спасительный каскад событий.
Гомеостаз — это активность клеток во всем организме для поддержания физиологического состояния в узком диапазоне, совместимом с жизнью. Гомеостаз регулируется петлями отрицательной обратной связи и, гораздо реже, петлями положительной обратной связи. Оба имеют одни и те же компоненты стимула, сенсора, центра управления и эффектора; однако петли отрицательной обратной связи предотвращают чрезмерную реакцию на стимул, тогда как петли положительной обратной связи усиливают реакцию до тех пор, пока не будет достигнута конечная точка.
Концентрация воды в организме имеет решающее значение для нормального функционирования. Тело человека сохраняет очень жесткий контроль над уровнем воды без сознательного контроля со стороны человека. Посмотрите это видео, чтобы узнать больше о концентрации воды в организме. Какой орган в первую очередь контролирует количество воды в организме?
Почки.
После того, как вы пообедали, нервные клетки в желудке реагируют на растяжение (стимул), вызванное едой. Они передают эту информацию ________.
- центр управления
- уставка
- эффекторы
- датчики
А
Стимуляция центра теплоотдачи вызывает ________.
- кровеносные сосуды кожи для сужения
- дыхание становится медленным и поверхностным
- потовые железы для повышения их производительности
- Все вышеперечисленное
C
Что из перечисленного является примером нормального физиологического процесса, в котором используется петля положительной обратной связи?
- регулирование артериального давления
- роды
- регулировка баланса жидкости
- регулирование температуры
Б
Определите четыре компонента петли отрицательной обратной связи и объясните, что произойдет, если секреция химического вещества в организме, контролируемая системой отрицательной обратной связи, станет слишком большой.
Четыре компонента петли отрицательной обратной связи: стимул, датчик, центр управления и эффектор. Если выделялось слишком большое количество химического вещества, сенсоры активировали центр управления, который, в свою очередь, активировал эффектор. В этом случае эффектор (секретирующие клетки) будет смещен вниз.
Какие регулирующие процессы задействовал бы ваш организм, если бы вы оказались в ловушке из-за метели в неотапливаемой неизолированной хижине в лесу?
Любое длительное воздействие сильного холода активирует центр накопления тепла в мозгу. Это уменьшит приток крови к вашей коже и перенаправит кровь, возвращающуюся из ваших конечностей, от пальцев в сеть глубоких вен. Центр накопления тепла в вашем мозгу также усилит сокращение мышц, заставляя вас дрожать. Это увеличивает потребление энергии скелетными мышцами и вырабатывает больше тепла. Ваше тело также будет производить гормон щитовидной железы и адреналин, химические вещества, которые способствуют увеличению метаболизма и выработке тепла.