Биологические ритмы. Фотопериодизм – онлайн-тренажер для подготовки к ЕНТ, итоговой аттестации и ВОУД
Многие явления в природе имеют явно ритмичный характер: периоды солнечной активности (11-летний цикл), смена времен года (весна, лето, осень, зима), фаз луны, изменение времени суток (день, ночь) и т. д.
Биологические ритмы – периодически повторяющиеся изменения активности процессов жизнедеятельности организмов.
Различают суточные и годовые ритмы активности живых организмов. А для обитателей побережий морей и океанов, где наблюдаются такие явления, как прилив и отлив, характерны приливно-отливные ритмы. Исключение составляют бактерии и вирусы, наличие ритмов у которых пока не доказано.
Большинство биологических процессов также имеют ритмичный характер, связанный или не связанный с внешними ритмами. Например, сезонные и суточные изменения у растений. У животных это годовые, месячные и суточные ритмы, например, период зимней спячки, линька, период сна и бодрствования, более частые ритмы мозга и сердца.
Суточные ритмы – ритмы, которые приспосабливают организмы к смене дня и ночи. Причиной суточных ритмов является вращение Земли вокруг своей оси. Суточные ритмы обнаружены как у многоклеточных, так и у одноклеточных организмов. У растений интенсивный рост, распускание цветков, закрывание и открывание устьиц приурочены к определенному времени суток. Наблюдаются ритмы и в протекании процессов дыхания и фотосинтеза, что проявляется в их усилении или ослаблении. У животных сильно меняется активность в течение суток. По этому признаку различают дневных и ночных животных. Проявляются суточные ритмы в чередовании сна и бодрствования, изменении двигательной активности, частоты пульса, температуры тела.
Годовые ритмы – это ритмы, которые приспосабливают организмы к сезонной смене условий. Причина: движение Земли вокруг Солнца. При годовом движении Земли по орбите вокруг Солнца на нашей планете происходит смена времен года: зимы, весны, лета и осени. В жизни видов периоды роста, размножения, линек, миграций, глубокого покоя закономерно чередуются и повторяются таким образом, что критическое время года организмы встречают в наиболее устойчивом состоянии. Самый же уязвимый процесс – размножение и выращивание молодняка – приходится на самый благоприятный период. На этот же период приходится цветение растений, созревание плодов и семян (вегетационный период). Подобные биологические ритмы обнаружены у всех организмов. Внутренний механизм, позволяющий организму не только чувствовать течение времени, но и измерять его промежутки, называется биологическими часами. Иногда биологические часы могут нарушаться, например, при длительных полетах и пересечении часовых поясов. Хорошо известно, что у летчиков и стюардесс часто нарушается время сна и бодрствования. Наука, изучающая биоритмы, называется хронобиологией. Жизнь на Земле развивалась в условиях регулярной смены дня и ночи и чередования времен года из-за вращения планеты вокруг своей оси.
В жизни видов периоды роста, размножения, линек, миграций, глубокого покоя закономерно чередуются и повторяются таким образом, что критическое время года организмы встречают в наиболее устойчивом состоянии. Самый же уязвимый процесс – размножение и выращивание молодняка – приходится на самый благоприятный период. На этот же период приходится цветение растений, созревание плодов и семян (вегетационный период). Подобные биологические ритмы обнаружены у всех организмов. Внутренний механизм, позволяющий организму не только чувствовать течение времени, но и измерять его промежутки, называется биологическими часами. Иногда биологические часы могут нарушаться, например, при длительных полетах и пересечении часовых поясов. Хорошо известно, что у летчиков и стюардесс часто нарушается время сна и бодрствования. Наука, изучающая биоритмы, называется хронобиологией. Жизнь на Земле развивалась в условиях регулярной смены дня и ночи и чередования времен года из-за вращения планеты вокруг своей оси. Знание сформировавшихся закономерностей биоритмов имеет большое значение в сельском хозяйстве и в профилактической медицине.
Знание сформировавшихся закономерностей биоритмов имеет большое значение в сельском хозяйстве и в профилактической медицине.
Фотопериодизм. Многим организмам свойственна реакция на суточные ритмы освещения, т. е. на соотношение светлого (длина дня) и темного (длина ночи) периодов суток. Реакция, выражающаяся в изменении процессов роста и развития, называется фотопериодизмом (от греч. фотос – свет и период). В зависимости от длительности освещения растения делятся на растения длинного дня и растения короткого дня. Растения длинного дня растут в основном в умеренных и приполярных широтах. Например, растение дурнишник зацветает только тогда, когда длина светлого времени суток достигает 21 ч. Если длина светлого времени будет меньше 21 ч, то это растение может попасть под заморозки. Южные растения соя, бамбук, хлопчатник, просо, кукуруза, табак являются короткодневными. Они быстрее зацветают в условиях короткого дня. Фотопериодизм присущ не только растениям, но и животным. Так, у птиц и крупных млекопитающих с фотопериодизмом связана сезонная миграция, осенняя и весенняя линьки, переход к зимней спячке и многое другое. Не менее важна фотопериодическая регуляция и для сезонной половой активности животных. Фотопериодизм влияет и на пищевое поведение: под влиянием коротких дней животные умеренных широт начинают искать более калорийную пищу. Фотопериодизм у человека влияет на сезонные эмоциональные состояния, например, всем известен весенний пик эмоциональной активности.
Так, у птиц и крупных млекопитающих с фотопериодизмом связана сезонная миграция, осенняя и весенняя линьки, переход к зимней спячке и многое другое. Не менее важна фотопериодическая регуляция и для сезонной половой активности животных. Фотопериодизм влияет и на пищевое поведение: под влиянием коротких дней животные умеренных широт начинают искать более калорийную пищу. Фотопериодизм у человека влияет на сезонные эмоциональные состояния, например, всем известен весенний пик эмоциональной активности.
БИОЛОГИЧЕСКИЕ РИТМЫ • Большая российская энциклопедия
БИОЛОГИ́ЧЕСКИЕ РИ́ТМЫ (биоритмы), периодически повторяющиеся изменения интенсивности и характера биологич. процессов и явлений. Под Б. р. в широком смысле понимают все циклич. процессы, протекающие в живых системах разного уровня организации. Периоды этих циклов варьируют от миллисекундных колебаний нейрональной активности до многолетних вариаций динамики популяций. Среди них выделяют т. н. рабочие, или физиологические, ритмы, которые являются необходимым условием функционирования биологич. систем (напр., ритмы дыхания и сокращений сердца). Их частоты определяются как физико-химич. особенностями ритмически работающих органов, так и потребностями организма. Так, во время сна сердечный ритм человека обычно составляет от 40 до 90 ударов в минуту, а при интенсивной физич. работе может возрастать до 200 и более ударов. Кроме того, всем живым организмам свойственны колебания, периоды которых равны осн. геофизич. циклам, обусловленным вращением Земли вокруг собств. оси (суточный ритм) и вокруг Солнца (годовой, или сезонный, ритм). Мн. виды организмов реагируют на изменения ночной освещённости, связанной с фазами Луны, а активность обитателей морского побережья подчиняется также ритмам приливов и отливов.
Среди них выделяют т. н. рабочие, или физиологические, ритмы, которые являются необходимым условием функционирования биологич. систем (напр., ритмы дыхания и сокращений сердца). Их частоты определяются как физико-химич. особенностями ритмически работающих органов, так и потребностями организма. Так, во время сна сердечный ритм человека обычно составляет от 40 до 90 ударов в минуту, а при интенсивной физич. работе может возрастать до 200 и более ударов. Кроме того, всем живым организмам свойственны колебания, периоды которых равны осн. геофизич. циклам, обусловленным вращением Земли вокруг собств. оси (суточный ритм) и вокруг Солнца (годовой, или сезонный, ритм). Мн. виды организмов реагируют на изменения ночной освещённости, связанной с фазами Луны, а активность обитателей морского побережья подчиняется также ритмам приливов и отливов.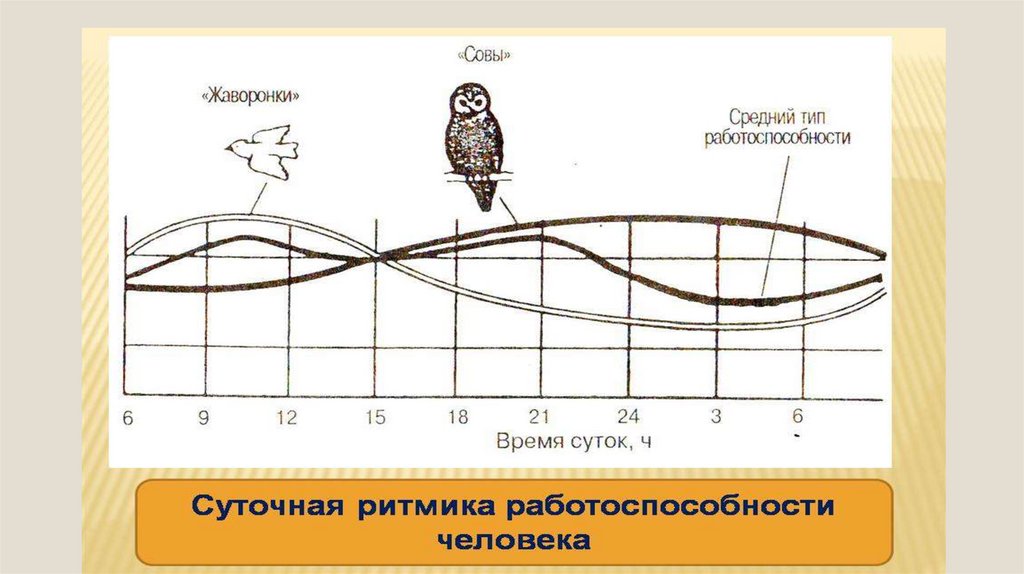 Такие Б. р. называют адаптивными, т. к. они направлены на координацию жизнедеятельности организмов при колебаниях абиотических и биотических параметров среды. Напр., у растений ритмы фотосинтеза непосредственно определяются чередованием светлого и тёмного времени суток (фотопериодом). Ночная активность, характерная для мн. видов грызунов, повышает вероятность их выживания в условиях пресса хищников. Благодаря годовым ритмам, особенно значимым в условиях резко континентального климата, растения и животные могут заблаговременно подготовиться к изменению погодных условий, напр. к зимнему похолоданию. При этом одни из них залегают в спячку (сурки, суслики), у других уплотняется меховой покров и усиливается способность к термогенезу (лисицы, белки, хомячки и др.), у третьих происходит накопление во внеклеточной жидкости веществ, предотвращающих её замерзание при минусовых темп-рах (лягушки, улитки, некоторые виды растений и др.
Такие Б. р. называют адаптивными, т. к. они направлены на координацию жизнедеятельности организмов при колебаниях абиотических и биотических параметров среды. Напр., у растений ритмы фотосинтеза непосредственно определяются чередованием светлого и тёмного времени суток (фотопериодом). Ночная активность, характерная для мн. видов грызунов, повышает вероятность их выживания в условиях пресса хищников. Благодаря годовым ритмам, особенно значимым в условиях резко континентального климата, растения и животные могут заблаговременно подготовиться к изменению погодных условий, напр. к зимнему похолоданию. При этом одни из них залегают в спячку (сурки, суслики), у других уплотняется меховой покров и усиливается способность к термогенезу (лисицы, белки, хомячки и др.), у третьих происходит накопление во внеклеточной жидкости веществ, предотвращающих её замерзание при минусовых темп-рах (лягушки, улитки, некоторые виды растений и др. ).
).
Б. р. не являются простым отражением колебаний условий во внешней среде. Растения, животные или микроорганизмы, изолированные от осн. геофизич. циклов («датчиков времени»), сохраняют в течение длительного времени присущую им ритмич. активность. Однако наблюдаемые в условиях изоляции периоды свободнотекущих ритмов близки, но не равны внешним циклам. Из-за этого несовпадения Б. р. получили назв. «циркаритмы» (от лат. circa – около). Соответственно колебания с периодом близким к 24 часам называются околосуточными или циркадными (от лат. dies – день) ритмами, а с периодом близким к 12 месяцам – окологодовыми или цирканнуальными (от лат. annual – год) ритмами. Устойчивое отклонение периодов свободнотекущих ритмов от внешних циклов служит гл. аргументом в пользу их эндогенной природы. С этих позиций суточные, годовые (сезонные), лунные и приливные колебания функций организма рассматриваются как результат синхронизации биологич.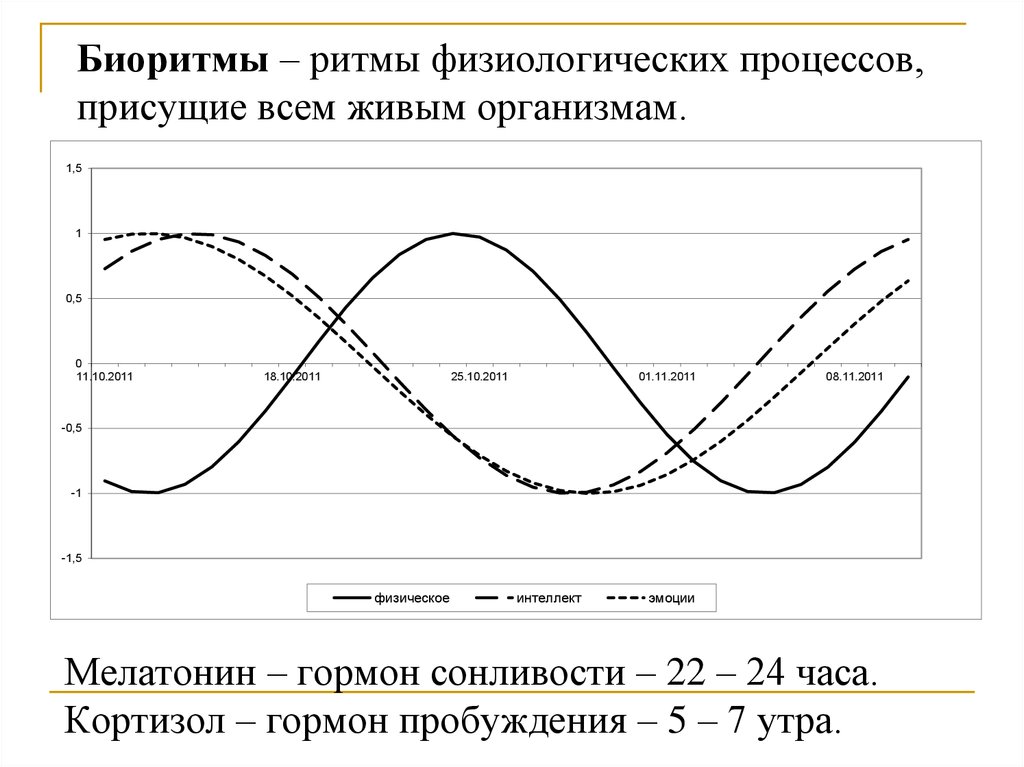 часов внешними циклич. процессами, среди которых ведущую роль играет фотопериод.
часов внешними циклич. процессами, среди которых ведущую роль играет фотопериод.
Периоды свободнотекущих ритмов могут не совпадать у разных систем организма. Так, у людей, изолированных от внешних датчиков времени, периоды колебаний темп-ры тела, выведения калия и гормонов коры надпочечников с мочой в ср. равны 25 часам. В то же время периоды ритмов сна и бодрствования, выведения кальция и натрия с мочой у 1/3 людей могут либо превышать 40 часов, либо быть короче 20 часов. Это свидетельствует о способности отд. систем организма к самостоят. генерации циклов. Представления о множественной локализации биологич. часов с циркадным периодом подкреплены исследованиями изолированных органов (надпочечники, сердце, печень и др.), сохраняющих ритмич. активность в течение нескольких суток вне организма. Но даже у одноклеточной водоросли Gonialax poliedra периоды свободнотекущих ритмов хемолюминисценции и передвижения могут не совпадать друг с другом, т. е. отд. клетка также обладает несколькими часами.
е. отд. клетка также обладает несколькими часами.
Рис. 1. Циркадная организация млекопитающих и человека. Центральные биологические часы в супрахиазматических ядрах (СХЯ) гипоталамуса благодаря нервным (вегетативная нервная система, ВНС) и эндокринны…
Пока детально изучены лишь механизмы эндогенных околосуточных ритмов. Их молекулярно-генетич. основу составляют внутриклеточные системы с отрицательной обратной связью, в которых полный цикл – экспрессия специализир. генов биологич. часов (т. н. часовых генов), подавление их активности собственными белковыми продуктами и последующая деградация белков – занимает ок. 24 часов. Дополнительные контуры обратной связи, регулирующие образование факторов транскрипции, дающих начало следующему циклу, стабилизируют работу биологич. часов. Периоды свободнотекущих ритмов остаются неизменными в широком диапазоне температур даже у одноклеточных организмов.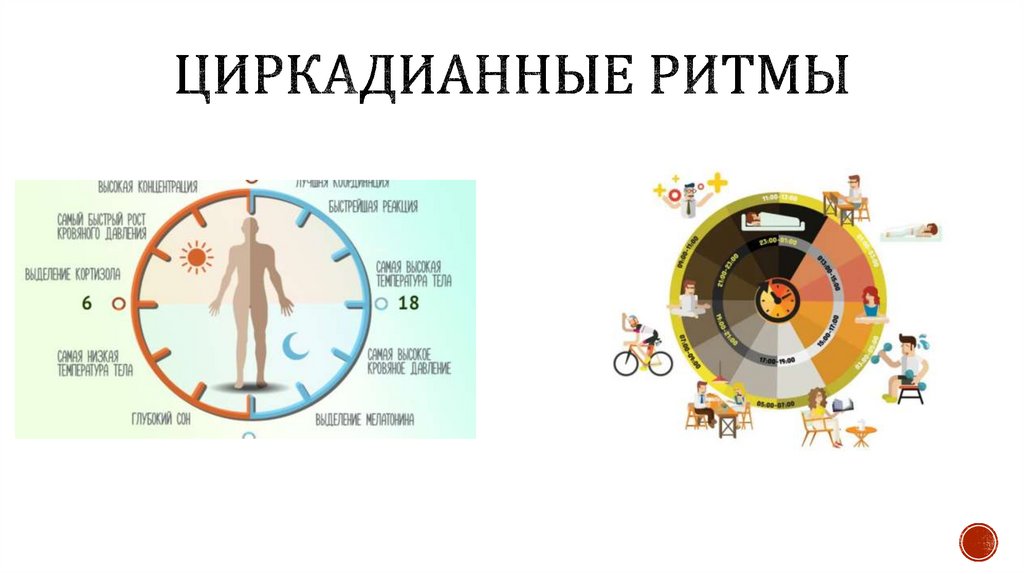 Так, цианобактерии сохраняют ритм обменных процессов близкий к 25 часам при вариациях темп-ры от 30 до 60 °C.
Так, цианобактерии сохраняют ритм обменных процессов близкий к 25 часам при вариациях темп-ры от 30 до 60 °C.
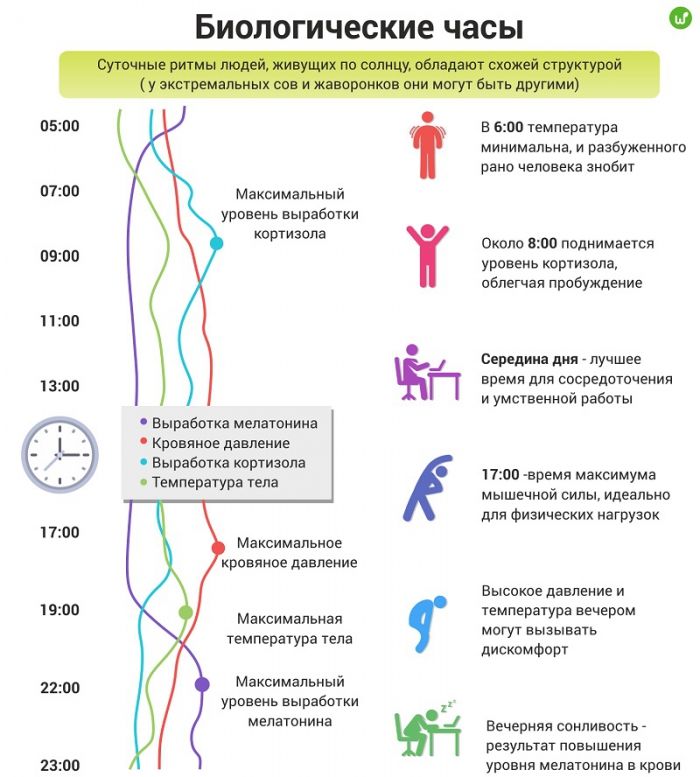
На основе внутриклеточных биологич. часов формируются циркадные ритмы отд. органов многоклеточного организма. Их ритмич. активность координируется центр. водителями ритма. У млекопитающих и человека роль центр. часов выполняют супрахиазматические ядра (СХЯ) гипоталамуса, которые связаны с вегетативными нервными ганглиями и эпифизом (рис. 1). Гормон эпифиза мелатонин синхронизирует экспрессию часовых генов в разл. отделах нейроэндокринной системы и совместно с нейромедиаторами и гормонами желёз внутренней секреции обеспечивает координацию ритмич. процессов, которая проявляется в упорядоченном чередовании подъёмов и спадов функциональной активности разл. систем организма. Так, перед пробуждением после сна происходит всплеск секреции гормонов коры надпочечников (кортизола и кортикостерона), играющих ключевую роль в энергетич.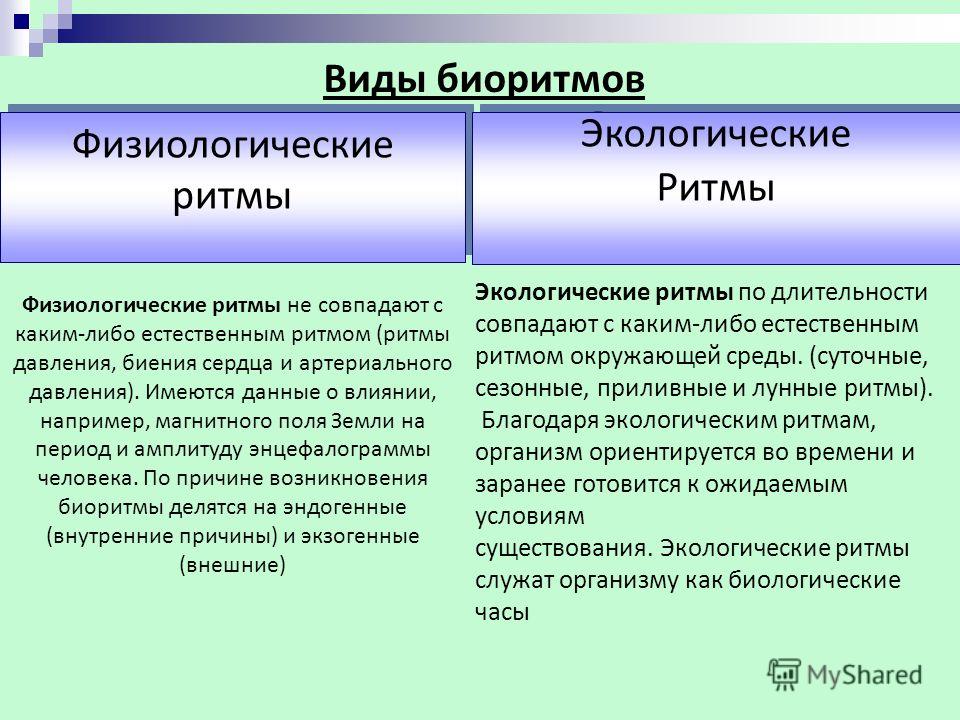 обеспечении физич. активности во время бодрствования, а увеличение секреции гормона роста, наблюдаемое в первой половине сна, стимулирует синтез белков, что необходимо для восстановления организма во время отдыха.
обеспечении физич. активности во время бодрствования, а увеличение секреции гормона роста, наблюдаемое в первой половине сна, стимулирует синтез белков, что необходимо для восстановления организма во время отдыха.
Рис. 2. У полярников в Антарктиде суточные ритмы температуры тела (А) и сила сердечных сокращений (В) реагируют на изменения фотопериода, а ритмы концентрации кортизола в слюне (Б) и частота пульса (Г…
Ход биологич. часов настраивается внешними циклич. процессами и, прежде всего, фотопериодом. У млекопитающих, включая человека, в фотопериодич. синхронизации Б. р. участвуют зрительные рецепторы, реагирующие на освещённость. Сигналы от них передаются к СХЯ гипоталамуса и далее к эпифизу. Свет подавляет секрецию мелатонина, который выделяется эпифизом исключительно в ночные часы, независимо от того, к какому времени суток приурочены сон или бодрствование конкретного вида животных. Благодаря влиянию на экспрессию генов биологич. часов мелатонин с одинаковым успехом синхронизирует циркадные ритмы ночных животных (мыши, крысы), у которых вечерний подъём активности совпадает с повышением концентрации гормона, и ритмы дневных видов (люди, обезьяны), у которых активность повышается утром при снижении его уровня.
Благодаря влиянию на экспрессию генов биологич. часов мелатонин с одинаковым успехом синхронизирует циркадные ритмы ночных животных (мыши, крысы), у которых вечерний подъём активности совпадает с повышением концентрации гормона, и ритмы дневных видов (люди, обезьяны), у которых активность повышается утром при снижении его уровня.
Мелатонин используется позвоночными животными и для синхронизации цирканнуальных ритмов жизнедеятельности, поскольку из всего многообразия сезонных феноменов именно изменения фотопериода являются наиболее надёжным индикатором времени года. При этом не столько количество мелатонина, сколько продолжительность его ночной секреции определяют сезонные морфофизиологич. преобразования, характер которых, в свою очередь, определяется образом жизни, свойственным определённому виду. Напр., осеннее увеличение ночной активности эпифиза стимулирует генеративную функцию у копытных, но подавляет её у грызунов.
Напр., осеннее увеличение ночной активности эпифиза стимулирует генеративную функцию у копытных, но подавляет её у грызунов.
Наряду с фотопериодом в синхронизации суточных ритмов участвуют и др. периодич. процессы, напр. у млекопитающих нарушение ритмов активности, вызванное разрушением СХЯ, восстанавливается при периодич. приёме пищи. Биологич. часы, координирующие работу разных органов, отличаются по способности к восприятию синхронизирующего влияния внешних «датчиков времени» разл. природы. В результате этого циркадная организация физиологич. процессов существенно меняется при быстром перемещении в новый часовой пояс, при сменной работе, при сезонных изменениях фотопериода, что особенно важно для жителей полярных районов (рис. 2). Наблюдаемые при этом отклонения в последовательности подъёмов и спадов Б.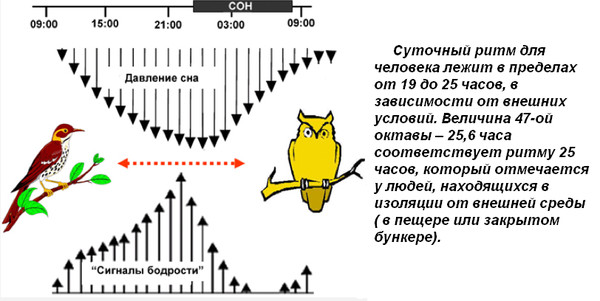 р. разл. систем организма (десинхроноз) отрицательно сказываются на работоспособности и сопротивляемости организма негативному влиянию разл. патогенных факторов. Наука, изучающая Б. р., – биоритмология, или хронобиология.
р. разл. систем организма (десинхроноз) отрицательно сказываются на работоспособности и сопротивляемости организма негативному влиянию разл. патогенных факторов. Наука, изучающая Б. р., – биоритмология, или хронобиология.
Развитие циркадных ритмов: от животных к человеку
1. Panda S, Hogenesch JB, Kay SA. Циркадные ритмы от мух до человека. Природа. 2002; 417 (6886): 329–35. [PubMed] [Google Scholar]
2. Ko CH, Takahashi JS. Молекулярные компоненты циркадных часов млекопитающих. Хум Мол Жене. 2006; 15 (2): R271–7. [PubMed] [Google Scholar]
3. Maywood ES, O’Neill J, Wong GK, Reddy AB, Hastings MH. Циркадный ритм здоровья и болезней. Прог Мозг Res. 2006; 153: 253–69.. [PubMed] [Google Scholar]
4. Moore-Ede MC, Czeisler CA, Richardson GS. Циркадный хронометраж здоровья и болезней. Часть 1. Основные свойства циркадных кардиостимуляторов. NEnglJMed. 1983;309(8):469–76. [PubMed] [Google Scholar]
5. Moore-Ede MC, Czeisler CA, Richardson GS. Циркадный хронометраж здоровья и болезней. Часть 2. Клинические проявления циркадной ритмичности. NEnglJMed. 1983;309(9):530–6. [PubMed] [Google Scholar]
Moore-Ede MC, Czeisler CA, Richardson GS. Циркадный хронометраж здоровья и болезней. Часть 2. Клинические проявления циркадной ритмичности. NEnglJMed. 1983;309(9):530–6. [PubMed] [Google Scholar]
6. Арендт Дж. Мелатонин и ритмы человека. Хронобиол Инт. 2006;23(12):21–37. [PubMed] [Академия Google]
7. Тофлер Г.Х., Мюллер Дж.Е. Запуск острого сердечно-сосудистого заболевания и потенциальные профилактические стратегии. Тираж. 2006; 114 (17): 1863–72. [PubMed] [Google Scholar]
8. Weaver DR. Супрахиазматическое ядро: 25-летняя ретроспектива. J Биол Ритмы. 1998;13(2):100–12. [PubMed] [Google Scholar]
9. Эбисава Т. Циркадные ритмы в ЦНС и расстройства периферических часов: расстройства сна человека и гены часов. J Pharmacol Sci. 2007 [PubMed] [Академия Google]
10. Maemura K, Takeda N, Nagai R. Циркадные ритмы в ЦНС и нарушениях периферических часов: роль биологических часов в сердечно-сосудистых заболеваниях. J Pharmacol Sci. 2007 [PubMed] [Google Scholar]
11. Одо С. Циркадные ритмы в ЦНС и расстройства периферических часов: хронофармакологические данные о противоопухолевых препаратах. J Pharmacol Sci. 2007 [PubMed] [Google Scholar]
Одо С. Циркадные ритмы в ЦНС и расстройства периферических часов: хронофармакологические данные о противоопухолевых препаратах. J Pharmacol Sci. 2007 [PubMed] [Google Scholar]
12. Welsh DK, Logothetis DE, Meister M, Reppert SM. Отдельные нейроны, диссоциированные от супрахиазматического ядра крысы, экспрессируют независимо фазированные циркадные ритмы возбуждения. Нейрон. 1995;14(4):697–706. [PubMed] [Google Scholar]
13. Liu C, Weaver DR, Strogatz SH, Reppert SM. Клеточная конструкция циркадных часов: определение периода в супрахиазматических ядрах. Клетка. 1997;91(6):855–60. [PubMed] [Google Scholar]
14. Morin LP. Циркадная зрительная система. Мозг ResBrain ResRev. 1994;19(1):102–27. [PubMed] [Google Scholar]
15. Morin LP, Allen CN. Циркадная зрительная система, 2005 г. Res Brain Res Rev. 2006; 51(1):1–60. [PubMed] [Академия Google]
16. Корнхаузер Дж.М., Майо К.Е., Такахаши Дж.С. Легкие, ранние гены и циркадные ритмы. Поведение Жене. 1996;26(3):221–40. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
17. Watts AG, Swanson LW, Sanchez-Watts G. Эфферентные проекции супрахиазматического ядра: I. Исследования с использованием антероградного транспорта лейкоагглютинина Phaseolus vulgaris у крыс. JCompNeurol. 1987;258(2):204–29. [PubMed] [Google Scholar]
18. Watts AG, Swanson LW. Эфферентные проекции супрахиазматического ядра: II. Исследования с использованием ретроградного транспорта флуоресцентных красителей и одновременной иммуногистохимии пептидов у крыс. JCompNeurol. 1987;258(2):230–52. [PubMed] [Google Scholar]
19. Мур Р.Ю. Организация циркадной системы приматов. JBiolRhythms. 1993; 8 (прил.): S3–9. С3–9. [PubMed] [Google Scholar]
20. Lydic R, Schoene WC, Czeisler CA, Moore-Ede MC. Супрахиазматическая область гипоталамуса человека: гомолог циркадного водителя ритма приматов? Спать. 1980;2(3):355–61. [PubMed] [Google Scholar]
21. Lydic R, Albers HE, Tepper B, Moore-Ede MC. Трехмерная структура супрахиазматических ядер млекопитающих: сравнительное исследование пяти видов. JCompNeurol. 1982;204(3):225–37. [PubMed] [Google Scholar]
JCompNeurol. 1982;204(3):225–37. [PubMed] [Google Scholar]
22. Лушингтон К., Галка Р., Сасси Л.Н., Кеннауэй Д.Дж., Доусон Д. Экстраокулярное воздействие света не сдвигает по фазе ритмы мелатонина слюны у спящих субъектов. J Биол Ритмы. 2002;17(4):377–86. [PubMed] [Google Scholar]
23. Schwartz WJ, Reppert SM, Eagan SM, Moore-Ede MC. Метаболическая активность супрахиазматических ядер in vivo: сравнительное исследование. Мозг Res. 1983; 274(1):184–7. [PubMed] [Google Scholar]
24. Ривкис С.А., Хофман П.Л., Фортман Дж. Новорожденные приматы увлекаются освещением низкой интенсивности. ProcNatlAcadSciUSA. 1997;94(1):292–7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Reppert SM, Perlow MJ, Ungerleider LG, et al. Влияние повреждения супрахиазматической области переднего гипоталамуса на суточные ритмы мелатонина и кортизола у макак-резусов. Дж. Нейроски. 1981; 1(12):1414–25. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Edgar DM, Dement WC, Fuller CA.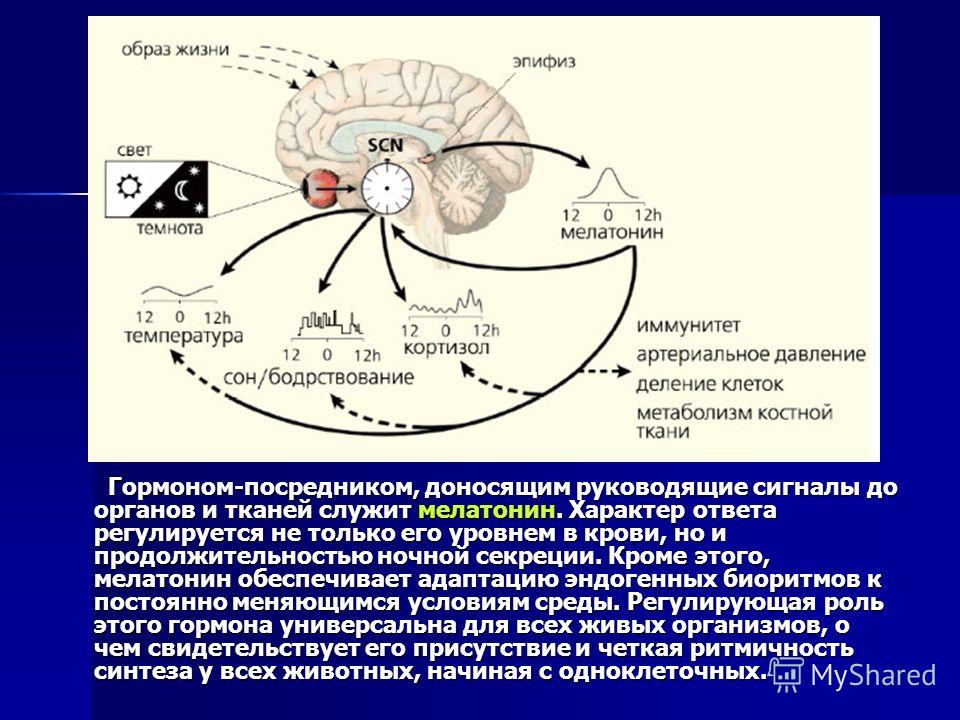 Влияние поражений СХЯ на сон у беличьих обезьян: свидетельство противоположных процессов в регуляции сна и бодрствования. Дж. Нейроски. 1993;13(3):1065–79. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Влияние поражений СХЯ на сон у беличьих обезьян: свидетельство противоположных процессов в регуляции сна и бодрствования. Дж. Нейроски. 1993;13(3):1065–79. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Schwartz WJ, Busis NA, Hedley-Whyte ET. Дискретное поражение вентрального гипоталамуса и перекреста зрительных нервов, нарушающее суточный температурный ритм. JNeurol. 1986; 233(1):1–4. [PubMed] [Google Scholar]
28. Rivkees S. Аритмичность у ребенка с септооптической дисплазией и установление цикличности сна-бодрствования мелатонином. J Педиатрия. 2001; 139: 463–5. [PubMed] [Google Scholar]
29. Фридман Д.И., Джонсон Дж.К., Чорский Р.Л., Стопа Э.Г. Мечение ретино-гипоталамического тракта человека карбоцианиновым красителем DiI. Мозг Res. 1991;560(12):297–302. [PubMed] [Google Scholar]
30. Садун А.А., Шехтер Д.Д., Смит Л.Е. Ретино-гипоталамический путь у человека: световое опосредование циркадианных ритмов. Мозг Res. 1984;302(2):371–377. [PubMed] [Google Scholar]
31. Кэмпбелл С.С., Мерфи П.Дж. Экстраокулярная циркадианная фототрансдукция у человека. Наука. 1998;279(5349):396–9. [PubMed] [Google Scholar]
Кэмпбелл С.С., Мерфи П.Дж. Экстраокулярная циркадианная фототрансдукция у человека. Наука. 1998;279(5349):396–9. [PubMed] [Google Scholar]
32. Hebert M, Martin SK, Eastman CI. Ночная секреция мелатонина не подавляется световым облучением за коленом у людей. Нейроски Летт. 1999;274(2):127–30. [PubMed] [Google Scholar]
33. Фостер Р.Г. Пролить свет на биологические часы. Нейрон. 1998;20(5):829–32. [PubMed] [Google Scholar]
34. Жан-Луи Г., Крипке Д.Ф., Коул Р.Дж., Эллиот Дж.А. Нет подавления мелатонина при освещении подколенных ямок или век. J Биол Ритмы. 2000;15(3):265–9. [PubMed] [Google Scholar]
35. Weitzman ED, Czeisler CA, Zimmerman JC, Moore-Ede MC. Биологические ритмы у человека: взаимосвязь сна и бодрствования, кортизола, гормона роста и температуры во время временной изоляции. Adv Biochem Psychopharmacol. 1981;28:475–99. [PubMed] [Google Scholar]
36. Czeisler CA, Klerman EB. Циркадная и зависимая от сна регуляция высвобождения гормонов у человека. Недавние прог.горм.разр. 1999; 54: 97–130. [PubMed] [Google Scholar]
Недавние прог.горм.разр. 1999; 54: 97–130. [PubMed] [Google Scholar]
37. Reppert SM, Weaver DR, Rivkees SA. Материнская связь циркадной фазы с развивающимся млекопитающим. Психонейроэндокринология. 1988;13(12):63–78. [PubMed] [Google Scholar]
38. Репперт С.М. Взаимодействие циркадных часов матери и плода. CibaFoundSymp. 1995;183:198–207. обсудить:198-207; обсуждение — [PubMed] [Google Scholar]
39. van Eerdenburg FJ, Rakic P. Ранний нейрогенез в переднем гипоталамусе макаки-резус. Мозг ResDevBrain Res. 1994;79(2):290–6. [PubMed] [Google Scholar]
40. Nowell-Morris L, Faherenbruch CE. Практические и эволюционные соображения по использованию модели нечеловеческих приматов в пренатальных исследованиях. Нью-Йорк: Алан Р. Лисс; 1985. [Google Scholar]
41. Reppert SM, Weaver DR, Rivkees SA, Stopa EG. Предполагаемые рецепторы мелатонина в биологических часах человека. Наука. 1988;242(4875):78–81. [PubMed] [Google Scholar]
42. Rivkees SA, Lachowicz JE. Функциональные дофаминовые рецепторы D1 и D5 экспрессируются в супрахиазматических, супраоптических и паравентрикулярных ядрах приматов. Синапс. 1997;26(1):1–10. [PubMed] [Google Scholar]
Функциональные дофаминовые рецепторы D1 и D5 экспрессируются в супрахиазматических, супраоптических и паравентрикулярных ядрах приматов. Синапс. 1997;26(1):1–10. [PubMed] [Google Scholar]
43. Reppert SM, Schwartz WJ. Функциональная активность супрахиазматических ядер у плода примата. NeurosciLett. 1984;46(2):145–9. [PubMed] [Google Scholar]
44. Джолли А. Час рождения у приматов и человека. Фолиа Приматол. 1972;18(1):108–21. [PubMed] [Google Scholar]
45. Swaab DF. Развитие гипоталамуса человека. Нейрохим Рез. 1995;20(5):509–19. [PubMed] [Google Scholar]
46. Glotzbach SF, Sollars P, Pagano M. Развитие ретино-гипоталамического тракта человека. Соц Неврологии. 1992;18:857. [Google Scholar]
47. Rivkees SA, Hao H. Развитие циркадной ритмичности. Семин Перинатол. 2000;24(4):232–42. [PubMed] [Google Scholar]
48. Boivin DB, Duffy JF, Kronauer RE, Czeisler CA. Зависимость доза-реакция для сброса циркадных часов человека светом. Природа. 1996;379(6565):540–2.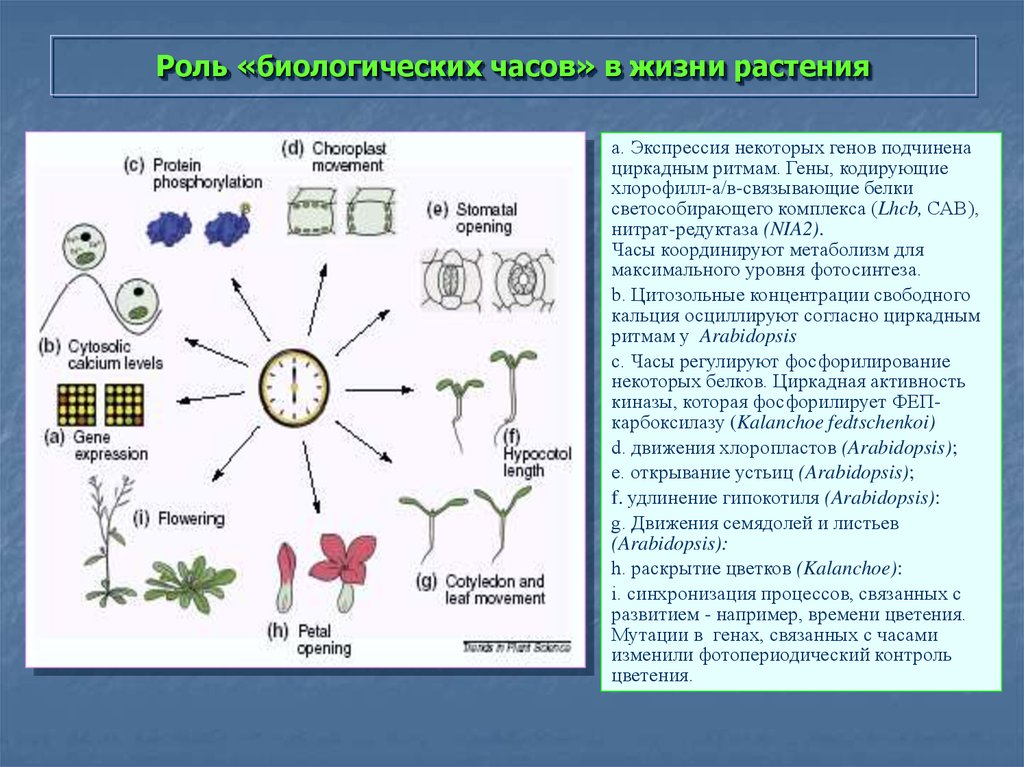 [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
49. Shanahan TL, Czeisler CA. Физиологическое воздействие света на циркадный водитель ритма человека. Семин Перинатол. 2000;24(4):299–320. [PubMed] [Google Scholar]
50. Hao H, Rivkees SA. Биологические часы очень недоношенных детенышей приматов реагируют на свет. Proc Natl Acad Sci U S A. 1999;96(5):2426–9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Seron-Ferre M, Ducsay CA, Valenzuela GJ. Циркадные ритмы во время беременности. EndocrRev. 1993;14(5):594–609. [PubMed] [Google Scholar]
52. Серон-Ферре М., Торрес-Фарфан С., Форселледо М.Л., Валенсуэла Г.Дж. Развитие циркадных ритмов у плода и новорожденного. Семин Перинатол. 2001;25(6):363–70. [PubMed] [Google Scholar]
53. Parmelee AH, Jr Циклы сна у младенцев. DevMedChild Нейрол. 1969; 11 (6): 794–5. [PubMed] [Google Scholar]
54. Meier-Koll A, Hall U, Hellwig U, Kott G, Meier-Koll V. Система биологических осцилляторов и развитие поведения сна и бодрствования в раннем младенчестве. Хронобиология. 1978;5(4):425–40. [PubMed] [Google Scholar]
Хронобиология. 1978;5(4):425–40. [PubMed] [Google Scholar]
55. Kleitman J, Engelman Характеристики сна младенцев. J Appl Физиолол. 1953; 6: 127–34. [Google Scholar]
56. Darnall RA, Ariagno RL, Kinney HC. Поздний недоношенный ребенок и контроль дыхания, сна и развития ствола мозга: обзор. Клин Перинатол. 2006;33(4):883–914. абстракция х. [PubMed] [Google Scholar]
57. Дженни О.Г., Дебур Т., Акерманн П. Развитие 24-часового режима покоя-активности у младенцев. Младенец Поведение Дев. 2006;29(2): 143–52. [PubMed] [Google Scholar]
58. McKay SM, Angulo-Barroso RM. Продольная оценка двигательной активности ног и характера сна у младенцев с синдромом Дауна и без него. Младенец Поведение Дев. 2006;29(2):153–68. [PubMed] [Google Scholar]
59. Kennaway DJ, Stamp GE, Goble FC. Развитие выработки мелатонина у младенцев и влияние недоношенности. JClinEndocrinolMetab. 1992;75(2):367–9. [PubMed] [Google Scholar]
60. Kennaway DJ, Goble FC, Stamp GE. Факторы, влияющие на развитие ритмичности мелатонина у человека.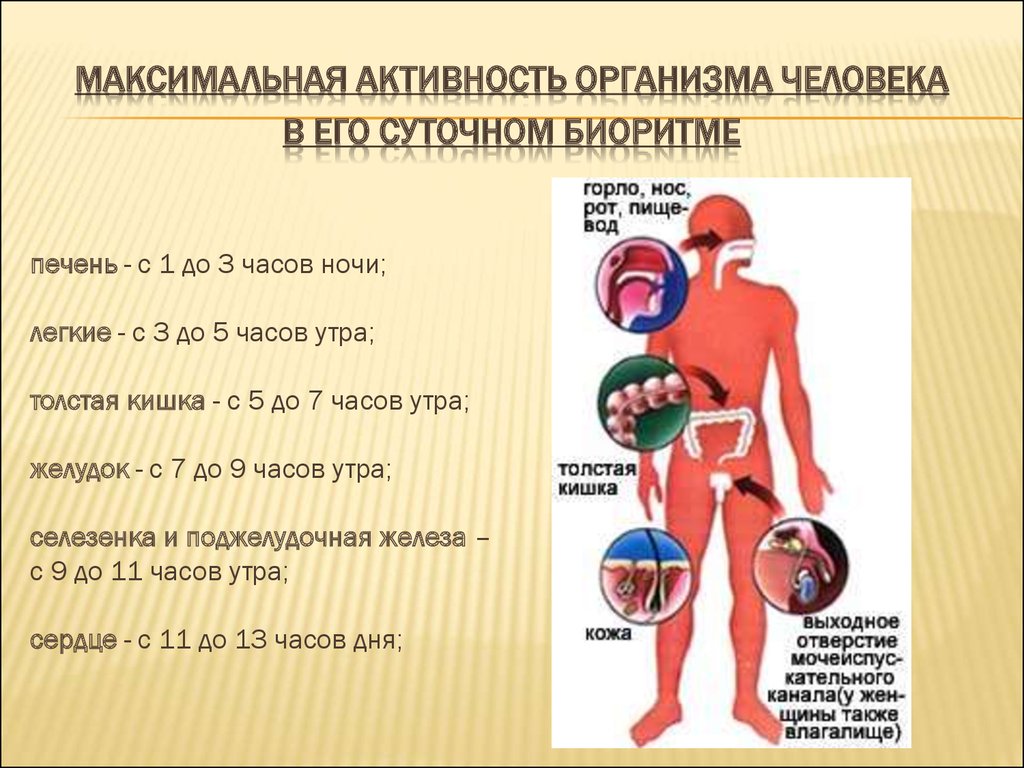 JClinEndocrinolMetab. 1996;81(4):1525–32. [PubMed] [Google Scholar]
JClinEndocrinolMetab. 1996;81(4):1525–32. [PubMed] [Google Scholar]
61. Beitins IZ, Kowarski A, Migeon CJ, Graham GG. Функция надпочечников у нормальных детей, а также при маразме и квашиоркоре. Секреция кортизола, суточные колебания уровня кортизола в плазме и экскреция с мочой 17-гидроксикортикоидов, свободных кортикоидов и кортизола. J Педиатр. 1975; 86 (2): 302–8. [PubMed] [Google Scholar]
62. Onishi S, Miyazawa G, Nishimura Y, et al. Постнатальное развитие циркадных ритмов уровня кортизола в сыворотке крови у детей. Педиатрия. 1983;72(3):399–404. [PubMed] [Google Scholar]
63. Price DA, Close GC, Fielding BA. Возраст появления циркадного ритма в значениях кортизола слюны в младенчестве. ArchDisChild. 1983; 58 (6): 454–6. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Hellbrugge T, Lange JE, Rutenfranz J. Суточная периодичность физиологических функций на разных этапах младенчества и детства. Энн NY Acad Sci. 1964; 117: 361–73. [PubMed] [Google Scholar]
65. Parmelee AH, Jr, Wenner WH, Akiyama Y, Schultz M, Stern E. Состояния сна у недоношенных детей. DevMedChild Нейрол. 1967;9(1):70–7. [PubMed] [Google Scholar]
Parmelee AH, Jr, Wenner WH, Akiyama Y, Schultz M, Stern E. Состояния сна у недоношенных детей. DevMedChild Нейрол. 1967;9(1):70–7. [PubMed] [Google Scholar]
66. Rivkees SA. Развитие циркадных ритмов. Основные и клинические аспекты. PediatrClinNorth Am. 1997;44(2):467–87. [PubMed] [Google Scholar]
67. Д’Суза С.В., Тенрейро С., Минорс Д., Чизвик М.Л., Симс Д.Г., Уотерхаус Дж. Температура кожи и ритмы сердечного ритма у глубоко недоношенных детей. ArchDisChild. 1992;67(7):784–8. [PMC free article] [PubMed] [Google Scholar]
68. Андерс Т.Ф., Кинер М.А., Кремер Х. Организация состояния сна-бодрствования, неонатальная оценка и развитие недоношенных детей в течение первого года жизни. II. Спать. 1985;8(3):193–206. [PubMed] [Google Scholar]
69. Glotzbach SF, Edgar DM, Boeddiker M, Ariagno RL. Биологическая ритмичность у нормальных детей первых 3 мес жизни. Педиатрия. 1994; 94 (4 части 1): 482–8. [PubMed] [Google Scholar]
70. Glotzbach SF, Edgar DM, Ariagno RL. Биологическая ритмичность у недоношенных детей до выписки из реанимации новорожденных. Педиатрия. 1995;95(2):231–237. [PubMed] [Google Scholar]
Биологическая ритмичность у недоношенных детей до выписки из реанимации новорожденных. Педиатрия. 1995;95(2):231–237. [PubMed] [Google Scholar]
71. Rivkees SA, Mayes L, Jacobs H, Gross I. Режим покоя и активности недоношенных детей регулируется циклическим освещением. Педиатрия. 2004; 113 в печати. [PubMed] [Академия Google]
72. Мирмиран М., Арианьо Р.Л. Влияние света в отделении интенсивной терапии новорожденных на развитие циркадных ритмов у недоношенных детей. Семин Перинатол. 2000;24(4):247–57. [PubMed] [Google Scholar]
73. Манн Н.П., Хэддоу Р., Стоукс Л., Гудли С., Раттер Н. Влияние дня и ночи на недоношенных детей в отделении для новорожденных: рандомизированное исследование. Br Med J (Clin Res Ed) 1986;293(6557):1265–7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
74. Brandon DH, Holditch-Davis D, Belyea M. У недоношенных детей, рожденных на сроке менее 31 недели гестации, наблюдается улучшение роста при циклическом освещении по сравнению с непрерывной полной темнотой. J Педиатр. 2002;140(2):192–9. [PubMed] [Google Scholar]
J Педиатр. 2002;140(2):192–9. [PubMed] [Google Scholar]
75. Мирмиран М., Лунсхоф С. Перинатальное развитие циркадных ритмов человека. ProgBrain Res. 1996; 111: 217–26. 217–26. [PubMed] [Google Scholar]
76. McGraw K, Hoffmann R, Harker C, Herman JH. Развитие циркадных ритмов у младенцев. Спать. 1999;22(3):303–10. [PubMed] [Google Scholar]
77. Арианьо Р.Л., Мирмиран М. Проливая свет на младенцев с очень низкой массой тела при рождении. J Педиатр. 2001;139(4):476–7. [PubMed] [Академия Google]
78. Гласс П., Эйвери Г.Б., Субраманиан К.Н., Киз М.П., Состек А.М., Френдли Д.С. Влияние яркого света в родильном доме на заболеваемость ретинопатией недоношенных. NEnglJMed. 1985;313(7):401–4. [PubMed] [Google Scholar]
79. Fielder AR, Moseley MJ. Окружающий свет и недоношенный ребенок. Семин Перинатол. 2000;24(4):291–8. [PubMed] [Google Scholar]
80. Рейнольдс Дж. Д., Харди Р. Дж., Кеннеди К. А., Спенсер Р., ван Хойвен В. А., Филдер А. Р. Недостаточная эффективность снижения освещенности в профилактике ретинопатии недоношенных.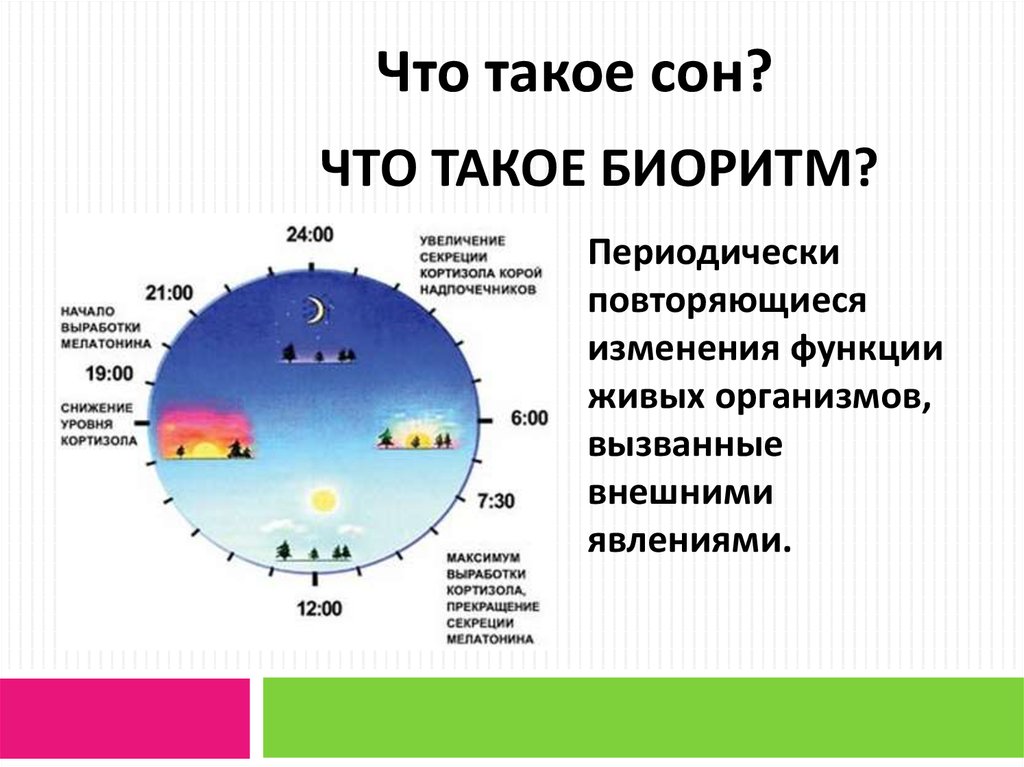 Кооперативная группа по уменьшению света при ретинопатии недоношенных (LIGHT-ROP). NEnglJMed. 1998;338(22):1572–156. [PubMed] [Google Scholar]
Кооперативная группа по уменьшению света при ретинопатии недоношенных (LIGHT-ROP). NEnglJMed. 1998;338(22):1572–156. [PubMed] [Google Scholar]
81. Кеннеди К.А., Филдер А.Р., Харди Р.Дж., Танг Б., Гордон Д.К., Рейнольдс Д.Д. Уменьшенное освещение не улучшает медицинские результаты у младенцев с очень низкой массой тела при рождении. J Педиатр. 2001;139(4):527–31. [PubMed] [Google Scholar]
82. Элс Х., Лоухон Дж., Даффи Ф.Х., Маканулти ГБ, Гибс-Гроссман Р., Бликман Дж.Г. Индивидуальный уход за недоношенным ребенком с очень низкой массой тела при рождении. Медицинские и нейрофункциональные эффекты. Джама. 1994;272(11):853–8. [PubMed] [Академия Google]
83. Ariagno RL, Thoman EB, Boeddiker MA, et al. Уход за развитием не влияет на сон и развитие недоношенных детей. Педиатрия. 1997;100(6):Е9. [PubMed] [Google Scholar]
84. Symington A, Pinelli J. Уход за развитием для содействия развитию и предотвращения заболеваемости у недоношенных детей. Cochrane Database Syst Rev. 2006;(2):CD001814. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2006;(2):CD001814. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Циркадные ритмы животных | Психология Вики
Оценка |
Биопсихология |
Сравнительный |
Познавательный |
Развивающие |
Язык |
Индивидуальные различия |
Личность |
Философия |
Социальные |
Методы |
Статистика |
Клинический |
Образовательные |
промышленный |
Профессиональные товары |
Мировая психология |
Животные · Этология животных · Сравнительная психология · Модели животных · Контур · Индекс
- Информацию о циркадных ритмах у людей см. в: Биологические ритмы человека
Циркадный ритм представляет собой приблизительно 24-часовой цикл физиологических процессов у животных. Термин «циркадный», введенный Францем Хальбергом, [1] происходит от латинского circa , «около», а dies , «день», что буквально означает «около дня». Формальное изучение биологических временных ритмов, таких как суточные, недельные, сезонные и годовые ритмы, называется хронобиологией.
Строго говоря, циркадные ритмы генерируются эндогенно, хотя они могут модулироваться внешними сигналами, такими как солнечный свет и температура.
Содержание
- 1 История
- 2 Критерий
- 3 Происхождение
- 4 Циркадные ритмы животных
- 4.1 Влияние цикла свет-темнота
- 4.2 Супрахиазматическое ядро
- 4.3 Вне SCN «главные часы»
- 4.4 Разрушение
- 4.5 Связь с кокаином
- 5 Свет и биологические часы
- 6 См. также
- 7 Примечания
- 8 Дополнительная литература
- 9 Внешние ссылки
- 10 См. также
- 11 Ссылки и библиография
- 12 ключевых текстов
- 12.1 Книги
- 12.2 Бумаги
- 13 Дополнительный материал
- 13.1 Книги
- 13.2 Бумаги
- 14 Внешние ссылки
История
Первое эндогенное циркадное колебание наблюдал в 1700-х годах французский ученый Жан-Жак д’Орту де Меран, который заметил, что 24-часовые закономерности в движении листьев растения Mimosa pudica сохранялись, даже когда растения изолировали от внешних раздражителей.
Самое раннее известное описание циркадного ритма датируется четвертым веком до нашей эры, когда Андростенес в описании походов Александра Македонского описал суточные движения листьев тамариндового дерева.
Критерии
Общие критерии циркадных ритмов
- Ритм сохраняется в постоянных условиях (например, при постоянной темноте) с периодом около 24 часов: Обоснование этого критерия состоит в том, чтобы отличить циркадные ритмы от тех «кажущихся ритмов», которые просто реагируют на внешние периодические сигналы. Например, вы бы не хотели, чтобы поведение «носит солнцезащитные очки» классифицировалось как циркадный ритм — если бы не было солнечного света, поведение не сохранялось бы.
- Ритм имеет один и тот же период в диапазоне температур (т. е. он компенсирован по температуре): Обоснование этого критерия состоит в том, чтобы отличить циркадные ритмы от других биологических ритмов, возникающих из-за кругового характера пути реакции, например цикла Кребса в метаболизме.
 . При достаточно низкой или достаточно высокой температуре период круговой реакции может достигать 24 часов, но это будет просто совпадение, а не намерение.
. При достаточно низкой или достаточно высокой температуре период круговой реакции может достигать 24 часов, но это будет просто совпадение, а не намерение. - Ритм может быть сброшен при воздействии внешнего раздражителя: обоснование этого критерия состоит в том, чтобы отличить циркадные ритмы от других вообразимых эндогенных 24-часовых ритмов, которые невосприимчивы к сбросу внешними сигналами и, следовательно, не служат цели оценки местного времени. . Ваши наручные часы не имеют возможности определять местное время, но ваши биологические часы могут.
 . При достаточно низкой или достаточно высокой температуре период круговой реакции может достигать 24 часов, но это будет просто совпадение, а не намерение.
. При достаточно низкой или достаточно высокой температуре период круговой реакции может достигать 24 часов, но это будет просто совпадение, а не намерение.Происхождение
Считается, что циркадные ритмы возникли в самых ранних клетках с целью защиты реплицирующейся ДНК от сильного ультрафиолетового излучения в дневное время. В результате репликация была переведена в темноту. Грибок Neurospora , существующий сегодня, сохраняет этот регулируемый часами механизм.
Простейшими известными циркадными часами являются часы прокариотических цианобактерий.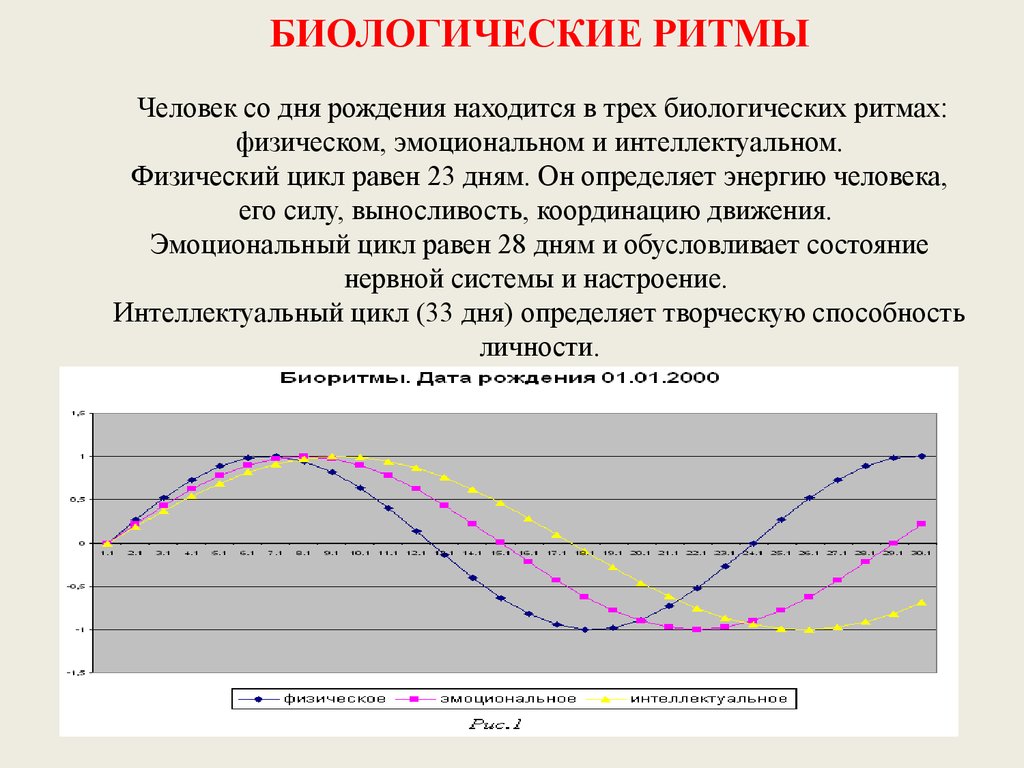 Недавние исследования показали, что циркадные часы Synechococcus elongatus может быть воссоздан in vitro всего тремя белками их центрального осциллятора. Было показано, что эти часы поддерживают 22-часовой ритм в течение нескольких дней после добавления АТФ. Предыдущие объяснения прокариотического циркадного хронометриста зависели от механизма обратной связи транскрипции / трансляции ДНК, и, хотя это не было доказано, по-прежнему считается, что это справедливо для эукариотических организмов.
Действительно, хотя циркадианные системы эукариот и прокариот имеют одинаковую базовую архитектуру: вход — центральный осциллятор — выход, они не имеют никакой гомологии. Это предполагает вероятное независимое происхождение.
Недавние исследования показали, что циркадные часы Synechococcus elongatus может быть воссоздан in vitro всего тремя белками их центрального осциллятора. Было показано, что эти часы поддерживают 22-часовой ритм в течение нескольких дней после добавления АТФ. Предыдущие объяснения прокариотического циркадного хронометриста зависели от механизма обратной связи транскрипции / трансляции ДНК, и, хотя это не было доказано, по-прежнему считается, что это справедливо для эукариотических организмов.
Действительно, хотя циркадианные системы эукариот и прокариот имеют одинаковую базовую архитектуру: вход — центральный осциллятор — выход, они не имеют никакой гомологии. Это предполагает вероятное независимое происхождение.
В 1971 году Конопка и Бенцер впервые определили генетический компонент биологических часов, используя плодовую мушку в качестве модельной системы. Три мутантные линии мух демонстрировали аберрантное поведение: у одной период был короче, у другой — длиннее, а у третьей не было. Все три мутации сопоставлены с одним и тем же геном и окрещены периодом . Тот же самый ген был идентифицирован как дефектный при расстройстве сна под названием FASPS (семейный синдром продвинутой фазы сна) у людей тридцать лет спустя, что подчеркивает законсервированную природу молекулярных циркадных часов в ходе эволюции. Теперь мы знаем гораздо больше генетических компонентов биологических часов. Их взаимодействие приводит к замкнутой петле обратной связи генных продуктов, что приводит к периодическим колебаниям, которые клетки тела интерпретируют как определенное время дня.
Все три мутации сопоставлены с одним и тем же геном и окрещены периодом . Тот же самый ген был идентифицирован как дефектный при расстройстве сна под названием FASPS (семейный синдром продвинутой фазы сна) у людей тридцать лет спустя, что подчеркивает законсервированную природу молекулярных циркадных часов в ходе эволюции. Теперь мы знаем гораздо больше генетических компонентов биологических часов. Их взаимодействие приводит к замкнутой петле обратной связи генных продуктов, что приводит к периодическим колебаниям, которые клетки тела интерпретируют как определенное время дня.
Наше понимание биологических часов прошло долгий путь от «Представьте, что это генератор синусоидальных волн». Теперь мы знаем, что молекулярные циркадные часы могут функционировать в пределах одной клетки, то есть они клеточно автономны. В то же время разные ячейки могут общаться друг с другом, что приводит к синхронизированному и демократичному выходу электрических сигналов. Они могут взаимодействовать с эндокринными железами головного мозга, что приводит к периодическому выбросу гормонов. Рецепторы этих гормонов могут располагаться далеко по всему телу и синхронизировать периферические часы (например, те, которые присутствуют в печени). Таким образом, информация о времени суток, определяемая глазами, может передаваться часам в мозгу, и благодаря этому часы в остальной части тела могут быть синхронизированы. Это то, как многие виды поведения, такие как питье воды, сон/бодрствование и температура тела, координировано контролируются биологическими часами.
Рецепторы этих гормонов могут располагаться далеко по всему телу и синхронизировать периферические часы (например, те, которые присутствуют в печени). Таким образом, информация о времени суток, определяемая глазами, может передаваться часам в мозгу, и благодаря этому часы в остальной части тела могут быть синхронизированы. Это то, как многие виды поведения, такие как питье воды, сон/бодрствование и температура тела, координировано контролируются биологическими часами.
Циркадные ритмы животных
Суточные ритмы играют важную роль в определении режима сна и питания всех животных, включая людей. Существуют четкие закономерности активности мозговых волн, выработки гормонов, регенерации клеток и других биологических процессов, связанных с этим суточным циклом.
Суточные ритмы также играют роль в ретикулярной активирующей системе ретикулярной формации.
Влияние цикла свет-темнота
Ритм связан с циклом свет-темнота. Животные, которых долгое время держали в полной темноте, в конечном итоге функционируют в «свободном» ритме.
Свободноживущие организмы по-прежнему имеют консолидированный цикл сна-бодрствования, когда находятся в среде, защищенной от внешних сигналов, но этот ритм не укоренился и может не совпадать по фазе с другими циркадными или ультрадианными ритмами, такими как температура и пищеварение. Это исследование повлияло на дизайн космических кораблей, поскольку было обнаружено, что системы, имитирующие цикл свет/темнота, очень полезны для астронавтов.
Супрахиазматическое ядро
Суточные «часы» у млекопитающих расположены в супрахиазматическом ядре (СХЯ), отдельной группе клеток, расположенных в гипоталамусе.
Похоже, что СХЯ получает информацию о продолжительности дня от сетчатки, интерпретирует ее и передает в шишковидную железу (горохоподобная структура, обнаруженная на эпиталамусе), которая затем в ответ выделяет гормон мелатонин. Секреция мелатонина достигает максимума ночью и снижается в течение дня.
Вне «главных часов» СХЯ
Недавно появились доказательства того, что циркадные ритмы обнаруживаются во многих клетках тела за пределами «главных часов» СХЯ. Кроме того, клетки печени, например, реагируют на питание, а не на свет. Клетки из многих частей тела, по-видимому, имеют «свободные» ритмы.
Клетки тела, имеющие собственные ритмы, называются периферическими осцилляторами. Эти ткани включают пищевод, легкие, печень, селезенку и тимус. Есть некоторые свидетельства того, что обонятельная луковица и простата также могут испытывать колебания при культивировании, что позволяет предположить, что эти структуры также могут быть слабыми осцилляторами.
Нарушение ритма
Нарушение ритма обычно имеет краткосрочные негативные последствия. Многие путешественники испытали состояние, известное как синдром смены часовых поясов, с сопутствующими ему симптомами усталости, дезориентации и бессонницы. Ряд других расстройств, например биполярное расстройство и некоторые нарушения сна, связаны с нерегулярным или патологическим функционированием циркадианных ритмов. Недавние исследования показывают, что на нарушения циркадного ритма, обнаруживаемые при биполярном расстройстве, оказывает положительное влияние литий на часовые гены.
Нарушение ритмов в долгосрочной перспективе, как полагают, имеет серьезные неблагоприятные последствия для здоровья периферических органов за пределами головного мозга, особенно при развитии или обострении сердечно-сосудистых заболеваний. Выбор времени лечения в соответствии с биологическими часами может значительно повысить эффективность и снизить токсичность препарата или побочные реакции. Например, своевременное лечение ингибиторами ангиотензинпревращающего фермента (АПФ) может снизить ночное артериальное давление, а также улучшить (обратное) ремоделирование левого желудочка.
Связь с кокаином
Кроме того, циркадные ритмы и часовые гены, экспрессируемые в областях мозга за пределами СХЯ, могут значительно влиять на эффекты, вызываемые такими наркотиками, как кокаин. [3] [4]
Более того, генетические манипуляции с часовыми генами сильно влияют на действие кокаина. [5]
Свет и биологические часы
Способность света сбрасывать биологические часы зависит от кривой фазового отклика (на свет). В зависимости от фазы сна свет может ускорять или замедлять циркадный ритм. Требуемая освещенность варьируется от вида к виду, для сброса часов у ночных грызунов требуется гораздо более низкий уровень освещенности, чем у людей.
В дополнение к интенсивности света длина волны (или цвет) света является важным фактором, определяющим степень переустановки часов. Наиболее эффективно меланопсин возбуждается синим светом (420-440 нм). [6]
См. также
- Актиграфия
- Синдром продвинутой фазы сна (ASPS) и семейный синдром продвинутой фазы сна (FASPS)
- Биологические ритмы животных
- Собор
- Сумеречный
- Дневной
- Метаповоротный
- Ночь
- Нарушения циркадного ритма сна
- Биологические ритмы человека
Примечания
- ↑ http://www.msi.umn.edu/~halberg Центр хронобиологии Халберга
- ↑ http://www.nimh.nih.gov/press/lithiumenzyme. cfm
- ↑ Уз Т., Ахисароглу М., Ахмед Р., Манев Х. (2003). Шишковидная железа имеет решающее значение для циркадной экспрессии Period1 в полосатом теле и для циркадной сенсибилизации кокаином у мышей. Нейропсихофармакология 28 (12): 2117-23.
- ↑ Куртунку М., Арслан А., Ахисароглу М., Манев Х., Уз Т. (2004). Участие шишковидной железы в дневном кокаиновом вознаграждении у мышей. Eur J Pharmacol 489 (3): 203-5.
- ↑ МакКланг С., Сидиропулу К., Витатерна М., Такахаши Дж., Уайт Ф., Купер Д., Нестлер Э. (2005). Регуляция дофаминергической передачи и кокаинового вознаграждения геном Clock. Proc Natl Acad Sci U S A 102 (26): 9377-81.
- ↑ Ньюман Л.А., Уокер М.Т., Браун Р.Л., Кронин Т.В., Робинсон П.Р.: «Меланопсин образует функциональный коротковолновый фотопигмент», Биохимия. 11 ноября 2003 г .; 42 (44): 12734-8.
Дополнительная литература
- Ашофф Дж. (ред.) (1965) Циркадные часы . North Holland Press, Амстердам
- Авиви А., Альбрехт У., Остер Х., Джоэл А., Бейлес А., Нево Е. 2001. Биологические часы в полной темноте: циркадианная система Clock/MOP3 слепого подземного землекопа. Proc Natl Acad Sci USA 98:13751- 13756.
- Авиви А., Остер Х., Джоэл А., Бейлес А., Альбрехт У., Нево Е. 2002. Циркадные гены у слепого подземного млекопитающего II: сохранение и уникальность трех гомологов периода у слепого подземного землекопа, Spalax ehrenbergi надвидов. Proc Natl Acad Sci USA 99:11718-11723.
- Ditty JL, Williams SB, Golden SS (2003) Цианобактериальный циркадный механизм синхронизации. Анну Рев Жене 37:513-543
- Данлэп Дж. К., Лорос Дж., ДеКурси П. Дж. (2003) Хронобиология: Биохронометрия . Синауэр, Сандерленд
- Дворник В., Виноградова О.Н., Нево Е. (2003) Происхождение и эволюция генов циркадных часов у прокариот. Proc Natl Acad Sci USA 100:2495-2500
- Koukkari WL, Southern RB (2006) Знакомство с биологическими ритмами . Спрингер, Нью-Йорк
- Мартино Т., Араб С., Страуме М., Белшам Д.Д., Тата Н., Кай Ф., Лю П., Тривьери М., Ральф М., Соле М.Дж. Дневные/ночные ритмы в экспрессии генов нормального мышиного сердца. Дж. Мол Мед. 2004 г., апрель; 82 (4): 256–64. Epub 2004, 24 февраля. PMID: 14985853
- Refinetti R (2006) Циркадная физиология, 2-е изд. . CRC Press, Бока-Ратон
- Takahashi JS, Zatz M (1982) Регуляция циркадной ритмичности. Наука 217:1104–1111
- Tomita J, Nakajima M, Kondo T, Iwasaki H (2005) Отсутствие обратной связи транскрипция-трансляция в циркадном ритме фосфорилирования KaiC. Наука 307: 251–254
Внешние ссылки
- www.circadian.org
- Биологические часы Описание циркадных ритмов растений де Майраном, Линнеем и Дарвином
- 2006-01-18 Центр медицинских наук Техасского университета в Хьюстоне Темнота представляет жизненно важный метаболический топливный переключатель между сахаром и жиром
- Искусственный свет сбрасывает естественные часы
- Циркадные часы идут in vitro: чисто посттрансляционные осцилляторы у цианобактерий
- Пора спать, сказали часы надпочечников
- Джетлаг, циркадные часы
См.