Локализация Высших Психических Функций — это… Что такое Локализация Высших Психических Функций?
- Локализация Высших Психических Функций
- Локализация высших психических функций — теоретический концепт, выражающий собой более или менее однозначное соответствие определенных участков мозга и психических процессов. Проблема связи психики — и мозга ставилась еще в труд
Психологический словарь. 2000.
- Лозоискательство
- Локк Дж. Отрывки
Смотреть что такое «Локализация Высших Психических Функций» в других словарях:
локализация высших психических функций — (от лат. localis местный) отнесение высших психических функций к конкретным мозговым структурам (см. мозг). Проблема Л. в. п. ф. разрабатывается нейропсихологией, нейроанатомией, нейрофизиологией и др.
 История изучения Л. в. п. ф. восходит к… … Большая психологическая энциклопедия
История изучения Л. в. п. ф. восходит к… … Большая психологическая энциклопедияЛокализация высших психических функций — Исторически сложилась в дискуссиях между локализационизмом и эквипотенциализмом. В советской нейропсихологии концепция Л.в.п.ф. разработана А.Р. Лурия и его школой. Базируется на понимании психических функций как сложных системных образований, а… … Толковый словарь психиатрических терминов
Локализация высших психических функций динамическая — (лат. localis греч. dynamikos относящийся к силе, сильный) теория А.Р.Лурия, согласно которой материальным субстратом психологических функций являются сложные нейропсихологические структуры, образованные путём системной интеграции активности… … Энциклопедический словарь по психологии и педагогике
ДИНАМИЧЕСКАЯ ЛОКАЛИЗАЦИЯ ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ — множество высокоспециализированных отделов мозга (субстрата психических функций, работающего как единое целое), каждый из которых выполняет свою специфическую роль, обеспечивает не всю психическую функцию и даже не отдельные ее звенья, а те… … Психомоторика: cловарь-справочник
восстановление высших психических функций — (восстановление высших психических функций) раздел нейропсихологии, посвященный изучению механизмов и методов восстановления функций психических высших, нарушенных вследствие поражений локальных мозга головного.
На базе представлений об… … Большая психологическая энциклопедияТЕОРИЯ СИСТЕМНОЙ ДИНАМИЧЕСКОЙ ЛОКАЛИЗАЦИИ ВЫСШИХ ПСИХИЧЕСКИХ ФУНКЦИЙ — см. Локализация высших психических функций, Лурия А. Р. Большой психологический словарь. М.: Прайм ЕВРОЗНАК. Под ред. Б.Г. Мещерякова, акад. В.П. Зинченко … Большая психологическая энциклопедия
локализация функций в коре больших полушарий
ЛОКАЛИЗАЦИЯ ФУНКЦИЙ ДИНАМИЧЕСКАЯ — согласно теории системной динамической локализации высших психических функций мозг рассматривается как субстрат, состоящий из дифференцированных по своим функциям отделов и работающий как еди ное целое … Психомоторика: cловарь-справочник
функция психическая высшая: локализация — отнесение функций психических высших к конкретным мозговым структурам.
Представители узкого локализационизма рассматривали психологические функции как единые, неразложимые на компоненты «психические способности», реализуемые ограниченными… … Большая психологическая энциклопедиянейропсихология — отрасль психологической науки, сложившаяся на стыке психологии, медицины (неврологии, нейрохирургии) и физиологии, изучающая мозговые механизмы высших психических функций на материале локальных поражений головного мозга (см. компенсация… … Большая психологическая энциклопедия
 История изучения Л. в. п. ф. восходит к… … Большая психологическая энциклопедия
История изучения Л. в. п. ф. восходит к… … Большая психологическая энциклопедия На базе представлений об… … Большая психологическая энциклопедия
На базе представлений об… … Большая психологическая энциклопедия Представители узкого локализационизма рассматривали психологические функции как единые, неразложимые на компоненты «психические способности», реализуемые ограниченными… … Большая психологическая энциклопедия
Представители узкого локализационизма рассматривали психологические функции как единые, неразложимые на компоненты «психические способности», реализуемые ограниченными… … Большая психологическая энциклопедияНейропсихология и основные термины — chandervander — LiveJournal
Предметом нейропсихологии является изучение особенностей нарушения психических процессов, состояний и личности в целом при локальных поражениях мозга.
«Нейропсихологический симптом» – нарушение психической функции (как элементарной, так и высшей), возникающее вследствие локального поражения головного мозга.
«Первичные нейропсихологические симптомы» – нарушения психических функций, непосредственно связанные с локальным органическим повреждением головного мозга под воздействием биологического фактора.
«Вторичные нейропсихологические симптомы» – нарушения психических функций, возникающие как системное следствие первичных нейропсихологических симптомов.
«Нейропсихологический синдром» – закономерное сочетание нейропсихологических симптомов, связанное с выпадением (или нарушением) определенного фактора (или нескольких факторов).
«Нейропсихологический фактор» – физиологическое понятие, обозначающее принцип физиологической деятельности определенной мозговой структуры (участка мозга), нарушение которого ведет к появлению нейропсихологического синдрома.
«Синдромный анализ» – анализ нейропсихологических синдромов с целью обнаружения общего основания (фактора), объясняющего происхождение различных нейропсихологических симптомов (синоним — «факторный анализ»).
«Нейропсихологическая диагностика» — исследование больных с локальными поражениями головного мозга с помощью нейропсихологических методов с целью установления места поражения мозга (т. е. с целью постановки топического диагноза).
е. с целью постановки топического диагноза).
«Функциональная система» – физиологическое понятие, заимствованное из концепции функциональных систем П. К. Анохина, используемое для объяснения физиологической основы высших психических функций; совокупность афферентных и эфферентных механизмов (звеньев), объединенных в единую систему для достижения конечного полезного результата. Различные по содержанию высшие психические функции (гностические, мнестические, интеллектуальные и др.) обеспечиваются качественно различными функциональными системами.
«Мозговые механизмы высших психических функций» (или «мозговая организация психической деятельности») — совокупность морфологических структур (зон, участков) в коре больших полушарий и в подкорковых образованиях и протекающих в них физиологических процессов, входящих в единую систему и необходимых для осуществления психической деятельности; морфо-физиологическая основа психических функций, синоним «локализации высших психических функций».
«Полифункциональность мозговых структур» – положение, согласно которому мозговые структуры (особенно ассоциативные зоны коры больших полушарий) под влиянием новых афферентных воздействий могут перестраивать свои функции; на этом положении основаны принципы внутрисистемной и межсистемной перестройки высших психических функций, нарушенных вследствие локальных поражений головного мозга.
«Норма функции» – понятие, на котором основано нейропсихологическое изучение нарушений высших психических функций. Показатели реализации функции (в психологических единицах продуктивности, объема, скорости и т. д.), которые характеризуют среднюю норму в данной популяции (т. е. показатели, характерные для подавляющего большинства здоровых людей).
«Межполушарная асимметрия мозга» – неравноценность, качественное различие того «вклада», который делает левое и правое полушария мозга в каждую психическую функцию; различия в мозговой организации высших психических функций в левом и правом полушариях мозга.
«Функциональная специфичность больших полушарий» – специфика переработки информации и мозговой организации функций, присущая левому или правому полушарию головного мозга, определяемая интегральными полушарными факторами.
«Межполушарное взаимодействие» – особый механизм объединения левого и правого полушарий в единую целостно функционирующую систему, формирующийся в онтогенезе.
нейропсихология — это… Что такое нейропсихология?
- нейропсихология
НЕЙРОПСИХОЛО́ГИЯ -и; ж. Раздел психологии, изучающий психическую деятельность человека в связи с функционированием центральной нервной системы в определённых условиях.
◁ Нейропсихологи́ческий, -ая, -ое.
* * *
нейропсихоло́гияотрасль психологии, изучающая мозговую основу психических процессов и их связь с отдельными системами головного мозга; первоначально развивалась как раздел неврологии.
* * *
НЕЙРОПСИХОЛОГИЯНЕЙРОПСИХОЛО́ГИЯ, отрасль психологии, изучающая мозговую основу психических процессов и их связь с отдельными системами головного мозга; первоначально развивалась как раздел неврологии.

Энциклопедический словарь. 2009.
- нейропептиды
- нейросекреция
Смотреть что такое «нейропсихология» в других словарях:
нейропсихология — нейропсихология … Орфографический словарь-справочник
нейропсихология — отрасль психологической науки, сложившаяся на стыке психологии, медицины (неврологии, нейрохирургии) и физиологии, изучающая мозговые механизмы высших психических функций на материале локальных поражений головного мозга (см. компенсация… … Большая психологическая энциклопедия
Нейропсихология — Нейропсихология междисциплинарное научное направление, лежащее на стыке психологии и нейронауки, нацелена на понимание связи структуры и функционирования головного мозга с психическими процессами и поведением живых существ.
Термин… … ВикипедияНЕЙРОПСИХОЛОГИЯ — отрасль психологии, изучающая мозговую основу психических процессов и их связь с отдельными системами головного мозга; первоначально развивалась как раздел неврологии … Большой Энциклопедический словарь
Нейропсихология — раздел психологии, посвященный изучению мозговых механизмов высших психических функций на материале наблюдений за локальными поражениями головного мозга. Основы отечественной нейропсихологии были заложены А.Р. Лурия , который разработал теорию… … Психологический словарь
НЕЙРОПСИХОЛОГИЯ — [Словарь иностранных слов русского языка
нейропсихология — сущ., кол во синонимов: 3 • медицина (189) • нейросемантика (1) • психология (21) … Словарь синонимов
НЕЙРОПСИХОЛОГИЯ — (от греч. neuron – жила, нерв + психология). Отрасль психологии, изучающая содержание психических процессов и их связь с отдельными системами головного мозга.
Н. имеет важное значение для понимания механизмов психических процессов. Новые… … Новый словарь методических терминов и понятий (теория и практика обучения языкам)НЕЙРОПСИХОЛОГИЯ — наука, изучающая мозговую организацию психических процессов. Нейропсихология основана на наблюдении и экспериментах, которые проводятся в клиниках и лабораториях. К числу экспериментальных методов относятся электрическая стимуляция мозга и… … Энциклопедия Кольера
нейропсихология — (нейро + психология) раздел психологии, изучающий связь психических процессов с определенными системами головного мозга … Большой медицинский словарь
 Термин… … Википедия
Термин… … Википедия Н. имеет важное значение для понимания механизмов психических процессов. Новые… … Новый словарь методических терминов и понятий (теория и практика обучения языкам)
Н. имеет важное значение для понимания механизмов психических процессов. Новые… … Новый словарь методических терминов и понятий (теория и практика обучения языкам)Когнитивные функции и основные параметры физиологии человеческого мозга связаны с хронотипом
Участники
Тридцать два здоровых молодых взрослых добровольца (16 женщин, средний возраст = 26,62 ± 5,01) квалифицированы как ранний хронотип (ЭК) или поздний хронотип (LC) были набраны из Технического университета Дортмунда, Рурского университета Бохума и окружающего сообщества. Хронотип определялся на основании баллов по опроснику «Утро-вечерность» (DMEQ) 68 .Из 269 добровольцев, выполнивших DMEQ в ходе эксперимента, 69 были вечерними типами (6 определенных вечерних типов) и 35 — утренними типами (5 определенных утренних типов). Чтобы ограничить вариабельность хронотипов во времени и продолжительности сна, были включены только умеренные хронотипы. Шестнадцать человек от каждого хронотипа (средний тип), которые соответствовали критериям включения, были включены в группы EC (8 женщин, N = 16) и LC (8 женщин, N = 16). Размер выборки был рассчитан априори на основе анализа мощности, который показал, что для среднего размера эффекта (частичный квадрат этажа = 0.10) (предлагается для исследований NIBS 69 ), требуется минимум 24 субъекта для достижения 95% мощности при альфа 0,05 для первичного статистического теста смешанной модели ANOVA. Мы увеличили размер выборки до 32, чтобы полностью уравновесить порядок задач в каждой группе ( N = 16) и компенсировать непредвиденную изменчивость и отсев.
Хронотип определялся на основании баллов по опроснику «Утро-вечерность» (DMEQ) 68 .Из 269 добровольцев, выполнивших DMEQ в ходе эксперимента, 69 были вечерними типами (6 определенных вечерних типов) и 35 — утренними типами (5 определенных утренних типов). Чтобы ограничить вариабельность хронотипов во времени и продолжительности сна, были включены только умеренные хронотипы. Шестнадцать человек от каждого хронотипа (средний тип), которые соответствовали критериям включения, были включены в группы EC (8 женщин, N = 16) и LC (8 женщин, N = 16). Размер выборки был рассчитан априори на основе анализа мощности, который показал, что для среднего размера эффекта (частичный квадрат этажа = 0.10) (предлагается для исследований NIBS 69 ), требуется минимум 24 субъекта для достижения 95% мощности при альфа 0,05 для первичного статистического теста смешанной модели ANOVA. Мы увеличили размер выборки до 32, чтобы полностью уравновесить порядок задач в каждой группе ( N = 16) и компенсировать непредвиденную изменчивость и отсев. Поскольку пол и возраст могут объяснять различия в хронотипе, мы сбалансировали пол участников и сохранили возрастной диапазон до раннего взросления. Все участники были правшами, некурящими и прошли медицинское обследование для проверки отсутствия в анамнезе неврологических заболеваний, эпилепсии или припадков, лекарств, металлических имплантатов и текущей беременности.Каждый участник принял участие в тестовой сессии TMS, чтобы познакомиться с ощущением стимуляции и понять протокол исследования. Женщины не обследовались во время лютеиновой фазы менструального цикла (примерно на 3 неделе после менструации), чтобы гарантировать, что гормональные изменения, влияющие на возбудимость коры головного мозга 70 , не повлияют на их хронотип. Потребление кофеина и другие факторы, потенциально влияющие на возбудимость коры головного мозга (например, массивная физическая активность), контролировались перед каждым экспериментальным сеансом, и в случае несоблюдения соответствующих критериев эти сеансы откладывались.
Поскольку пол и возраст могут объяснять различия в хронотипе, мы сбалансировали пол участников и сохранили возрастной диапазон до раннего взросления. Все участники были правшами, некурящими и прошли медицинское обследование для проверки отсутствия в анамнезе неврологических заболеваний, эпилепсии или припадков, лекарств, металлических имплантатов и текущей беременности.Каждый участник принял участие в тестовой сессии TMS, чтобы познакомиться с ощущением стимуляции и понять протокол исследования. Женщины не обследовались во время лютеиновой фазы менструального цикла (примерно на 3 неделе после менструации), чтобы гарантировать, что гормональные изменения, влияющие на возбудимость коры головного мозга 70 , не повлияют на их хронотип. Потребление кофеина и другие факторы, потенциально влияющие на возбудимость коры головного мозга (например, массивная физическая активность), контролировались перед каждым экспериментальным сеансом, и в случае несоблюдения соответствующих критериев эти сеансы откладывались. Это исследование соответствовало руководящим принципам Хельсинкской декларации и было одобрено институциональным наблюдательным советом Исследовательского центра Лейбница по производственной среде и человеческому фактору. Участники дали информированное согласие и получили денежную компенсацию. Они могли выйти в любое время.
Это исследование соответствовало руководящим принципам Хельсинкской декларации и было одобрено институциональным наблюдательным советом Исследовательского центра Лейбница по производственной среде и человеческому фактору. Участники дали информированное согласие и получили денежную компенсацию. Они могли выйти в любое время.
Опросник по утрам и вечерам (MEQ)
Немецкая версия MEQ 71 (DMEQ) 68 использовалась для определения хронотипов. Он состоит из 19 вопросов, которые просят людей определить их ритмы «наилучшего самочувствия», указать предпочтительные временные интервалы часов, а не реальное время сна и участие в других ежедневных / еженедельных занятиях (например.g., физические упражнения, тесты, работа), а также оценить утреннюю бодрость, утренний аппетит и вечернюю усталость. Каждый вопрос имеет балл, и сумма баллов варьируется от 16 до 86, где баллы ниже 42 указывают на вечерний тип или поздний хронотип (LC), а баллы выше 58 указывают на утренний тип или ранний хронотип (EC). Определенный вечер (16–30), умеренный вечер (31–41), средний или нейтральный (42–58), умеренное утро (59–69) и определенное утро (70–86) — это пять категорий хронотипов, определенных MEQ. .Анкета демонстрирует высокую достоверность сообщений и имеет значительную корреляцию с гормональными изменениями, связанными с циркадным ритмом, включая мелатонин 68 . В дополнение к MEQ мы определили фенотип дихотомического хронотипа, задав два идентичных вопроса («Вы от природы ночной человек или утренний человек?») 2 . Лица с несогласованными или нейтральными ответами на оба вопроса были исключены. Средний балл хронотипа для раннего и позднего хронотипов составил 62,25 ± 3.47 и 35,37 ± 4,31, показывая, что обе группы представляют умеренные ранние и поздние хронотипы.
Определенный вечер (16–30), умеренный вечер (31–41), средний или нейтральный (42–58), умеренное утро (59–69) и определенное утро (70–86) — это пять категорий хронотипов, определенных MEQ. .Анкета демонстрирует высокую достоверность сообщений и имеет значительную корреляцию с гормональными изменениями, связанными с циркадным ритмом, включая мелатонин 68 . В дополнение к MEQ мы определили фенотип дихотомического хронотипа, задав два идентичных вопроса («Вы от природы ночной человек или утренний человек?») 2 . Лица с несогласованными или нейтральными ответами на оба вопроса были исключены. Средний балл хронотипа для раннего и позднего хронотипов составил 62,25 ± 3.47 и 35,37 ± 4,31, показывая, что обе группы представляют умеренные ранние и поздние хронотипы.
Время эксперимента, цикл сна / бодрствования и изменение освещенности
Время эксперимента определялось на основе среднего времени сна и уровня активности ранних и поздних хронотипов на основе предыдущих исследований 6,62 , а также ежедневных распорядок дня жизни. Для утренних занятий время начала выбиралось исходя из нормы начала учебы и работы (8 часов утра).Это также время, когда оба хронотипа имеют самый низкий уровень активности 62 . Вечернее время (19:00) было указано на основе времени, когда оба хронотипа имеют сопоставимый уровень активности в соответствии с предыдущими профилями набора данных 62 . Обе группы были проинструктированы ложиться спать (около 22: 00–23: 00), вставать утром в одно и то же время дня, а также спать не менее 8 часов для утренних сессий (среднее значение ЭК = 8,41 ч, среднее значение LC = 8.11 ч). На вечернем сеансе участникам разрешалось лечь спать в удобное для них время, но до 24:00, а также при необходимости разрешалось спать более 8 часов (среднее значение ЭК, = 8,36 ч, среднее значение ЭК, = 8,38). час). Всем участникам настоятельно рекомендовалось соблюдать инструкции цикла сна / бодрствования, а в случае нарушений, плохого качества сна и сонливости (измеряемой оценкой участников перед каждым сеансом) экспериментальный сеанс переносился на другой день.
Для утренних занятий время начала выбиралось исходя из нормы начала учебы и работы (8 часов утра).Это также время, когда оба хронотипа имеют самый низкий уровень активности 62 . Вечернее время (19:00) было указано на основе времени, когда оба хронотипа имеют сопоставимый уровень активности в соответствии с предыдущими профилями набора данных 62 . Обе группы были проинструктированы ложиться спать (около 22: 00–23: 00), вставать утром в одно и то же время дня, а также спать не менее 8 часов для утренних сессий (среднее значение ЭК = 8,41 ч, среднее значение LC = 8.11 ч). На вечернем сеансе участникам разрешалось лечь спать в удобное для них время, но до 24:00, а также при необходимости разрешалось спать более 8 часов (среднее значение ЭК, = 8,36 ч, среднее значение ЭК, = 8,38). час). Всем участникам настоятельно рекомендовалось соблюдать инструкции цикла сна / бодрствования, а в случае нарушений, плохого качества сна и сонливости (измеряемой оценкой участников перед каждым сеансом) экспериментальный сеанс переносился на другой день. Субъективная сонливость участников и их настороженность оценивались с помощью шкалы Каролинской сонливости (KSS) 72 , которая измеряет сонливость по шкале Лайкерта от 1 до 10 (дополнительная информация). Не наблюдалось значительной разницы между оценками сонливости как для ранних, так и для поздних хронотипов на утренних сессиях (среднее значение ЭК = 3,2, среднее значение LC = 3,71; t = 1,61, P = 0,11) и вечерних сессиях ( среднее значение ЭК = 3.58, среднее значение LC = 3,65; т = 0,25, P = 0,80). Из-за изменчивости сезонного освещения экспериментальные сеансы не проводились летом, когда разница между восходом и заходом солнца самая большая в Германии. В другие месяцы эксперимент было запланировано случайным образом распределить по другим сезонам в обеих группах. Кроме того, окружающий свет поддерживался постоянным со стандартной интенсивностью экспозиции для внутреннего освещения (около 500 люкс) на протяжении всех экспериментальных сессий утром и вечером.
Субъективная сонливость участников и их настороженность оценивались с помощью шкалы Каролинской сонливости (KSS) 72 , которая измеряет сонливость по шкале Лайкерта от 1 до 10 (дополнительная информация). Не наблюдалось значительной разницы между оценками сонливости как для ранних, так и для поздних хронотипов на утренних сессиях (среднее значение ЭК = 3,2, среднее значение LC = 3,71; t = 1,61, P = 0,11) и вечерних сессиях ( среднее значение ЭК = 3.58, среднее значение LC = 3,65; т = 0,25, P = 0,80). Из-за изменчивости сезонного освещения экспериментальные сеансы не проводились летом, когда разница между восходом и заходом солнца самая большая в Германии. В другие месяцы эксперимент было запланировано случайным образом распределить по другим сезонам в обеих группах. Кроме того, окружающий свет поддерживался постоянным со стандартной интенсивностью экспозиции для внутреннего освещения (около 500 люкс) на протяжении всех экспериментальных сессий утром и вечером. Во время познавательных занятий задания и записи ЭЭГ проводились в звукоизолированном экранированном помещении с постоянной температурой 24–25 ° C для утренних и вечерних сеансов.
Во время познавательных занятий задания и записи ЭЭГ проводились в звукоизолированном экранированном помещении с постоянной температурой 24–25 ° C для утренних и вечерних сеансов.
Мониторинг корковой возбудимости с помощью TMS
Протоколы одноимпульсной и парноимпульсной ТМС для моторного порога покоя (RMT), активного моторного порога (AMT), кривой I – O, короткого интракортикального торможения и облегчения (SICI-ICF), внутрикортикального Облегчение I-волны и кратковременное афферентное торможение (SAI) использовались для мониторинга кортикоспинальной и внутрикортикальной возбудимости в моторной коре.RMT, AMT и кривая I – O исследуют кортикоспинальную возбудимость, SICI-ICF измеряет как внутрикортикальное облегчение, так и торможение, а интракортикальное облегчение I-волны и SAI измеряет внутрикортикальное ингибирование моторной коры человека 31,32,36 .
Одноимпульсный MEP, покой и активный моторный порог
Одноимпульсный двухфазный TMS с частотой 0,25 Гц ± 10% (случайный) подавался магнитным стимулятором PowerMag ppTMS (Mag & More, Мюнхен, Германия) через восьмерку магнитная катушка (диаметр одной обмотки 70 мм; пиковое магнитное поле 2 Тл), удерживаемая под углом 45 ° к средней линии и накладываемая на левую первичную моторную кору.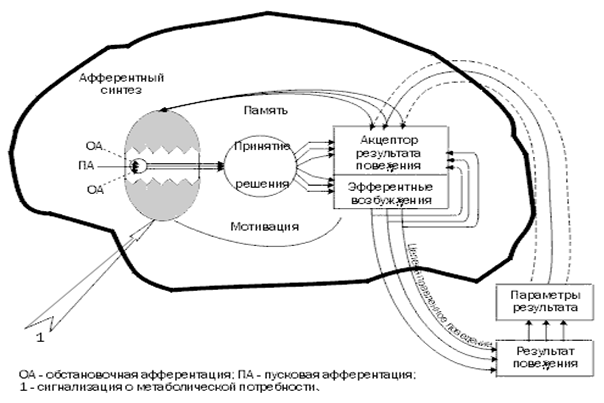 Поверхностные MEP регистрировались от правой мышцы, отводящей минимальный палец (ADM) с помощью золотых чашечных электродов при монтаже живота и сухожилия. Сигналы усиливались и фильтровались (1000; 3 Гц – 3 кГц) с помощью D440-2 (Digitimer, Велвин-Гарден-Сити, Великобритания) и оцифровывались (частота дискретизации 5 кГц) с помощью преобразователя Micro 1401 AD (Cambridge Electronic Design, Кембридж , Великобритания), контролируемый Signal Software (Cambridge Electronic Design, v. 2.13). RMT был исследован с помощью инструмента оценки моторных пороговых значений TMS (MTAT 2.0, http: // www.Clinicalresearcher.org/software.htm) 73 и была определена как наименьшая интенсивность стимулятора, необходимая для вызова максимального размаха MEP 50 мкВ в расслабленной мышце ADM по крайней мере в пяти из десяти последовательных испытаний. AMT была определена как самая низкая интенсивность стимулятора, необходимая для вызова MEP-ответа ∼200–300 мкВ во время умеренного тонического сокращения правой ADM-мышцы (∼20% от максимальной мышечной силы) 74 по крайней мере в трех из шести последовательных испытаний.
Поверхностные MEP регистрировались от правой мышцы, отводящей минимальный палец (ADM) с помощью золотых чашечных электродов при монтаже живота и сухожилия. Сигналы усиливались и фильтровались (1000; 3 Гц – 3 кГц) с помощью D440-2 (Digitimer, Велвин-Гарден-Сити, Великобритания) и оцифровывались (частота дискретизации 5 кГц) с помощью преобразователя Micro 1401 AD (Cambridge Electronic Design, Кембридж , Великобритания), контролируемый Signal Software (Cambridge Electronic Design, v. 2.13). RMT был исследован с помощью инструмента оценки моторных пороговых значений TMS (MTAT 2.0, http: // www.Clinicalresearcher.org/software.htm) 73 и была определена как наименьшая интенсивность стимулятора, необходимая для вызова максимального размаха MEP 50 мкВ в расслабленной мышце ADM по крайней мере в пяти из десяти последовательных испытаний. AMT была определена как самая низкая интенсивность стимулятора, необходимая для вызова MEP-ответа ∼200–300 мкВ во время умеренного тонического сокращения правой ADM-мышцы (∼20% от максимальной мышечной силы) 74 по крайней мере в трех из шести последовательных испытаний. .
.
Кривая ввода-вывода (кривая ввода-вывода)
Кривая ввода-вывода представляет собой одноимпульсный протокол TMS, который отражает возбудимость кортикоспинальных нейронов.Он модулируется глутаматергической активностью и относится к увеличению амплитуды MEP с увеличением интенсивности TMS 32 . Наклон кривой набора увеличивается при более высоких интенсивностях ТМС с более высокой глутаматергической и адренергической передачей и уменьшается при приеме препаратов, усиливающих эффекты ГАМК 32,35 . В протоколе кривой I – O амплитуды MEP в расслабленной правой ADM-мышце измерялись в четырех блоках с различной интенсивностью стимула (100%, 110%, 130% и 150% RMT) 75 , каждый блок с 15 импульсами, и среднее значение (амплитуды МВП) было рассчитано для каждой интенсивности.
Кратковременное внутрикортикальное ингибирование и внутрикортикальное облегчение (SICI-ICF)
SICI-ICF — это протокол парных импульсов TMS для мониторинга кортикального торможения, опосредованного ГАМК-эргическим действием, и опосредованного глутаматом коркового облегчения 32 . В этом протоколе за подпороговым условным стимулом (определяемым как 70% AMT) следует надпороговый тестовый стимул, который был скорректирован таким образом, чтобы вызвать исходный MEP ~ 1 мВ. Парные стимулы представлены в интерстимулах (ISI) 2, 3, 5, 10 и 15 мс 36 .ISI в 2 и 3 мс представляют интракортикальное торможение с короткой задержкой (SICI) и оказывают тормозящее влияние на амплитуды тестовых импульсов MEP, а ISI в 10 и 15 мс представляют внутрикортикальное облегчение (ICF) и оказывают усиливающее влияние на MEP, вызванное одноимпульсным TMS. амплитуды 36,76,77 . Стимулы (подпороговые и надпороговые) были организованы в блоки, в которых каждый ISI и один тестовый стимул применялись один раз в псевдослучайном порядке. Каждый блок повторялся 15 раз, в результате чего в общей сложности получалось 90 одноимпульсных или парных MEP за сеанс.Точный интервал между парными импульсами был рандомизирован (4 ± 0,4 с).
В этом протоколе за подпороговым условным стимулом (определяемым как 70% AMT) следует надпороговый тестовый стимул, который был скорректирован таким образом, чтобы вызвать исходный MEP ~ 1 мВ. Парные стимулы представлены в интерстимулах (ISI) 2, 3, 5, 10 и 15 мс 36 .ISI в 2 и 3 мс представляют интракортикальное торможение с короткой задержкой (SICI) и оказывают тормозящее влияние на амплитуды тестовых импульсов MEP, а ISI в 10 и 15 мс представляют внутрикортикальное облегчение (ICF) и оказывают усиливающее влияние на MEP, вызванное одноимпульсным TMS. амплитуды 36,76,77 . Стимулы (подпороговые и надпороговые) были организованы в блоки, в которых каждый ISI и один тестовый стимул применялись один раз в псевдослучайном порядке. Каждый блок повторялся 15 раз, в результате чего в общей сложности получалось 90 одноимпульсных или парных MEP за сеанс.Точный интервал между парными импульсами был рандомизирован (4 ± 0,4 с).
Облегчение коротких интервалов внутрикортикальной I-волны
Этот протокол TMS основан на I (непрямых) волнах, которые относятся к высокочастотным повторяющимся разрядам кортикоспинальных нейронов, вызванным одноимпульсной стимуляцией моторной коры 78 (для подробный обзор см. ссылки 37,78 ). В этом протоколе два последовательных стимула (над- и подпороговый) разделены короткими ISI, но этот протокол включает надпороговый первый стимул и подпороговый второй стимул 37 .ISI варьируются от 1,1 мс до 4,5 мс с задержкой и представлены в псевдослучайном порядке. Мы сгруппировали ISI в ранние (средняя MEP при ISI 1.1, 1.3, 1.5 мс), средняя (средняя MEP при ISI 2.3, 2.5, 2.7, 2.9 мс) и поздняя (средняя MEP при ISI 4.1, 4.3, 4.5 мс) эпохи. Интенсивность первого кондиционирующего надпорогового стимула (S1) регулируется для получения исходного MEP ~ 1 мВ, когда его вводят отдельно, и за ним следует второй подпороговый стимул (S2), который был установлен на уровне 70% от RMT) 75 . Для каждого ISI было зарегистрировано 15 импульсов.Еще 15 импульсов были записаны для контрольных MEP, в которых суперпороговый стимул (S1) подавался отдельно и корректировался для достижения исходного MEP ~ 1 мВ. Пары стимулов были организованы в блоки, в которых каждый ISI и один тестовый импульс были представлены один раз и были псевдослучайно.
ссылки 37,78 ). В этом протоколе два последовательных стимула (над- и подпороговый) разделены короткими ISI, но этот протокол включает надпороговый первый стимул и подпороговый второй стимул 37 .ISI варьируются от 1,1 мс до 4,5 мс с задержкой и представлены в псевдослучайном порядке. Мы сгруппировали ISI в ранние (средняя MEP при ISI 1.1, 1.3, 1.5 мс), средняя (средняя MEP при ISI 2.3, 2.5, 2.7, 2.9 мс) и поздняя (средняя MEP при ISI 4.1, 4.3, 4.5 мс) эпохи. Интенсивность первого кондиционирующего надпорогового стимула (S1) регулируется для получения исходного MEP ~ 1 мВ, когда его вводят отдельно, и за ним следует второй подпороговый стимул (S2), который был установлен на уровне 70% от RMT) 75 . Для каждого ISI было зарегистрировано 15 импульсов.Еще 15 импульсов были записаны для контрольных MEP, в которых суперпороговый стимул (S1) подавался отдельно и корректировался для достижения исходного MEP ~ 1 мВ. Пары стимулов были организованы в блоки, в которых каждый ISI и один тестовый импульс были представлены один раз и были псевдослучайно. Этот протокол парных импульсов TMS (первый надпороговый стимул и второй подпороговый стимул) оказывает облегчающее действие на пики MEP 37 , которые возникают при ISI примерно 1,3, 2,6 и 4,2 мс. Предполагается, что этот эффект возникает в результате вызванных I-волн (непрямые волны: нисходящие залпы, производимые непрямой активацией нейронов пирамидного тракта через пресинаптические нейроны) подпороговым S2 и контролируется связанными с ГАМК нейронными цепями 35,37 , 79
Этот протокол парных импульсов TMS (первый надпороговый стимул и второй подпороговый стимул) оказывает облегчающее действие на пики MEP 37 , которые возникают при ISI примерно 1,3, 2,6 и 4,2 мс. Предполагается, что этот эффект возникает в результате вызванных I-волн (непрямые волны: нисходящие залпы, производимые непрямой активацией нейронов пирамидного тракта через пресинаптические нейроны) подпороговым S2 и контролируется связанными с ГАМК нейронными цепями 35,37 , 79
Коротко-латентное афферентное торможение (SAI)
SAI — это протокол TMS в сочетании со стимуляцией периферических нервов, основанный на концепции, согласно которой периферические соматосенсорные входы оказывают тормозящее влияние на возбудимость моторной коры через короткие промежутки времени (например,g., 20–40 мс) 38 . SAI был связан с холинергической 31 и ГАМКергической системами 33 на корковом уровне. В этом протоколе одноимпульсный TMS служит в качестве тестового стимула и настраивается так, чтобы вызвать ответ MEP с размахом амплитуды ~ 1 мВ. Обусловливающие афферентные стимулы представляли собой одиночные импульсы (200 мкс) электрической стимуляции, прикладываемые к правому локтевому нерву на уровне запястья (проксимальный катод) через биполярные электроды, подключенные к стимулятору Digitimer D185 (Digitimer Ltd., Велвин-Гарден-Сити, Великобритания). Обусловливающие афферентные стимулы применялись с интенсивностью примерно в 2,5–3 раза превышающей порог восприятия, отрегулированной таким образом, чтобы вызвать минимальное видимое подергивание тенарных мышц 31 , за которым следовало одиночный импульс TMS (тестовый стимул), применяемый к моторному кортикальному представлению правильный ADM. Стимулы применялись в блоках, содержащих только тестовый стимул (контрольное условие) и два блока парных стимулов с ISI 20 и 40 мс в псевдослучайном порядке. Каждый блок повторялся 20 раз, в результате чего было проведено 60 попыток.
Обусловливающие афферентные стимулы представляли собой одиночные импульсы (200 мкс) электрической стимуляции, прикладываемые к правому локтевому нерву на уровне запястья (проксимальный катод) через биполярные электроды, подключенные к стимулятору Digitimer D185 (Digitimer Ltd., Велвин-Гарден-Сити, Великобритания). Обусловливающие афферентные стимулы применялись с интенсивностью примерно в 2,5–3 раза превышающей порог восприятия, отрегулированной таким образом, чтобы вызвать минимальное видимое подергивание тенарных мышц 31 , за которым следовало одиночный импульс TMS (тестовый стимул), применяемый к моторному кортикальному представлению правильный ADM. Стимулы применялись в блоках, содержащих только тестовый стимул (контрольное условие) и два блока парных стимулов с ISI 20 и 40 мс в псевдослучайном порядке. Каждый блок повторялся 20 раз, в результате чего было проведено 60 попыток.
Методика эксперимента
Сеансы мониторинга корковой возбудимости проводились один раз утром и один раз вечером в одно и то же фиксированное время для всех сеансов исследования. Измерения были запланированы на 8:30 утра для утреннего сеанса и 19:00 для вечернего сеанса после этапа подготовки (идентификация горячих точек моторной коры, процедуры определения RMT и AMT). Между сеансами был интервал в 1 неделю. Участников проинструктировали не употреблять кофеин, алкоголь и не заниматься интенсивными физическими нагрузками за 24 часа до каждого сеанса, чтобы обеспечить стабильный уровень моторно-корковой возбудимости.На каждом сеансе участники удобно сидели в кресле с откидной спинкой, под правой рукой лежала подушка, а вокруг шеи — вакуумная подушка, чтобы предотвратить движение головы. Сначала горячая точка (положение катушки над областью первичного двигателя, которая производит наибольшую MEP в правом ADM с заданной средней интенсивностью TMS) была идентифицирована с помощью TMS. Затем интенсивность стимуляции регулировали, чтобы вызвать MEP с размахом амплитуды в среднем 1 мВ, как описано выше. После этого шага были получены RMT и AMT, как описано выше.После записи AMT был сделан 10-минутный перерыв, чтобы избежать влияния сокращения мышц на следующие измерения.
Измерения были запланированы на 8:30 утра для утреннего сеанса и 19:00 для вечернего сеанса после этапа подготовки (идентификация горячих точек моторной коры, процедуры определения RMT и AMT). Между сеансами был интервал в 1 неделю. Участников проинструктировали не употреблять кофеин, алкоголь и не заниматься интенсивными физическими нагрузками за 24 часа до каждого сеанса, чтобы обеспечить стабильный уровень моторно-корковой возбудимости.На каждом сеансе участники удобно сидели в кресле с откидной спинкой, под правой рукой лежала подушка, а вокруг шеи — вакуумная подушка, чтобы предотвратить движение головы. Сначала горячая точка (положение катушки над областью первичного двигателя, которая производит наибольшую MEP в правом ADM с заданной средней интенсивностью TMS) была идентифицирована с помощью TMS. Затем интенсивность стимуляции регулировали, чтобы вызвать MEP с размахом амплитуды в среднем 1 мВ, как описано выше. После этого шага были получены RMT и AMT, как описано выше.После записи AMT был сделан 10-минутный перерыв, чтобы избежать влияния сокращения мышц на следующие измерения. После перерыва следующие протоколы TMS были мерами для мониторинга возбудимости коры: SAI, SICI-ICF, облегчение I-волны и кривая ввода-вывода. Порядок измерений был рандомизирован, за исключением SAI, который всегда был первым мероприятием, так как требовалось время на подготовку около 10 минут, которое планировалось провести во время 10-минутного перерыва. Это было сделано для того, чтобы продолжительность сеанса была как можно ближе к сеансам tDCS и чтобы участники придерживались своего предпочтительного и нежелательного циркадного ритма.В случае изменения MEP, вызванного одиночным тестовым импульсом, на> 20% во время сеанса, интенсивность стимуляции была скорректирована 80 . Каждый сеанс корковой возбудимости длился 60–70 мин. Все протоколы ТМС проводились с помощью магнитного стимулятора PowerMag ppTMS (Mag & More, Мюнхен, Германия) через магнитную катушку в форме восьмерки (диаметр одной обмотки, 70 мм; пиковое магнитное поле, 2 Тл), удерживаемую под углом 45 ° к Срединная линия и накладывалась на левую первичную моторную кору.
После перерыва следующие протоколы TMS были мерами для мониторинга возбудимости коры: SAI, SICI-ICF, облегчение I-волны и кривая ввода-вывода. Порядок измерений был рандомизирован, за исключением SAI, который всегда был первым мероприятием, так как требовалось время на подготовку около 10 минут, которое планировалось провести во время 10-минутного перерыва. Это было сделано для того, чтобы продолжительность сеанса была как можно ближе к сеансам tDCS и чтобы участники придерживались своего предпочтительного и нежелательного циркадного ритма.В случае изменения MEP, вызванного одиночным тестовым импульсом, на> 20% во время сеанса, интенсивность стимуляции была скорректирована 80 . Каждый сеанс корковой возбудимости длился 60–70 мин. Все протоколы ТМС проводились с помощью магнитного стимулятора PowerMag ppTMS (Mag & More, Мюнхен, Германия) через магнитную катушку в форме восьмерки (диаметр одной обмотки, 70 мм; пиковое магнитное поле, 2 Тл), удерживаемую под углом 45 ° к Срединная линия и накладывалась на левую первичную моторную кору. Устройство было оборудовано для применения как одноимпульсной ТМС, так и парно-импульсной ТМС.
Устройство было оборудовано для применения как одноимпульсной ТМС, так и парно-импульсной ТМС.
Индукция нейропластичности с помощью tDCS
Стимуляция моторной коры постоянным током
Электрический постоянный ток подавался через пару губчатых электродов, пропитанных физиологическим раствором (35 см 2 ), и подавался через стимулятор постоянного тока с батарейным питанием ( neuroConn GmbH, Ильменау, Германия). Целевой электрод фиксировали над моторно-кортикальной репрезентативной областью правого ADM, как определено с помощью TMS, а контрольный электрод помещали над контрлатеральной надглазничной областью.Расстояние на коже черепа между краями электродов сохранялось минимум 6 см, чтобы уменьшить шунтирование тока через кожу головы 81 . В зависимости от рандомизированных условий, анодные, катодные или фиктивные tDCS с интенсивностью 1 мА применялись в течение 7 минут с 15-секундным повышением / понижением в начале и в конце стимуляции. Для фиктивного состояния стимуляция проводилась в течение 30 секунд с 30-секундным нарастанием и спадом. Используя эту процедуру, участники не могут отличить настоящую tDCS от фиктивной 82 .Мы смоделировали поток электрического тока в голове, индуцированный этим протоколом, чтобы показать, как tDCS влияет на первичную моторную кору (рис. 3а). Модель основана на стандартной модели головы, которая не учитывает возможные структурные различия (например, белое вещество) в головном мозге разных хронотипов 64 . Таким образом, модель следует принимать во внимание только в иллюстративных целях.
Используя эту процедуру, участники не могут отличить настоящую tDCS от фиктивной 82 .Мы смоделировали поток электрического тока в голове, индуцированный этим протоколом, чтобы показать, как tDCS влияет на первичную моторную кору (рис. 3а). Модель основана на стандартной модели головы, которая не учитывает возможные структурные различия (например, белое вещество) в головном мозге разных хронотипов 64 . Таким образом, модель следует принимать во внимание только в иллюстративных целях.
Мониторинг MEP с помощью TMS
Одноимпульсные MEP были получены таким же образом, как описано в предыдущем разделе, а интенсивность TMS была настроена так, чтобы вызывать MEP с размахом амплитуды ~ 1 мВ.
Экспериментальная процедура
После отбора участников на пригодность для tDCS, каждый участник посетил шесть сеансов tDCS (утренний анодный, утренний катодный, утренний имитационный и вечерний анодный, вечерний катодный, вечерний имитационный) в случайном порядке. Сеансы TDCS начинались в фиксированное время утром и вечером с интервалом в 1 неделю между сеансами. Утренние занятия начинались в 8:00, а вечерние — в 18:30; время начала tDCS было запланировано примерно на 8:30 на утренней сессии и 7:00 на вечерней сессии, после подготовительного этапа, который длился ~ 20–30 минут.На каждом сеансе участники удобно сидели в кресле с откидной спинкой, под правой рукой располагалась подушка, а вокруг шеи — вакуумная подушка, чтобы предотвратить движение головы. В начале каждого сеанса измеряли базовую возбудимость коры головного мозга, сначала индуцируя MEP над левым M1, чтобы идентифицировать область, которая произвела наибольшую MEP целевой мышцы (правая ADM) с заданной интенсивностью TMS. Затем область была отмечена, и последующие импульсы подавались из этого оптимального положения.Интенсивность стимуляции регулировали для достижения максимальной амплитуды МВП 1 мВ (SI1 мВ), которая затем использовалась для остальных измерений. После базового измерения 25 MEP было проведено 7 минут анодной, катодной или фиктивной стимуляции, как описано выше.
Сеансы TDCS начинались в фиксированное время утром и вечером с интервалом в 1 неделю между сеансами. Утренние занятия начинались в 8:00, а вечерние — в 18:30; время начала tDCS было запланировано примерно на 8:30 на утренней сессии и 7:00 на вечерней сессии, после подготовительного этапа, который длился ~ 20–30 минут.На каждом сеансе участники удобно сидели в кресле с откидной спинкой, под правой рукой располагалась подушка, а вокруг шеи — вакуумная подушка, чтобы предотвратить движение головы. В начале каждого сеанса измеряли базовую возбудимость коры головного мозга, сначала индуцируя MEP над левым M1, чтобы идентифицировать область, которая произвела наибольшую MEP целевой мышцы (правая ADM) с заданной интенсивностью TMS. Затем область была отмечена, и последующие импульсы подавались из этого оптимального положения.Интенсивность стимуляции регулировали для достижения максимальной амплитуды МВП 1 мВ (SI1 мВ), которая затем использовалась для остальных измерений. После базового измерения 25 MEP было проведено 7 минут анодной, катодной или фиктивной стимуляции, как описано выше. После удаления электродов tDCS, измерения MEP проводились сразу через эпохи каждые 5 минут до 30 минут после tDCS (всего 7 эпох). Этот протокол tDCS дает короткие эффекты последействия, зависящие от полярности, которые исчезают через 30 минут после стимуляции 40 .В конце каждого сеанса участники заполняли опрос о побочных эффектах, в котором предлагалось оценить наличие и тяжесть визуальных явлений, зуда, покалывания, жжения и боли во время стимуляции по шкале Лайкерта от 0 до 5, а также угадать интенсивность стимуляции. полученный (т. е. интенсивность 0 мА или интенсивность 1 мА). Каждый сеанс tDCS занимал около 60 минут.
После удаления электродов tDCS, измерения MEP проводились сразу через эпохи каждые 5 минут до 30 минут после tDCS (всего 7 эпох). Этот протокол tDCS дает короткие эффекты последействия, зависящие от полярности, которые исчезают через 30 минут после стимуляции 40 .В конце каждого сеанса участники заполняли опрос о побочных эффектах, в котором предлагалось оценить наличие и тяжесть визуальных явлений, зуда, покалывания, жжения и боли во время стимуляции по шкале Лайкерта от 0 до 5, а также угадать интенсивность стимуляции. полученный (т. е. интенсивность 0 мА или интенсивность 1 мА). Каждый сеанс tDCS занимал около 60 минут.
Поведенческие меры: моторное обучение и когнитивные задачи
Мы использовали одну конкретную поведенческую задачу для измерения моторного обучения, которое критически затрагивает первичную моторную кору головного мозга, включая LTP-подобную пластичность этой области 17 .Кроме того, участники выполнили три когнитивные задачи для мониторинга рабочей памяти и внимания.
Последовательная задача времени реакции (SRTT)
SRTT — это задача обучения двигательной последовательности. Выполнение этой задачи связано с повышенной активностью и корковой возбудимостью моторных, премоторных и дополнительных моторных областей, а раннее обучение влияет в первую очередь на первичную моторную кору 57,83,84 . В этой задаче последовательное моторное обучение обозначается сокращением времени реакции на нажатие соответствующей кнопки в поле ответа после представления визуально обозначенного стимула на экране компьютера.Участникам предлагается нажать соответствующую кнопку соответствующим пальцем правой руки (указательный палец для кнопки 1, средний палец для кнопки 2, безымянный палец для кнопки 3 и мизинец для кнопки 4). Визуальная подсказка (здесь черная точка) появляется в одном из четырех горизонтальных положений на экране компьютера. Каждое положение на экране соответствует соответствующей кнопке окна ответа. Участникам предлагается как можно быстрее нажимать кнопку, соответствующую положению точек.Продолжительность каждого испытания определяется временем ответа участника (RT), а продолжительность задачи составляет ~ 15–20 минут. В конце каждой пары «стимул – ответ» есть задержка в 500 мс перед представлением следующей реплики. Задача состоит из восьми блоков по 120 испытаний в каждом (всего 960 испытаний). В блоках 1 и 6 последовательность точек следует псевдослучайному порядку таким образом, чтобы точки представлялись одинаково часто в каждой позиции и никогда в одном и том же положении в двух последующих испытаниях. Во время блоков с фиксированной последовательностью (блоки 2–5, 7–8) стимулы появляются в соответствии с фиксированной последовательностью из 12 пунктов, которая повторяется 10 раз (например.г., A – B – A – D – B – C – D – A – C – B – D – C). Усредненная разница RT в блоке 5 (порядок последовательности) по сравнению с блоком 6 (случайный порядок) является основной мерой приобретения моторного обучения, поскольку она указывает ответ на обучение последовательности по сравнению с независимой от обучения характеристикой. Предполагается, что разница RT между блоком 6 (случайный порядок) и блоком 7 (упорядоченный порядок) указывает на дополнительное удержание при обучении. В дополнение к RT, который является основным показателем неявного моторного обучения, вариабельность и точность RT также были рассчитаны в качестве исходных переменных.Участникам не сказали о повторяющейся последовательности, и в конце занятия их спросили, заметили ли они последовательность, и если да, то написать последовательность, чтобы оценить явное изучение задачи. В таких случаях соответствующие данные исключались из окончательного анализа. Две разные последовательности задачи, без перекрывающихся частей и с сопоставимыми трудностями, были представлены на двух сессиях в уравновешенном порядке.
Познавательные задания
Задание на 3 письма .Задача WM в нашем исследовании включала вариант буквенной версии задачи n-back, в которой испытуемые должны были указать, соответствует ли буква, представленная на экране («целевая буква»), букве, представленной ранее («контрольная буква») 85 . Здесь мы использовали 3-заднюю версию задачи 86 , в которой «Попадания» (правильные ответы) определялись как любая буква, идентичная той, которая была представлена тремя попытками назад. Стимулы представляли собой псевдослучайные последовательности из 10 букв (A – J), представленные в фиксированном центральном месте на экране компьютера.Каждая буква была видна в течение 200 мс с интервалом между стимулами 1800 мс, что делало уровень сложности задачи высоким. Буквы были представлены черным цветом на белом фоне и растянуты на 2,4 см (при просмотре с расстояния 50 см от глаз до экрана). Участники выполнили два блока, состоящих из 44 (тренировочный блок) и 143 попыток (основной блок), соответственно, в результате чего общее количество попыток составило 187. Между блоками был предусмотрен короткий перерыв (5–20 с), чтобы участники могли отдохнуть. Две разные версии задачи использовались в двух сессиях (утренняя сессия и вечерняя сессия), а порядок условий был рандомизирован среди участников.Время реакции и меры точности были получены для каждого испытания.
Задача цветного слова Stroop . Задача Струпа по вмешательству — это нейропсихологический тест, широко используемый для измерения избирательного внимания, когнитивного торможения и скорости обработки информации 87,88 . Мы использовали компьютеризированный тест цвета / слова Струпа, аналогичный версии Victoria, основанный на предыдущих исследованиях 88 . В слове Stroop названия цветов были написаны черным и цветом Stroop, некоторые XXX были представлены красными, зелеными, желтыми и синими чернилами, и участники должны были ответить соответствующими клавишами.В задании Струпа по раскрашиванию слов, которое представляло наш особый интерес, участникам предлагаются либо «конгруэнтные», либо «несовместимые» цветные слова. В неконгруэнтных испытаниях цвет чернил, которыми было отображено слово, отличался от значения слова (например, слово «красный» было написано синим), в то время как в конгруэнтных испытаниях и слово, и цвет слова. чернила были идентичны. Стимулы предъявлялись на экране с черным фоном в течение 2000 мс с межстимульным интервалом 500 мс.Размер стимула составлял 1,4 см на расстоянии ~ 50 см от глаза до экрана. Поле ответа с четырьмя клавишами красного, синего, желтого и зеленого цветов помещалось перед испытуемыми, и испытуемые должны были нажимать соответствующую клавишу того цвета, которым было написано слово. Мы увеличили количество конгруэнтных и неконгруэнтных испытаний в задаче «цветное слово» по сравнению с цветовыми блоками Струпа и Струпа, чтобы повысить эффективность анализа ЭЭГ. Всего блок помех Струпа включал 40 совпадающих и 120 неконгруэнтных испытаний, в результате чего в общей сложности было 160 испытаний.
Тест непрерывной работы AX (AX-CPT) . AX-CPT используется для оценки функционирования внимания (устойчивое или временное внимание) или исполнительного контроля, в зависимости от применяемых версий, которые включают базовый уровень, проактивный контроль и реактивный контроль 89,90 . Здесь мы использовали базовую версию задачи, которая короче (~ 15 минут), менее требовательна и измеряет временное внимание 90 . Визуальные стимулы представляли собой белые буквы на темном фоне, появляющиеся по одной на экране компьютера в течение 150 мс каждый с интервалом между стимулами 2000 мс.Испытуемые были проинструктированы нажимать кнопку правым указательным пальцем всякий раз, когда за буквой A (правильная реплика) следовала буква X (правильная цель), как можно быстрее и точнее. Все другие последовательности должны были игнорироваться, включая последовательности, в которых за неправильной репликой (обозначенной «B», но содержащей все буквы, кроме A или X) следовала целевая буква (X), или последовательности, в которых правильная реплика (A ) следовала неверная цель (обозначенная «Y», но содержащая все буквы, кроме A или X).Последовательности AX представлены с высокой вероятностью, чтобы гарантировать сильное смещение ответа. Задания состояли из 240 пар букв (480 проб) с 40% «AX», 40% «BY», 10% «BX» и 10% «AY». Точность и RT были записаны для целевых испытаний.
Процедура
Участники выполняли задания в двух случайно назначенных сеансах утром и вечером в то же время, что и предыдущие занятия, с интервалом не менее 1 недели. Порядок выполнения задач у всех участников был сбалансированным, за исключением SRTT, которая всегда проводилась первой и должна была начаться примерно в то время, когда были применены мониторинг возбудимости коры головного мозга и tDCS.Все задачи (SRTT, N-back, Stroop и AX-CPT) были представлены на экране компьютера (15,6 дюйма, Samsung) с помощью программного обеспечения E-prime 91 , при этом расстояние просмотра от монитора составляло ~ 50 см. Задания проводились в звуконепроницаемом помещении с электромагнитным экраном во время записи ЭЭГ.
ЭЭГ
Запись ЭЭГ
ЭЭГ записывалась непрерывно во время выполнения когнитивной задачи с 30 электродов на голове, расположенных в соответствии с международной системой 10–20, с использованием усилителя ЭЭГ NeurOne Tesla (Bittium, NeurOne, Bittium Corporation, Финляндия) с частотой дискретизации 1000 Гц.Электроды включали: Fp1, Fp2, Fz, F3, F4, F7, F8, Fc1, Fc2, Fc5, Fc6, Cz, C3, C4, T7, T8, Tp9, Tp10, Cp1, Cp2, Cp5, Cp6, Pz, P3, P4, P7, P8, Iz, O1 и O2 и крепились на головке с помощью крышки (EASYCAP GmbH, Herrsching, Германия). Электрод сравнения располагался на FCz, а заземляющий электрод — в положении AFz. Электроды соединяли с головкой с помощью высоковязкого электролитного геля (SuperVisc, Easycap, Herrsching, Германия). Все импедансы поддерживались ниже 10 кОм на протяжении экспериментальных сессий.Данные ЭЭГ были собраны в экранированном помещении и не наблюдались спектральные пики при 50 Гц. Необработанные данные ЭЭГ были записаны и сохранены для автономного анализа с использованием BrainVision Analyzer 2.1 (Brain Products GmbH, Мюнхен, Германия). Запись ЭЭГ включала измерение состояния покоя и состояла из состояний открытых и закрытых глаз, чередующихся каждые 60 с в течение 4 минут, и измерения на основе задачи.
Предварительная обработка и анализ данных ЭЭГ
Записи ЭЭГ были отфильтрованы в автономном режиме полосовым фильтром в диапазоне от 1 до 30 Гц (48 дБ / октава) и повторно привязаны к среднему эталону.Сигнал VEOG через канал Fp2 использовался для обработки артефактов движения глаз в записях ERP с использованием метода Граттона и Коулза 92 , встроенного в BrainVision Analyzer 2.1. Затем данные ЭЭГ были привязаны по времени к появлению интересующего стимула в каждой задаче. Эпохи начинались за 100 мс до начала стимула и заканчивались через 700 мс после начала стимула в SRTT, за 100 мс до начала целевого значения и заканчивались через 1000 мс после начала действия цели в задачах 3-back и AX-CPT и за 100 мс до начала стимула. и закончились через 1000 мс после начала действия стимула в задаче Струпа (как конгруэнтные, так и неконгруэнтные испытания).Эпохи были скорректированы по базовой линии с использованием временного окна -100-0 мс. Артефакты были идентифицированы с использованием комбинации автоматизированного (размах артефактов более 100 мкВ) и ручного процесса отбора. Сегменты были удалены на основе этого автоматического выбора и визуального осмотра для выявления артефактов, связанных с другими источниками ненейрогенной активности. Остальные эпохи были усреднены для расчета среднего ERP. Средняя ERP блоков 5, 6, 7 в задаче SRTT была основана на 120 попытках на блок.В задачах N-back и AX-CPT средний ERP попаданий (правильный ответ) был основан на 40 и 96 испытаниях соответственно. В задаче Струпа средний ERP конгруэнтных и неконгруэнтных испытаний был основан на 40 и 120 испытаниях, соответственно. Для анализа были исследованы следующие усредненные компоненты: (1) P300 на электроде Pz, Cz и P3 во временном окне 250–500 мс после начала стимула в обучающих блоках SRTT (блоки 5, 6, 7), (2) P300 на электроде Fz и Cz во временном окне 300-600 мс 93,94 после начала целевого стимула в задаче 3-back, (3) N200 и N450 на электроде Fz и Cz во временных окнах 200–300 мс и 400–550 мс, соответственно, после начала конгруэнтных и инконгруэнтных испытаний 50 в задаче Струпа и (4) P300 на электродах Fz и Cz в пределах временного окна 300–600 мс 93, 95 после начала действия цели (когда целевой букве X предшествовала метка A) в задаче AX-CPT.Временные окна были выбраны на основе предыдущих исследований и обозначены как максимальное положительное или отрицательное отклонение, возникающее в окне задержки после стимула. Данные состояния покоя ЭЭГ были предварительно обработаны, как описано выше. Здесь данные были разбиты на 2-х эпохи. Анализ с быстрым преобразованием Фурье (длина окна Хеннинга: 10%) был выполнен по эпохам для получения спектральных уровней мощности в бета (13–30 Гц), альфа (7–13 Гц), тета (4–7 Гц) и дельта (1–4 Гц) диапазон.
Статистический анализ
Анализ данных проводился с помощью статистического пакета SPSS, версия 26.0 (IBM, SPSS, Inc., Чикаго, Иллинойс) и GraphPad Prism 8.2.1 (GraphPad Software, Сан-Диего, Калифорния). Рисунки были созданы с помощью Microsoft PowerPoint и Prism 8.2.1. Основные анализы проводились с использованием смешанных моделей ANOVA с факторами как между субъектами, так и внутри них. Был проведен тест сферичности Мочли, и при необходимости применялась поправка Гринхауса – Гейссера. Нормальность и однородность дисперсии данных, собранных с помощью различных измерений, были подтверждены тестами Шапиро – Уилка и Левина соответственно.Размер выборки был предварительно определен с помощью анализа мощности (см. Раздел «Участники»).
Анализ данных корковой возбудимости
Для протоколов TMS с условием двойного импульса (т. Е. SICI-ICF, облегчение I-волны, SAI) полученные средние значения были нормированы на соответствующий тестовый импульс. Сначала были рассчитаны средние значения индивидуально, а затем индивидуальные средние значения были рассчитаны для каждого условия. Для кривых I – O использовались абсолютные значения MEP. Для проверки статистической значимости были выполнены смешанные факторные дисперсионные анализы с повторными измерениями (ISI × дневное время × хронотип) с ISI, интенсивностью TMS (только на кривой I – O) и временем дня (утро или вечер) в качестве факторов внутри субъекта. , хронотип (ЭК против LC) как фактор между субъектами, амплитуда MEP как зависимая переменная и возраст, пол и ИМТ как коварианты.В случае получения значимых результатов дисперсионного анализа, апостериорные сравнения были выполнены с использованием апостериорных тестов Бонферрони t для сравнения средних амплитуд МВП каждого состояния с исходным МВП и для сопоставления предпочтительного циркадного ритма и не предпочтительного циркадного времени в пределах и между группами. Чтобы определить, различались ли условия однократного импульса в течение дня, они были введены в качестве зависимых переменных в смешанный факторный анализ ANOVA с временем дня (утро, вечер) в качестве фактора внутри субъекта и хронотипом (ЭК по сравнению с LC) в качестве межсубъектный фактор.Средние значения условий одиночного импульса (контрольное условие) не различались между утренними и вечерними сессиями для обеих групп в SICI-ICF, фасилитации I-волны и SAI (дополнительная информация).
Анализ данных о нейропластичности, индуцированной tDCS
Размах амплитуды 25 MEP, полученных для каждой временной точки (BL, 0, 5, 10, 15, 20, 25, 30 минут после tDCS), вычисляли и усредняли вместе для каждого состояния tDCS утром и вечером. Чтобы определить, различались ли индивидуальные базовые показатели между сеансами, SI1mV и исходный MEP были введены в качестве зависимых переменных в ANOVA с повторными измерениями с сеансом (шесть уровней) в качестве внутрисубъектного фактора и хронотипом (ЭК против LC) в качестве межсубъектного фактора. .Базовые амплитуды MEP (абсолютные значения) для каждого состояния tDCS достоверно не различались (дополнительная таблица 3). Средняя амплитуда MEP для каждой временной точки измерения была нормализована к исходному уровню сеанса (индивидуальное отношение среднего значения к исходному среднему), в результате чего были получены значения, представляющие либо повышенную (> 1,0), либо пониженную (<1,0) возбудимость. Затем были рассчитаны индивидуальные средние значения нормализованного MEP для каждой временной точки и введены в качестве зависимых переменных в смешанный факторный план с повторными измерениями ANOVA (стимуляция × временная точка × дневное время × хронотип) с условием стимуляции (анодное, катодное, имитационное), временной точкой (восемь уровни), а также время дня (утро или вечер) в качестве внутрисубъектных факторов и хронотип (ЭК против LC) в качестве межсубъектного фактора, а также возраст, пол и ИМТ в качестве ковариант.В случае значимых результатов ANOVA, апостериорные сравнения амплитуд МВП в каждый момент времени выполнялись с использованием апостериорных тестов Бонферрони t , чтобы проверить, привела ли активная стимуляция к значительному различию по сравнению с фиктивным (сравнение 1), исходным (сравнение 2), соответствующие условия стимуляции в предпочтительные для циркадных ритмов по сравнению с не предпочтительными для циркадных ритмов (сравнение 3) и сравнения между группами в соответствующие моменты времени (сравнение 4).
Анализ поведенческих данных
Были рассчитаны средние значения RT и точности для блоков 5, 6 и 7 SRTT.Испытания с ошибочными ответами, а также испытания с RT <150 мс 96,97 или> 3000 мс, а также испытания, которые отклонялись на 3 или более стандартных отклонения от среднего индивидуального времени ответа, были отброшены 98 . Один участник мог определить последовательность стимулов, и соответствующие данные были исключены из анализа. Средние RT были стандартизированы для блока 1 для каждого субъекта в каждый момент измерения отдельно. Стандартное отклонение RT для каждого предмета и учебного блока рассчитывалось как индекс изменчивости RT.Среднее значение RT, вариабельность RT и точность блоков были введены в качестве зависимых переменных в смешанный факторный анализ ANOVA с повторными измерениями (блок × дневное время × хронотип) с блоками (5, против 6, 6 против 7) и временем дня (утро против вечером) как внутрисубъектные факторы и хронотип (ЭК против LC) как межсубъектный фактор. Поскольку различия RT между блоками 5 против 6 и 6 против 7 представляли большой интерес, апостериорные сравнения были выполнены на разнице RT между этими блоками с использованием тестов с парной выборкой t (двусторонний, P <0.05) без поправки на множественные сравнения. Для задач 3-back, Stroop и AX-CPT, среднее значение и стандартное отклонение RT и точности были рассчитаны и введены в качестве зависимых переменных в смешанный факторный анализ ANOVA (дневное время × хронотип) с указанием времени дня (утро или вечер) и хронотипа. как внутрисубъектные и межсубъектные факторы соответственно. Возраст, пол и ИМТ были введены как ковариаты во всех ANOVA. Для получения значимых результатов дисперсионного анализа, апостериорные сравнения зависимых переменных по времени дня (утро и вечер) были выполнены с использованием парных тестов t (двусторонний, P <0.05) без поправки на множественные сравнения, поскольку мы сравнивали только два условия.
Корреляционный анализ
Чтобы оценить взаимосвязь между индуцированной нейропластичностью и обучением двигательной последовательности, а также взаимосвязь между корковой возбудимостью и выполнением когнитивных задач, мы использовали двумерный линейный регрессионный анализ (корреляция Пирсона, односторонняя). Для первой корреляции мы использовали отдельные большие усредненные амплитуды МВП, полученные из анодных и катодных tDCS, объединенных для моментов времени от 0 до 20 минут после вмешательства, которые показали пластические реакции во всех условиях в предпочтительное время дня и индивидуальные показатели двигательного обучения. полученные на основе различий RT в блоке 5 и 6 как для предпочтительного циркадного ритма (утро для EC, вечер для LC), так и для не предпочтительного (вечер для EC, утро для LC) времени.Для второй корреляции мы использовали отдельные большие усредненные амплитуды MEP, полученные из каждого протокола TMS, и индивидуальную точность / RT, полученную для каждой задачи как в предпочтительные для циркадных ритмов, так и в нежелательные моменты времени.
Анализ данных ЭЭГ
Предварительная обработка и анализ данных ЭЭГ были описаны в предыдущем разделе. Для всех задач индивидуальные средние ERP были усреднены и введены как зависимые переменные в смешанный факторный анализ ANOVA (дневное время × хронотип) со временем дня (утро или вечер) как внутри субъекта и хронотипом (ЭК против LC) как между субъектами. факторы.Коррекция не использовалась для исследования нескольких положений электродов. Данные ЭЭГ одного участника в задаче Струпа, которые не были записаны должным образом из-за технических трудностей, и другого участника, чьи поведенческие данные SRTT были исключены, были отброшены.
Сводка отчетов
Дополнительная информация о дизайне исследования доступна в Сводке отчетов по исследованиям природы, связанной с этой статьей.
Границы | Конкуренция, сотрудничество и механизмы умственной деятельности
Инструкция, отбор и сотрудничество
Понимание природы психических функций представляет собой одну из самых серьезных проблем для современной нейробиологии.Хотя точный способ, которым различные мозговые структуры активируют когнитивные процессы более высокого порядка (тщательный путь, ведущий «от молекул к разуму»), все еще остается загадкой, ряд влиятельных моделей психических функций подчеркивают важность конкуренции между нейронными цепями как один из основных механизмов поведенческой гибкости и изощренности, проявляемых когнитивными процессами более высокого порядка. Такие концепции, как «избирательная стабилизация синапсов» (Changeux et al., 1973; Changeux and Danchin, 1976) и «нейронный дарвинизм» (Edelman, 1993), которые можно объединить в семейство объяснительных моделей, основанных на стратегиях вариации / отбора, попытайтесь понять, как взаимодействие с окружающей средой влияет на нейронную архитектуру, изменяя тонкую структуру синаптических связей.
Нельзя недооценивать важность избирательных механизмов для понимания психических процессов. Однако в этих моделях иногда трудно определить точные единицы репликации (особенно если кто-то хочет придерживаться строго дарвиновской парадигмы) и критерии отбора, используемые мозгом, по крайней мере, в отсутствие репродуктивных аргументов, представленных в стандартный биологический дарвинизм. Кроме того, несмотря на широко распространенное отсутствие предварительной спецификации и высокую степень универсальности, которая может быть отнесена к нейронным группам в модели Эдельмана, ясно, что корковые функции во многих случаях расположены в определенных областях (Blanco Pérez, 2017).Следовательно, проблема сочетания локализации и интеграции все еще остается в значительной степени нерешенной.
Кроме того, на теоретическом уровне также необходимо учитывать эффекты сотрудничества между нейронными цепями и то, каким образом этот процесс связан с конкуренцией. Конечно, решающая роль сотрудничества также признается наиболее влиятельными селективными моделями когнитивной функции, в которых сочетание генетики и отбора часто осуществляется через кооперативные стратегии, соединяющие различные уровни организации мозга в соответствии с адаптивными последствиями. определенного поведения.Тем не менее, стоит рассмотреть некоторые важные особенности механизмов сотрудничества, которые могут указывать на их концептуальную независимость.
Специфичность нейронных связей не может быть объяснена ни биохимическим соответствием один-к-одному, происходящим из генетической программы, ни избирательной стабилизацией, происходящей в результате приобретения нового опыта в часто непредсказуемой среде. Хотя этих процессов может быть достаточно, чтобы оправдать существование и преобладание определенных маршрутов обработки, они не могут объяснить, как информация интегрируется в единое восприятие.Если, в очень упрощенных терминах, генетические и селективные пути учитывают степень специализации, проявляющуюся в некоторых нейронных процессах, в которых сенсорная информация тщательно различается, на некоторых уровнях организации мозга они также должны отдавать предпочтение кооперативным стратегиям, которые вносят дополнительные степени изменчивости или «Неспециализация». Эта гибкость будет лежать в основе способности объединять разнородные черты в связное перцептивное целое, способность, которая, по-видимому, составляет одну из фундаментальных характеристик психической функции.
Кооперацию можно понять с точки зрения различных механизмов клеточной коммуникации между группами нейронов и более крупными сетями. Одним из них является так называемое «обучение Хебба», согласно которому синаптические связи укрепляются, когда задействованные нейроны имеют сильно коррелированные выходы. На уровне нейронов этот процесс влечет за собой возможность того, что схемы, обрабатывающие информацию конвергентной, но разнородной природы, могут создавать нейронные сети. Связь между цепями помогает нам понять объединение различных свойств объектов в единое восприятие.Возможность коммуникации, вероятно, лучше всего определяется физическими аспектами, такими как близость между схемами, которые тематически организованы, природа самой сенсорной системы и способность определенных нейронных сетей создавать общие паттерны активности, которые облегчают конвергенцию между различными видами Информация. Таким образом, гипотетически мы можем ожидать, что как близость в пространстве, так и временная синхронизация вызовут определенные паттерны проводки в нейронных сетях. Природа и свойства стимула воплощены в самих путях, то есть в структурной организации конкретной сенсорной системы.
Такой процесс интеграции должен происходить в разных масштабах. Вообще говоря, первый уровень определяется способностью определенного сенсорного пути извлекать информацию из стимула. На второй, многоуровневой стадии объединение произойдет внутри сенсорной системы в целом (например, визуальной системы), чтобы получить единое восприятие в рамках этой сенсорной модальности. Но все же возможно теоретизировать о более высоком уровне интеграции: перцептивном целом, в котором различные сенсорные модальности сходятся в единое целое (тайна интеграции, элегантно резюмированная Карлом Лэшли: «как [это так]] специализированные области коры головного мозга взаимодействуют, чтобы произвести интеграцию, очевидную в мышлении и поведении ») (Lashley, 1931).Эта способность связана с широким диапазоном связи, особенно если принять во внимание, что «никакая область коры головного мозга не соединяется только с одной или двумя другими областями» (Zeki and Shipp, 1988).
Если селективные стратегии анализируют и разлагают характеристики стимула посредством функциональной дифференциации и иерархического разделения труда, горизонтальные процессы помогают синтезировать эту информацию в единое целое. Говоря более абстрактно, механизмы функциональной дифференциации представляют собой успешные эволюционные стратегии, позволяющие справляться со сложностями окружающей среды.Однако те процессы, которые стремятся объединить эти различия в пространстве и времени в единое восприятие, являются необходимым противовесом опасностям, связанным с растущей степенью специализации, которая может препятствовать восприятию реальности в целом.
Следовательно, с этой точки зрения функциональная логика корковых связей подразумевает сосуществование генетически детерминированных, конкурентных и кооперативных процессов, построенных на действующих принципах различной концептуальной природы.Окончательные паттерны разводки, которые определяют нейронные связи, должны затем описываться по крайней мере тремя классами процессов: генетическими механизмами, которые инструктируют создание определенных нейронных сборок (через генные сети, которые кодируют нейронные сети; Greenspan, 2009), избирательными эпигенетическими стратегиями, которые следуют далее. нейронная активность и изменение формы ранее сформированных сетей, а также кооперативный эпигенез, основанный на возможности связи между различными сетями. Эта комбинация обучения, отбора и сотрудничества дает мозгу изобилие конфигурационных возможностей и может рассматриваться как существенная основа его поведенческой универсальности.
Некоторые сенсорные системы
Как хорошо известно, визуальная информация передается по разным маршрутам, каждый из которых достиг выдающейся степени функциональной специализации в обработке определенных характеристик зрительного стимула, таких как движение, форма и цвет (Zeki and Shipp, 1988). Поэтому принцип функциональной специализации зрительной коры головного мозга имеет первостепенное значение для понимания природы восприятия. Информация, касающаяся цвета, обрабатывается в области V4, тогда как информация о движении относится к системе V5, так что повреждение в первой области (например, ахроматопсия) может не повлиять на вторую систему (одной из самых известных патологий которой является акинетопсия; Zeki, 2003 ).
Специализация, рассматриваемая как разделение нейронной работы между группами корковых нейронов, регулируется иерархической обработкой информации, при которой возможно достижение растущих уровней абстракции (или «формализации») при усвоении стимула. Таким образом, в восходящих путях функциональная дифференциация сосуществует с кооперацией между различными группами клеток.
Учитывая, что информация в сенсорных системах обрабатывается параллельно и иерархически через ретрансляционные клетки, функциональная дифференциация гарантируется через таламо-кортикальную и кортико-корковую связь.От сенсорной периферии до высших мозговых центров (Guillery and Sherman, 2002) сложные клетки, такие как пирамидные нейроны большого размера, контролируют активность групп более простых нервных клеток в релейной цепи, проходящей через звездчатые клетки в слое 4C, ганглиозные клетки и т. Д. и фоторецепторы.
Таким образом, функциональность, демонстрируемая визуальной системой, обеспечивается некоторыми примечательными анатомическими особенностями. Среди них стоит отметить его иерархическую организацию (которая способствует возрастающей степени абстрагирования от содержания стимула), наличие параллельных маршрутов (каждый из которых приобрел важные уровни специализации и стратегий разделения и императора , в то время как на в то же время гарантируя конвергенцию между этими дифференцированными путями), топографическую организацию системы и существование вертикальных столбов нервных клеток, которые собирают нейроны, наделенные сходными свойствами (Hubel and Wiesel, 1968).Аналогичным образом, анализ таких структурных характеристик необходимо объединить с анализом активности нейронных цепей, действующих в этой сенсорной системе, в частности, с ответами нейронов обратной связи через реентерабельные или рекурсивные связи.
Объединение психических функций
На этом этапе можно оценить необходимость сочетания изучения основных молекулярных механизмов с подходом, вдохновленным системной нейробиологией, так что восходящие и нисходящие процессы могут сходиться в единую картину мозга и связанных с ним функций.В самом деле, кажется очевидным, что понимание мозга в целом требует интеграции редукционистской и холистической структур. Например, расшифровка фундаментальных химических механизмов дает возможность постепенно раскрыть обширную нить, ведущую от генов, нейронов, мессенджеров, глии и синапсов к психической функции в целом.
В этом динамическом взаимодействии между унаследованными и приобретенными элементами информации и, более того, между специфичностью и пластичностью (Merzenich et al., 1988), с помощью которых деятельность и опыт могут реконструировать церебральную функцию, можно выделить одну из самых мощных адаптивных функций мозга. В то время как фундаментальные нейроанатомические структуры могут обладать высокой степенью стабильности (определяемой генетической программой инструкций, применяемой с раннего и постнатального развития до критических периодов), функциональная взаимосвязь демонстрирует замечательный уровень пластичности. Равновесие между структурной стабильностью и функциональной изменчивостью достигается посредством процесса постоянной регуляции, при которой нервные клетки способны производить последовательные паттерны активности, восприимчивые к модификации в соответствии с новым опытом, накопленным субъектом.Происходящие из поучительных механизмов, восходящие причинно-следственные связи можно интерпретировать как ограничения, ограничивающие общую изменчивость системы; однако наличие нисходящих процессов позволяет усилить его изменчивость в пределах, установленных этими ограничениями.
Отсутствие предварительной спецификации в значительном количестве нейронных соединений предлагает мощный инструмент для адаптации к изменяющейся среде. Вместо того, чтобы создавать жесткие паттерны, которые связывают нейробиологические структуры и функции в уникальных формах, мозг разработал эффективный способ справляться с проблемами окружающей среды с помощью очень гибкой репрезентативной архитектуры (Dehaene-Lambertz et al., 2002).
Таким образом, сложное взаимодействие между, с одной стороны, локализацией (подход, который склонен рассматривать умственную деятельность как по существу кодифицированную в деятельности определенных нейронных связей) и, с другой стороны, интеграцией (согласно которой поведение и умственная деятельность проистекают из интегрированной деятельности мозга) необходимо обращать внимание на процессы функциональной специализации, опосредованные избирательными механизмами, и процессы функциональной интеграции.Последнее требует механизмов сотрудничества между нейронными сетями, многие из которых действительно могут быть переданы через определенные органы, отвечающие за соединение физически разделенных областей мозга или отдаленных зон в пределах определенной области.
Эту модель ISC (команда-выбор-сотрудничество) можно резюмировать как комбинацию инструкций, способных создать стабильную, но относительно гибкую нейронную архитектуру, выбор через взаимодействие с окружающей средой и сотрудничество, которые будут играть фундаментальную роль в продвижении унификация психических функций видна в интегрированном характере многих восприятий и поведения.Таким образом, развертывание тонкой нити Ариадны, которая соединяет самые элементарные нейробиологические структуры с высшими когнитивными функциями, потребует надлежащей интеграции этих трех основных классов процессов. С метафорической точки зрения, «эгоистичные схемы» должны сосуществовать с «альтруистическими сетями».
Авторские взносы
Автор подтверждает, что является единственным соавтором данной работы, и одобрил ее к публикации.
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Бланко Перес, К. (2017). «Философия, нейробиология и дар творчества», в Argumenta Philosophica: Revista de la Encyclopedia Herder , ed. Э. Гердер (Барселона: Гердер), 95–108.
Google Scholar
Changeux, J. P., Courrége, P., and Danchin, A. (1973). Теория эпигенеза нейронных сетей путем избирательной стабилизации синапсов. Proc. Natl. Акад. Sci. США 70, 2974–2978 DOI: 10.1073 / pnas.70.10.2974
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Changeux, J. P., and Danchin, A. (1976). Избирательная стабилизация развивающихся синапсов как механизм спецификации нейронных сетей. Природа 264, 705–712. DOI: 10.1038 / 264705a0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гринспен, Р. Дж. (2009). «Отбор, взаимодействие генов и гибкие генные сети», в Колд-Спринг-Харбор Симпозиумы по количественной биологии (Колд-Спринг-Харбор, Нью-Йорк, Нью-Йорк: Лаборатория Колд-Спринг-Харбор Пресс).
Google Scholar
Гильери, Р. В., и Шерман, С. М. (2002). Функции таламических реле и их роль в кортикокортикальной коммуникации: обобщения визуальной системы. Нейрон 33, 163–175. DOI: 10.1016 / S0896-6273 (01) 00582-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мерзенич, М. М., Реканзоне, Г., Дженкинс, В. М., Аллард, Т. Т., и Нудо, Р. Дж. (1988). Корковая репрезентативная пластичность. Neurobiol.Neocortex 41–67.
PubMed Аннотация | Google Scholar
Отображение мозга
Головной мозг, самая большая часть человеческого мозга, связан с функционированием более высокого порядка, включая контроль произвольного поведения. Мышление, восприятие, планирование и понимание языка — все это находится под контролем головного мозга.
На верхнем изображении показаны четыре основных отдела коры головного мозга: лобная доля, теменная доля, затылочная доля и височная доля.Такие функции, как движение, контролируются моторной корой, а сенсорная кора получает информацию о зрении, слухе, речи и других органах чувств. На нижнем изображении показано расположение основных внутренних структур мозга.
Головной мозг разделен на два полушария — правое полушарие и левое полушарие. Эти два полушария соединяет пучок волокон, называемый мозолистым телом. Два полушария сообщаются друг с другом через мозолистое тело.
Внешний слой головного мозга покрывает слой ткани, называемый корой головного мозга.Из-за серого цвета кору головного мозга часто называют серым веществом. Морщинистый вид человеческого мозга также можно отнести к характеристикам коры головного мозга. Более двух третей этого слоя загибают в бороздки. Канавки увеличивают площадь поверхности мозга, что позволяет включать гораздо больше нейронов.
Функцию коры головного мозга можно понять, условно разделив ее на зоны, как географическое расположение континентов.
Лобная доля отвечает за инициирование и координацию двигательных движений; более высокие когнитивные навыки, такие как решение проблем, мышление, планирование и организация; и для многих аспектов личности и эмоционального макияжа.
Теменная доля отвечает за сенсорные процессы, внимание и язык. Повреждение правой стороны теменной доли может привести к затруднениям в перемещении по пространствам, даже знакомым. Если травмирована левая сторона, способность понимать устную и / или письменную речь может быть нарушена.
Затылочная доля помогает обрабатывать визуальную информацию, включая распознавание форм и цветов.
Височная доля помогает обрабатывать слуховую информацию и интегрировать информацию от других органов чувств. Нейробиологи также считают, что височная доля играет роль в кратковременной памяти через формирование гиппокампа и в приобретенных эмоциональных реакциях через миндалину.
Все эти структуры составляют передний мозг. Другие ключевые части переднего мозга включают базальные ганглии, которые представляют собой ядра головного мозга глубоко в коре головного мозга; таламус; и гипоталамус.Ядра головного мозга помогают координировать движения мышц и поощряют полезное поведение; таламус передает большую часть сенсорной информации коре головного мозга после того, как помогает расставить приоритеты; гипоталамус — это центр управления аппетитом, защитным и репродуктивным поведением, а также бодрствованием во сне.
Средний мозг состоит из двух пар небольших холмов, называемых бугорками. Эти наборы нейронов играют решающую роль в зрительных и слуховых рефлексах и в передаче этого типа информации в таламус.В среднем мозге также есть кластеры нейронов, которые регулируют активность в обширных частях центральной нервной системы и считаются важными для механизмов вознаграждения и настроения.
Задний мозг включает мост и продолговатый мозг, которые контролируют дыхание, сердечный ритм и уровень глюкозы в крови.
Другая часть заднего мозга — мозжечок, который, как и большой мозг, также имеет два полушария. Два полушария мозжечка помогают контролировать движения и когнитивные процессы, которые требуют точного определения времени, а также играют важную роль в обучении по Павлову.
Спинной мозг — это продолжение головного мозга через позвоночный столб. Он получает сенсорную информацию от всех частей тела ниже головы. Он использует эту информацию, например, для рефлекторных реакций на боль, а также передает сенсорную информацию в мозг и его кору головного мозга. Кроме того, спинной мозг генерирует нервные импульсы в нервах, которые контролируют мышцы и внутренние органы, как посредством рефлекторной активности, так и посредством произвольных команд из головного мозга.
Реорганизация крупномасштабных мозговых сетей в детстве и подростковом возрасте
лица «обрезаются» в раннем детстве. «Cerebral »Cortex, №21, №191–199.
Цао, X., Cao, Q., Long, X., Sun, L., Sui, M., Zhu, C., … Wang, Y . (2009).«Аномальные» состояния покоя »функциональные
« паттерны »связи между« путаменом »наивных-наивных детей с дефицитом внимания и гиперактивностью.
BrainResearch, №1303, №195–206.
Casey, B.J., Soliman, F., Bath, K.G., & Glatt, C.E. (2010). Imaginggeneticsand development: Challengesи
обещания. HumanBrainMapping, 31, 838–851
Chai, X.J., Ofen, N., Jacobs, L. F., & Gabrieli, J.D.E. (2010). «Сцена» сложности: «Влияние» на восприятие, «память»,
и «развитие» во «медиальном» временном lobe.«Границы» в «Человеческой неврологии», №4, №21, №1–10.
Chiang, M.C., Barysheva, M., Shattuck, D.W., Lee, A.D., Madsen, S.K., Avedissian , C., … Thompson, P.M. (2009).
Генетическая структура мозгового волокна и интеллектуальная производительность. Журнал неврологии, №29, №2212–2224.
Christakou, A., Halari, R., Smith, A.B., Ifkovits, E., Brammer, M., И Rubia, K. (2009 г.) Зависимая от секса возрастная модуляция
лобно-стриатной и височно-теменной активации во время когнитивного контроля.«Нейроимидж», №48, №223–236.
Церковь, J.A., Fair, D.A., Dosenbach, N.U., Cohen, A.L., Miez in, F.M ., Petersen, S.E., & Schlaggar, B.L. (2009).
Контрольные сети в педиатрических условиях синдрома Туретта демонстрируют незрелые и аномальные модели функционального соединения
. «Мозг», №132, №225–238.
Crone, E., Wendelken, C., Donohue, S., VanLeijenhorst, L., & Bunge, S.A. (2006). разработка
способности манипулировать информацией в рабочей памяти.«Труды» Национальной академии наук США,
103, №9315–9320.
Дэвид, О., Гиймен, И., Сейллет, С., Рейт, С., Дерансарт, К., Сегебарт, О. . »(2008).« Идентификация »нейронных
драйверов» с «функциональной» МРТ: «Электрофизиологическая» валидация. «PLoS» Биология, №6, №2683–2697.
Davidson, M.C., Amso, D., Anderson, L.C., & Diamond, A. (2006). и исполнитель
функций от4 до13лет: «Доказательство» манипуляций с памятью, «запрещение», «переключение задач».
Neuropsychologia, №44, №2037–2078.
Dehaene-Lambertz, G., Montavont, A., Jobert, A., Allirol, L., Dubois, J., Hertz-Pannier, L., & Dehaene, S. (2010).
Язык — или музыка, «мать» или «Моцарт?» — структурное и экологическое влияние на детские языковые сети.
BrainandLanguage, 114, 53–65.
Depue, B.E., Burgess, G.C., Willcutt, E.G., Ruzic, L., & Banic h, M.T. (2010). «Тормозящий» контроль »извлечения« памяти »
и« моторной »обработки, связанной с« правой »боковой» префронтальной »корой:« Доказательства »дефицита» у людей »с
СДВГ.«Нейропсихология», №48, №3909–3917.
Depue, B.E., Curran, T., & Banic h, M.T. (2007). Предфронтальные регионы организуют подавление эмоциональных воспоминаний
через двухфазный процесс. Наука, №37, №215–219.
Dosenbach, N.U., Nardos, B., Cohen, A.L., Fair, D.A., Power, J.D., Church , J.A., … Schlaggar, B.L. (2010).
Прогнозирование индивидуальной зрелости мозга с помощью МРТ. Наука, №329, №1358–1361.
Durston, S., Davidson, M.C., Tottenham, N., Galvan, A., Spic er, J., Fossella, J.A., & Casey, B.J. (2006). Ashift с
диффузный на фокальный кортикальный «Деятельность» с развитием. «Развивающая» наука, №9, №1–8.
Emmerich, H.J., & Ackerman, B.P. (1978). Различия в развитии напомним: Кодированиеилиполучение? «Детская» Психология, №25, №514–525.
Fair, D.A., Cohen, A.L., Power, J.D., Dosenbach, N.U., Church, J.A. , Miezin, F.M., … Petersen, S.E. (2009).
Функциональные «мозговые» сети развиваются на основе «локальной — распределенной» организации.PLoSComputationalBiology, 5,
e1000381.
FanJ, FossellaJ, SommerT, Yanghong, W., & Posner »(2003). Отображение« генетической вариации »исполнительного внимания
на» деятельность мозга. «Материалы» Национальной академии наук США, №100, №7406–7411.
05-24_HPY_075783.indd
% PDF-1.4 % 1 0 объект > эндобдж 7 0 объект /Заголовок / Автор /Режиссер / Ключевые слова / CreationDate (D: 20211118111050-00’00 ‘) / ModDate (D: 20080207114531Z) >> эндобдж 2 0 obj > эндобдж 3 0 obj > эндобдж 4 0 obj > эндобдж 5 0 obj > эндобдж 6 0 obj > транслировать
! Z̭K7,> B.0 ~} ljC; Cʳ40 @ zDv6LJ, cȁN} e & fRҸӕ `’u; pOm8, F1Y1GNwabВŇZLCNP8 뻣 M @ ĉ -, / icT — {\ bbK 7TR 3} z {-NsDHQ / ݳ * 9} + euŭ ‘b | ғWb% l% sL \ -% T`) 96FD¸oJ) ; o, NV {UjUo.v } f2Z
| [1] | Джеймс У. (1890) Принципы психологии (Х.Холт, Нью-Йорк, штат Нью-Йорк). |
| [2] | Познер М.И., Петерсен С.Е. (1990) Система внимания человеческого мозга. Анну Рев Neurosci 13: 25-42. DOI: 10.1146 / annurev.ne.13.030190.000325 |
| [3] | Норман Д.А., Шаллис Т. (1986) Внимание к действию: волевое и автоматическое управление поведением. Сознание и саморегуляция , ред. Дэвисон Р.Дж., Шварц Г.Е., Шапиро Д. (Plenum Press, Нью-Йорк, Нью-Йорк), стр. 1-18. |
| [4] | Корбетта М., Шульман Г.Л. (2002) Контроль целенаправленного и стимулированного внимания в мозге. Nat Rev Neurosci 3: 201-215. |
| [5] | Познер М.И., ДиДжироламо Г.Дж. (1998) Внимание руководителей: конфликт, обнаружение целей и когнитивный контроль. Внимательный мозг , изд. Парасураман Р. (MIT Press, Кембридж, Массачусетс), стр. 401-423. |
| [6] | Д’Анджело М.С., Милликен Б., Хименес Л. и др. (2013) Реализация гибкости в автоматизме: данные контекстно-зависимого неявного последовательного обучения. Сознание 22: 64-81. DOI: 10.1016 / j.concog.2012.11.002 |
| [7] | Руэда М.Р., Познер М.И., Ротбарт М.К. (2005) Развитие исполнительного внимания: вклад в появление саморегулирования. Dev Neuropsychol 28: 573-94. DOI: 10.1207 / s15326942dn2802_2 |
| [8] | Луу П., Такер Д.М., Деррибери Д. и др. (2003) Электрофизиологические реакции на ошибки и обратная связь в процессе регуляции действий. Psychol Sci 14: 47-53. DOI: 10.1111 / 1467-9280.01417 |
| [9] | Dagenbach D, Carr TH (1994) Тормозные процессы внимания, памяти и языка (Academic Press, Сан-Диего, Калифорния). |
| [10] | Петерсен С.Е., Познер М.И. (2012) Система внимания человеческого мозга: 20 лет спустя. Annu Rev Neurosci 35: 73-89. DOI: 10.1146 / annurev-neuro-062111-150525 |
| [11] | Познер М.И., Руэда М.Р., Канске П. (2007) Исследование механизмов внимания. Handb Psychophysiol : 410-432. |
| [12] | Фан Дж., МакКэндлисс Б.Д., Соммер Т. и др. (2002) Проверка эффективности и независимости сетей внимания. J Cogn Neurosci 14: 340-347. DOI: 10.1162 / 0898927361886 |
| [13] | Каллехас А., Лупианес Дж., Фунес М.Дж. и др.(2005) Модуляции среди сетей оповещения, ориентирования и исполнительного контроля. Exp brain Res 167: 27-37. DOI: 10.1007 / s00221-005-2365-z |
| [14] | Fan J, Gu X, Guise KG и др. (2009) Тестирование поведенческого взаимодействия и интеграции сетей внимания. Brain Cogn 70: 209-220. DOI: 10.1016 / j.bandc.2009.02.002 |
| [15] | Hackley SA, Valle-Inclán F (2003) Какие этапы обработки ускоряются с помощью предупреждающего сигнала? Biol Psychol 64: 27-45. DOI: 10.1016 / S0301-0511 (03) 00101-7 |
| [16] | Вайнбах Н., Хеник А. (2012) Взаимосвязь между бдительностью и исполнительным контролем. J Exp Psychol Hum Percept Perform 38: 1530-1540. DOI: 10.1037 / a0027875 |
| [17] | Посуэлос Дж. П., Пас-Алонсо П. М., Кастильо А. и др. (2014) Развитие сетей внимания и их взаимодействия в детстве. Dev Psychol 50: 2405-2415. DOI: 10.1037 / a0037469 |
| [18] | Fox MD, Corbetta M, Snyder AZ и др.(2006) Спонтанная нейрональная активность различает дорсальную и вентральную системы внимания человека. Proc Natl Acad Sci U S A 103: 10046-10051. DOI: 10.1073 / pnas.0604187103 |
| [19] | Dosenbach NUF, Fair DA, Miezin FM и др. (2007) Отдельные сети мозга для адаптивного и стабильного управления задачами у людей. Proc Natl Acad Sci U S A 104: 11073-11078.DOI: 10.1073 / pnas.0704320104 |
| [20] | Познер М.И., Ротбарт М.К. (2007) Исследование сетей внимания как модели интеграции психологической науки. Annu Rev Psychol 58: 1-23. DOI: 10.1146 / annurev.psych.58.110405.085516 |
| [21] | Coull JT, Nobre AC, Frith CD (2001) Норадренергический агонист α2 клонидин модулирует поведенческие и нейроанатомические корреляты ориентации внимания и предупреждения человека. Cereb Cortex 11: 73-84. DOI: 10.1093 / cercor / 11.1.73 |
| [22] | Aston-Jones G, Cohen JD (2005) Интегративная теория функции голубого пятна и норэпинефрина: адаптивное усиление и оптимальная производительность. Annu Rev Neurosci 28: 403-450. DOI: 10.1146 / annurev.neuro.28.061604.135709 |
| [23] | Coull JT, Frith CD, Frackowiak RSJ, Grasby PM (1996) Лобно-теменная сеть для быстрой обработки зрительной информации: ПЭТ-исследование устойчивого внимания и рабочей памяти. Neuropsychologia 34: 1085-1095. DOI: 10.1016 / 0028-3932 (96) 00029-2 |
| [24] | Цуй Р.К., Эгхер А., Хутер Д. и др. (2000) Пространственно-временной анализ с высоким разрешением случайной отрицательной вариации в простых или сложных моторных задачах и немоторных задачах. Clin Neurophysiol 111: 1847-1859. DOI: 10.1016 / S1388-2457 (00) 00388-6 |
| [25] | Coull JT (2004) исследования временного внимания с помощью фМРТ: распределение внимания во времени или в направлении времени. Cogn Brain Res 21: 216-226. DOI: 10.1016 / j.cogbrainres.2004.02.011 |
| [26] | Хиллард С.А. (1985) Электрофизиология избирательного внимания человека. Trends Neurosci 8: 400-405. DOI: 10.1016 / 0166-2236 (85) -0 |
| [27] | Mangun GR, Hillyard SA (1987) Пространственное распределение визуального внимания, индексируемое связанными с событиями потенциалами мозга. Факторы шума 29: 195-211. |
| [28] | Desimone R, Duncan J (1995) Нейронные механизмы избирательного зрительного внимания. Анну Рев Neurosci 18: 193-222. DOI: 10.1146 / annurev.ne.18.030195.001205 |
| [29] | Корбетта М., Патель Г., Шульман Г.Л. (2008) Система переориентации человеческого мозга: от окружающей среды к теории разума. Нейрон 58: 306-324. |
| [30] | Greicius MD, Krasnow B, Reiss AL, et al.(2003) Функциональная связь в покоящемся мозгу: сетевой анализ гипотезы режима по умолчанию. Proc Natl Acad Sci U S A 100: 253-258. DOI: 10.1073 / pnas.0135058100 |
| [31] | Мантини Д., Перруччи М.Г., Дель Гратта С. и др. (2007) Электрофизиологические сигнатуры сетей состояния покоя в человеческом мозге. Proc Natl Acad Sci U S A 104: 13170-13175. DOI: 10.1073 / pnas.0700668104 |
| [32] | Визинтин Э, Де Панфилис С., Антонуччи С. и др. (2015) Анализ внутренних сетей, лежащих в основе внимания: исследование состояния покоя. Behav Brain Res 278: 315-322. DOI: 10.1016 / j.bbr.2014.10.002 |
| [33] | Умарова Р.М., Саур Д., Шнелл С. и др.(2010) Структурная связь для зрительно-пространственного внимания: значение вентральных путей. Cereb Cortex 20: 121-129. DOI: 10.1093 / cercor / bhp086 |
| [34] | Бушман Т.Дж., Миллер Е.К. (2007) Контроль внимания сверху вниз и снизу вверх в префронтальной и задней теменной коре. Sci 315: 1860-1862.DOI: 10.1126 / science.1138071 |
| [35] | Vossel S, Geng JJ, Fink GR (2013) Дорсальные и вентральные системы внимания: отдельные нейронные цепи, но совместные роли. Neurosci 20: 150-159. |
| [36] | Он Б.Дж., А.З. Снайдер, Дж. Л. Винсент и др.(2007) Нарушение функциональной связи в лобно-теменных сетях лежит в основе поведенческих дефицитов при пространственном пренебрежении. Нейрон 53: 905-918. DOI: 10.1016 / j.neuron.2007.02.013 |
| [37] | Giesbrecht B, Weissman DH, Woldorff MG, et al. (2006) Предварительно заданная активность в зрительной коре головного мозга позволяет прогнозировать поведенческие характеристики при выполнении задач на пространственное и особенное внимание. Brain Res 1080: 63-72. DOI: 10.1016 / j.brainres.2005.09.068 |
| [38] | Geng JJ, Mangun GR (2011) Активация правого височно-теменного соединения ярким контекстным сигналом облегчает распознавание цели. Neuroimage 54: 594-601. DOI: 10.1016 / j.neuroimage.2010.08.025 |
| [39] | Fan J, Flombaum JI, McCandliss BD, et al.(2003) Познавательные и мозговые последствия конфликта. Neuroimage 18: 42-57. DOI: 10.1006 / nimg.2002.1319 |
| [40] | Буш Г., Луу П., Познер М. И. (2000) Когнитивные и эмоциональные влияния в передней поясной коре головного мозга. Trends Cogn Sci 4: 215-222. DOI: 10.1016 / S1364-6613 (00) 01483-2 |
| [41] | Drevets WC, Raichle ME (1998) Взаимное подавление регионального мозгового кровотока во время эмоциональных и высших когнитивных процессов: последствия для взаимодействия между эмоциями и познанием. Cogn emotioin 12: 353-385. DOI: 10.1080 / 026999398379646 |
| [42] | Ботвиник М.М., Нистром Л., Фисселл К. и др. (1999) Мониторинг конфликтов в сравнении с выбором действия в передней поясной коре головного мозга. Природа 402: 179-181. DOI: 10.1038 / 46035 |
| [43] | Ботвиник М.М., Бравер Т.С., Барч Д.М. и др.(2001) Мониторинг конфликтов и когнитивный контроль. Psychol Rev 108: 624-652. DOI: 10.1037 / 0033-295X.108.3.624 |
| [44] | Копп Б., Табелинг С., Мошнер С. и др. (2006) Фракционирование нейронных механизмов когнитивного контроля. J Cogn Neurosci : 949-965. |
| [45] | Ван Вин В., Картер С.С. (2002) Сроки процессов мониторинга действий в передней поясной коре головного мозга. Дж. Cogn Neurosci 14: 593-602. DOI: 10.1162 / 089892045837 |
| [46] | Познер М.И., Шиз Б.Е., Одлудаш Й. и др. (2006) Анализ и формирование сетей внимания человека. Нейронные сети 19: 1422-1429. DOI: 10.1016 / j.neunet.2006.08.004 |
| [47] | Dosenbach NUF, Fair Da, Cohen AL, et al.(2008) Двойная сетевая архитектура управления сверху вниз. Trends Cogn Sci 12: 99-105. DOI: 10.1016 / j.tics.2008.01.001 |
| [48] | Руэда MR (2014) Развитие внимания. Oxford Handb Cogn Neurosci 1: 296-318. |
| [49] | Руэда М.Р., Познер М.И., Ротбарт М.К. и др.(2004) Разработка временного курса для обработки конфликта: исследование связанных с событием потенциалов с участием 4-летних и взрослых. BMC Neurosci 5: 39. DOI: 10.1186 / 1471-2202-5-39 |
| [50] | Абундис-Гутьеррес А., Чека П., Кастелланос С. и др. (2014) Электрофизиологические корреляты сетей внимания в детстве и раннем взрослении. Neuropsychologia 57: 78-92. DOI: 10.1016 / j.neuropsychologia.2014.02.013 |
| [51] | Gießing C, Thiel CM, Alexander-Bloch aF, et al. (2013) Изменения функциональной сети человеческого мозга, связанные с повышенным или нарушенным исполнением задач на внимание. J Neurosci 33: 5903-5914. DOI: 10.1523 / JNEUROSCI.4854-12.2013 |
| [52] | Гао В., Жуб Х.Т., Джованелло К.С. и др. (2009) Доказательства появления сети мозга по умолчанию у 2-недельных здоровых педиатрических субъектов в 2-летнем возрасте. Proc Natl Acad Sci U S A 106: 6790-6795. DOI: 10.1073 / pnas.0811221106 |
| [53] | Fair DA, Dosenbach NU, Church JA и др.(2007) Развитие отдельных сетей контроля через сегрегацию и интеграцию. Proc Natl Acad Sci U S A 104: 13507-13512. DOI: 10.1073 / pnas.0705843104 |
| [54] | Ярмарка Д.А., Коэн А.Л., Пауэр Д.Д. и др. (2009) Функциональные сети мозга развиваются от «локальной к распределенной» организации. PLoS Comput Biol 5: e1000381.DOI: 10.1371 / journal.pcbi.1000381 |
| [55] | Fan J, Wu Y, Fossella JA и др. (2001) Оценка наследуемости сетей внимания. BMC Neurosci 2:14. DOI: 10.1186 / 1471-2202-2-14 |
| [56] | Marrocco RT, Davidson MC (1998) Нейрохимия внимания. Внимательный мозг , изд Парасураман Р. (MIT Press, Кембридж, Массачусетс), стр. 35-50. |
| [57] | Congdon E, Lesch KP, Canli T (2008) Анализ полиморфизмов DRD4 и DAT и поведенческого торможения у здоровых взрослых: последствия для импульсивности. Am J Med Genet B Neuropsychiatr Genet 147: 27-32. |
| [58] | Rueda MR, Rothbart MK, McCandliss BD, et al.(2005) Тренировка, созревание и генетическое влияние на развитие исполнительного внимания. Proc Natl Acad Sci U S A 102: 14931-14936. DOI: 10.1073 / pnas.0506897102 |
| [59] | Diamond A (2007) Последствия вариаций генов, влияющих на дофамин в префронтальной коре. Кора головного мозга 17: i161-170.DOI: 10.1093 / cercor / bhm082 |
| [60] | Форбс Э.Е., Браун С.М., Кимак М. и др. (2009) Генетические вариации компонентов нейротрансмиссии дофамина влияют на вентральную полосатую реактивность, связанную с импульсивностью. Mol Psychiatry 14: 60-70. DOI: 10.1038 / sj.mp.4002086 |
| [61] | Конгдон Э., Констебль Р.Т., Леш К.П. и др.(2009) Влияние вариации SLC6A3 и COMT на нервную активацию во время ингибирования ответа. Biol Psychol 81: 144-152. DOI: 10.1016 / j.biopsycho.2009.03.005 |
| [62] | Мюллер Э.М., Макейг С., Стеммлер Г. и др. (2011) Влияние допамина на обработку ошибок человека зависит от генотипа катехол-О-метилтрансферазы VAL158MET. J Neurosci 31: 15818-15825. DOI: 10.1523 / JNEUROSCI.2103-11.2011 |
| [63] | Espeseth T, Sneve MH, Rootwelt H и др. (2010) Ген никотинового рецептора CHRNA4 взаимодействует с процессорной нагрузкой во внимании. PLoS One 5: e14407. DOI: 10.1371 / journal.pone.0014407 |
| [64] | Greenwood PM, Parasuraman R, Espeseth T (2012) Когнитивный фенотип полиморфизма в гене никотинового рецептора CHRNA4.Neurosci Biobeha Rev 36: 1331-1341. DOI: 10.1016 / j.neubiorev.2012.02.010 |
| [65] | Lundwall Ra, Guo DC, Dannemiller JL (2012) Экзогенная визуальная ориентация связана со специфическими генетическими маркерами нейротрансмиттеров: популяционное генетическое исследование ассоциации. PLoS One 7. |
| [66] | Зозулинский П., Гринбаум Л., Бранде-Эйлат Н. и др.(2014) Гены дофаминовой системы связаны с предвзятостью ориентации здоровых людей. Neuropsychologia 62: 48-54. DOI: 10.1016 / j.neuropsychologia.2014.07.005 |
| [67] | Sheese BE, Voelker P, Posner MI, et al. (2009) Генетическая изменчивость влияет на раннее развитие реактивных эмоций и их регуляцию вниманием. Cogn Neuropsychiatry 14: 332-355. DOI: 10.1080 / 135468004064 |
| [68] | Познер М.И., Ротбарт М.К., Шиз Б.Е. (2007) Гены внимания. Dev Sci 10: 24-29. DOI: 10.1111 / j.1467-7687.2007.00559.x |
| [69] | Bornstein MH, Bradley RH (2003) Социально-экономический статус, воспитание и развитие ребенка (Lawrence Erlbaum Associates Publishers, Махва, Нью-Джерси). |
| [70] | Бернье А., Карлсон С. М., Уиппл Н. (2010) От внешнего регулирования к саморегулированию: ранние родительские предшественники исполнительного функционирования маленьких детей. Child Dev 81: 326-339. DOI: 10.1111 / j.1467-8624.2009.01397.x |
| [71] | Гертнер Б.М., Спинрад Т.Л., Айзенберг Н. (2008) Сосредоточенное внимание у малышей: измерение, стабильность и отношения к негативным эмоциям и воспитанию детей. Infant Child Dev 17: 339-363. DOI: 10.1002 / icd.580 |
| [72] | Cipriano EA, Stifter CA (2010) Прогнозирование усердного контроля дошкольников на основе темперамента малыша и поведения родителей. J Appl Dev Psychol 31: 221-230. DOI: 10.1016 / j.appdev.2010.02.004 |
| [73] | Лью Дж., Чен К., Хьюз Дж. Н. (2010) Детский контроль, отношения между учителем и учеником, а также успеваемость у детей из группы риска: аддитивные и интерактивные эффекты. Early Child Res Q 25: 51-64. DOI: 10.1016 / j.ecresq.2009.07.005 |
| [74] | Hackman D, Farah M (2009) Социально-экономический статус и развивающийся мозг Дэниэл. Trends Cogn Sci 13: 65-73. DOI: 10.1016 / j.tics.2008.11.003 |
| [75] | Ванлесс С.Б., Макклелланд М.М., Томини С.Л. и др.(2011) Влияние демографических факторов риска на поведенческое регулирование детей в дошкольном и детском садах. Early Educ Dev 22: 461-488. DOI: 10.1080 / 10409289.2011.536132 |
| [76] | Mezzacappa E (2004) Предупреждающее, ориентирующее и исполнительное внимание: особенности развития и социально-демографические корреляты в эпидемиологической выборке маленьких городских детей. Чайлд Дев 75: 1373-1386. DOI: 10.1111 / j.1467-8624.2004.00746.x |
| [77] | Clearfield MW, Niman LC (2012) SES влияет на когнитивную гибкость младенцев. Infant Behav Dev 35: 29-35. DOI: 10.1016 / j.infbeh.2011.09.007 |
| [78] | Лоусон Дж. М., Дуда Дж. Т., Авантс ВВ и др.(2013) Связь между социально-экономическим статусом детей и толщиной префронтальной коры. Dev Sci 16: 641-652. DOI: 10.1111 / desc.12096 |
| [79] | Jolles DD, Crone EA (2012) Тренировка развивающегося мозга: нейрокогнитивная перспектива. Front Hum Neurosci 6: 76. |
| [80] | Тан И-И, Познер М.И. (2009) Тренировка внимания и тренировка состояния внимания. Trends Cogn Sci 13: 222-227. DOI: 10.1016 / j.tics.2009.01.009 |
| [81] | Карбах Дж., Край Дж. (2009) Насколько полезно обучение руководящего контроля? Возрастные различия в ближнем и дальнем переносе обучения с переключением задач. Dev Sci 12: 978-990. |
| [82] | Джегги С.М., Бушкуль М., Йонидес Дж. И др.(2011) Краткосрочные и долгосрочные преимущества когнитивной тренировки. Proc Natl Acad Sci U S A 108: 10081-10086. DOI: 10.1073 / pnas.1103228108 |
| [83] | Торелл Л. Б., Линдквист С., Бергман Натли С. и др. (2008) Эффекты обучения и передачи управляющих функций у дошкольников. Dev Sci 12: 106-113. |
| [84] | Олесен П.Дж., Вестерберг Х., Клингберг Т. (2004) Повышенная префронтальная и теменная активность после тренировки рабочей памяти. Nat Neurosci 7: 75-79. DOI: 10.1038 / nn1165 |
| [85] | Джоллес Д.Д., Ван Бучем М.А., Крон Е.А. и др.(2013) Функциональная связность мозга в состоянии покоя изменяется после тренировки рабочей памяти. Hum Brain Mapp 34: 396-406. DOI: 10.1002 / hbm.21444 |
| [86] | Макнаб Ф., Андреа В., Ларс Ф. и др. (2009) Изменения в связывании кортикального дофаминового рецептора D1, связанные с когнитивной тренировкой. Наука 323: 800-802. doi: 10.1126 / science.1166102 |
| [87] | Тан И-Й, Познер М.И. (2014) Тренировка сетей и состояний мозга. Trends Cogn Sci 18: 345-350. DOI: 10.1016 / j.tics.2014.04.002 |
| [88] | Малиновский П. (2013) Нейронные механизмы контроля внимания в медитации осознанности. Front Neurosci 7: 8. |
| [89] | Тан И-И, Ма Й.Х., Ван Дж. Х. и др. (2007) Краткосрочные тренировки по медитации улучшают внимание и саморегуляцию. Proc Natl Acad Sci U S A 104: 17152-17156. DOI: 10.1073 / pnas.0707678104 |
| [90] | Мур А., Грубер Т., Дероз Дж. И др.(2012) Регулярная, краткая практика медитации осознанности улучшает электрофизиологические маркеры контроля внимания. Front Hum Neurosci 6: 1-15. |
| [91] | Slagter HA, Lutz A, Greischar LL, et al. (2007) Психологическая тренировка влияет на распределение ограниченных ресурсов мозга. Плос Биол 5. |
| [92] | Hölzel BK, Ott U, Hempel H, et al.(2007) Дифференциальное взаимодействие передней поясной извилины и прилегающей медиальной лобной коры у опытных медитирующих и не медитирующих. Neurosci Lett 421: 16-21. DOI: 10.1016 / j.neulet.2007.04.074 |
| [93] | Тан И-И, Лу Кью, Фан М. и др. (2012) Механизмы изменения белого вещества, вызванные медитацией. Proc Natl Acad Sci : 1-5. |
| [94] | Познер М.И., Тан И-И, Линч Г. (2014) Механизмы изменения белого вещества, вызванные тренировкой по медитации. Front Psychol 5: 1-4. |
| [95] | Тан Y-Y, Lua Ql, Gengc XJ и др.(2010) Кратковременная медитация вызывает изменения белого вещества в передней части поясной извилины. Proc Natl Acad Sci U S A 107: 15649-15652. DOI: 10.1073 / pnas.1011043107 |
| [96] | Хебб Д.О. (1949) Организация поведения (John Wiley & Sons, Нью-Йорк, Нью-Йорк). |
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки вашего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
.