особенности, значение, примеры и механизмы регуляции
Автор Nat WorldВремя чтения 5 мин.Просмотры 1.3k.Опубликовано Обновлено
Гомеостаз — любой саморегулирующийся процесс, с помощью которого биологические системы устремляются к поддержанию внутренней стабильности, приспосабливаясь к оптимальным для выживания условиям. Если гомеостаз успешен, то жизнь продолжается; в противном случае, произойдет бедствие или смерть. Достигнутая стабильность фактически является динамическим равновесием, в котором происходят непрерывные изменения, но преобладают относительно однородные условия.
Особенности и роль гомеостаза
Любая система в динамическом равновесии желает достичь устойчивого состояния, баланса, который противостоит внешним изменениям. Когда такая система нарушена, встроенные регулирующие устройства реагируют на отклонения, чтобы установить новый баланс. Такой процесс является одним из элементов управления с обратной связью. Примерами гомеостатической регуляции являются все процессы интеграции и координации функций, опосредованные электрическими цепями и нервными или гормональными системами.
Такой процесс является одним из элементов управления с обратной связью. Примерами гомеостатической регуляции являются все процессы интеграции и координации функций, опосредованные электрическими цепями и нервными или гормональными системами.
Другим примером гомеостатической регуляции в механической системе является действие регулятора комнатной температуры или термостата. Сердцем термостата является биметаллическая полоса, которая реагирует на изменения температуры, завершая или нарушая электрическую цепь. Когда помещение охлаждается, то контур завершается и включается обогрев, а температура поднимается. На заданном уровне цепь прерывается, печь останавливается, и температура падает.
Однако биологические системы, имеющие большую сложность, обладают регуляторами, которые сложно сравнивать с механическими устройствами.
Как отмечалось ранее, термин гомеостаз относится к поддержанию внутренней среды тела в узких и жестко контролируемых пределах. Основными функциями, важными для поддержания гомеостаза, являются баланс жидкости и электролита, регулирование кислотной среды, терморегуляция и метаболический контроль.
Контроль температура тела у людей считается отличным примером гомеостаза в биологической системе. Нормальная температура тела человека составляет около 37° C, но различные факторы могут влиять на этот показатель, включая гормоны, скорость метаболизма и болезни, приводящие к чрезмерно высоким или низким температурам. Регулирование температуры тела контролируется областью мозга, называемой Гипоталамус.
Обратная связь о температуре тела переносится через кровоток в мозг и приводит к компенсационным корректировкам в скорости дыхания, уровне сахара в крови и скорости метаболизма. Потеря тепла у людей обеспечивается уменьшением активности, потоотделением и механизмами теплообмена, которые позволяют большему количеству крови циркулировать вблизи поверхности кожи.
Снижение потерь тепла осуществляется за счет изоляции, уменьшения циркуляции на коже и культурных изменений, таких как использование одежды, жилья и сторонних источников тепла. Диапазон между высокими и низкими уровнями температуры тела составляет гомеостатическое плато — «нормальный» диапазон, который поддерживает жизнь. По мере приближения к любой из двух крайностей, корректирующее действие (через отрицательную обратную связь) возвращает систему в нормальный диапазон.
По мере приближения к любой из двух крайностей, корректирующее действие (через отрицательную обратную связь) возвращает систему в нормальный диапазон.
Концепция гомеостаза также применяется к экологическим условиям. Впервые предложенная американским экологом Робертом Макартуром в 1955 году идея, что гомеостаз в экосистемах является продуктом сочетания биоразнообразия и большого количества экологических взаимодействий, происходящих между видами.
Такое предположение считалось концепцией, которая могла бы помочь объяснить устойчивость экологической системы, то есть ее сохранение как определенного типа экосистемы с течением времени. С тех пор концепция несколько изменилась, и включила неживую составляющую экосистемы. Этот термин использовался многими экологами для описания взаимности, которая происходит между живыми и неживыми составляющими экосистемы для поддержания статус-кво.
Гипотеза Геи — модель Земли, предложенная английским ученым Джеймсом Лавлоком, которая рассматривает различные живые и неживые составляющие, как компоненты более крупной системы или единого организма, делая предположение, что коллективные усилия отдельных организмов вносят вклад в гомеостаз на планетарном уровне.
Клеточный гомеостаз
Клетки зависят от среды тела, чтобы сохранять жизнеспособность и правильно функционировать. Гомеостаз поддерживает среду тела под контролем и сохраняет благоприятные условия для клеточных процессов. Без правильных условий тела определенные процессы (к примеру, осмос) и белки (к примеру, ферменты) не будут функционировать должным образом.
Почему гомеостаз важен для клеток? Живые клетки зависят от движения химических веществ вокруг них. Химические вещества, такие как кислород, углекислый газ и растворенная пища, необходимо транспортировать в клетки и из них. Это осуществляется процессами диффузии и осмоса, зависящих от баланса воды и соли в теле, которые поддерживаются гомеостазом.
Клетки зависят от ферментов, чтобы ускорить многие химические реакции, поддерживающие жизнедеятельность и функциональность клеток. Эти ферменты работают лучше всего при определенных температурах, и поэтому снова гомеостаз жизненно важен для клеток, поскольку он поддерживает постоянную температуру тела.
Примеры и механизмы гомеостаза
Вот несколько основных примеров гомеостаза в теле человека, а также поддерживающие их механизмы:
Температура тела
Наиболее распространенным примером гомеостаза у людей является регулирование температуры тела. Нормальная температура тела, как мы писали выше составляет 37° C. Температура выше или ниже нормальных показателей может вызывать серьезные осложнения.
Мышечная недостаточность возникает при температуре 28° C. При 33° C происходит потеря сознания. При температуре 42° C центральная нервная система начинает разрушаться. Смерть наступает при температуре 44° C. Тело контролирует температуру путем выработки или высвобождения избыточного тепла.
Концентрация глюкозы
Концентрация глюкозы относится к количеству глюкозы (сахара в крови), присутствующего в кровотоке. Организм использует глюкозу в качестве источника энергии, но ее избыток или недостаток может вызвать серьезные осложнения. Некоторые гормоны осуществляют регулирования концентрации глюкозы в крови. Инсулин снижает концентрацию глюкозы, в то время как кортизол, глюкагон и катехоламины увеличивают.
Инсулин снижает концентрацию глюкозы, в то время как кортизол, глюкагон и катехоламины увеличивают.
Уровни кальция
Кости и зубы содержат приблизительно 99% кальция в организме, в то время как оставшийся 1% циркулируют в крови. Слишком большое или недостаточное содержание кальция в крови имеют негативные последствия. Если уровень кальция в крови слишком сильно снижается, паращитовидные железы активируют свои рецепторы, чувствительные к кальцию, и высвобождают паратиреоидный гормон.
ПТГ сигнализирует костям он необходимости высвобождения кальция, чтобы увеличить его концентрацию в кровотоке. Если уровень кальция увеличивается слишком сильно, щитовидная железа высвобождает кальцитонин и фиксирует избыток кальция в костях, тем самым уменьшая количество кальция в крови.
Объем жидкости
Тело должно поддерживать постоянную внутреннюю среду, а это означает, что ему необходимо регулировать потерю или восполнение жидкости. Гормоны помогают регулировать этот баланс, вызывая экскрецию или удерживание жидкости. Если организму не хватает жидкости, антидиуретический гормон сигнализирует почкам о сохранении жидкости и уменьшает выход мочи. Если организм содержит слишком много жидкости, он подавляет альдостерон и сигнализирует о выделении большего количества мочи.
Если организму не хватает жидкости, антидиуретический гормон сигнализирует почкам о сохранении жидкости и уменьшает выход мочи. Если организм содержит слишком много жидкости, он подавляет альдостерон и сигнализирует о выделении большего количества мочи.
Не все нашли? Используйте поиск по сайту ↓
ГОМЕОСТАЗ • Большая российская энциклопедия
ГОМЕОСТА́З, гомеостазис (от гомео… и греч. στᾴσις – неподвижность, состояние).
В биологии Г. – способность живых организмов сохранять динамич. постоянство состава и свойств внутр. среды. Идея о наличии в организме комплекса физиологич. механизмов, направленных на поддержание постоянства внутр. среды, была высказана К. Бернаром во 2-й пол. 19 в. Он считал, что в основе свободной и независимой жизни организмов в постоянно меняющейся внешней среде лежит постоянство физико-химич. условий внутр. среды. Для обозначения комплекса процессов в живом организме, обеспечивающих постоянство таких условий, У. Кеннон предложил термин «Г.» (1929).
Кеннон предложил термин «Г.» (1929).
Появление на Земле одноклеточных организмов было связано с формированием и поддержанием внутри клетки в течение всей её жизни специфич. физико-химич. условий, отличных от условий окружающей среды. Дальнейшая эволюция жизни сопровождалась возникновением многоклеточных животных, дифференцировкой их клеток, формированием внутр. среды, в которой находятся и взаимодействуют между собой клетки. Появляются система внеклеточных жидкостей, лимфа, кровь, из которых клетки извлекают необходимые органич. и неорганич. вещества, O
У морских беспозвоночных Г. касается объёма жидкостей внутр. среды, концентрации в ней отд. ионов, рН. Адаптация организмов к пресным водам потребовала формирования новой системы регуляции – поддержания на постоянном уровне осмотич. давления жидкостей внутр. среды, удаления из организма избытка воды. К особо контролируемым физико-химич. параметрам внутр. среды относятся её осмотич. давление (изоосмия), концентрация отд. ионов (изоиония), объём крови (изоволемия), её рН, у птиц и млекопитающих также стабилизированная температура тела (изотермия) и др.
Постоянство физико-химич. условий во внутр. среде, состояние околоклеточной среды служат жизненно важным фактором, необходимым для эффективной работы клеток; их адекватная реакция на сигналы из внешней (напр., световые, звуковые, температурные раздражители) и внутренней (в т. ч. импульсы нервной системы, гормоны, аутакоиды) среды возможна при поддержании Г. Особенно высокая степень Г. характерна для млекопитающих, у которых наиболее строго поддерживаются осмоляльность крови, концентрация в ней ионов Са2+, рН, изотермия.
Г. создаёт возможность для адекватных реакций клеток, поддержания необходимого уровня их метаболизма и ответа на внешние воздействия. В регуляции физико-химич. параметров внутр. среды участвуют нервная и эндокринная системы, аутакоиды. Повышение качества регуляции для сохранения стабильности параметров внутр.
Термин «клеточный Г.» противоречит смысловому значению понятия, предложенного К. Бернаром и У. Кенноном.
Гомеостаз в кибернетике. В 1950-х гг. Н. Винер универсализировал понятие Г. и применил его к функционированию достаточно сложных саморегулирующихся систем. В результате понятие Г. стало широко использоваться не только в биологии, но и в др. науках. По Винеру, гомеостатич. алгоритм определяет базовые параметры системы, значит. изменения которых нарушают или разрушают её нормальное функционирование и развитие; фиксирует пределы допустимого изменения установленных параметров под влиянием как внешней, так и внутр. среды; выявляет совокупность механизмов, начинающих проявлять себя при критич. изменении базовых параметров системы. Гомеостатич.
В социальных и политических науках понятие Г.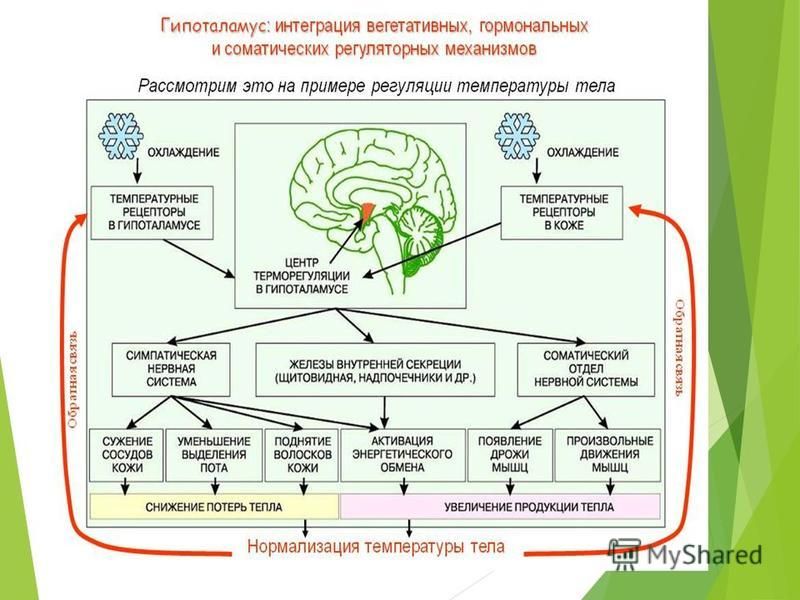 применяется преим. при анализе функционирования и динамики социальной и политич. систем, а также некоторых системных организаций (государства, партий, профсоюзов и др.). В этой сфере подвижное равновесное состояние систем (и подсистем) сохраняется через противодействие их структур, социальных групп и институтов внешним и внутр. факторам, нарушающим осн. принципы функционирования (Т. Парсонс, Д. Истон). В политич. анализе и управлении используются социодинамич. модели политич. и социальной систем общества, в которых выделяются прямые и обратные гомеостатич. взаимодействия системы с внешней средой. В социально-политич. системах велика роль человеческого фактора (риск ошибок и др.) при принятии решений, поскольку гиперактивное воздействие на среду и её изменение формируются самими социально-политич. агентами. Соответственно в этой сфере велика функциональная роль обществ.
применяется преим. при анализе функционирования и динамики социальной и политич. систем, а также некоторых системных организаций (государства, партий, профсоюзов и др.). В этой сфере подвижное равновесное состояние систем (и подсистем) сохраняется через противодействие их структур, социальных групп и институтов внешним и внутр. факторам, нарушающим осн. принципы функционирования (Т. Парсонс, Д. Истон). В политич. анализе и управлении используются социодинамич. модели политич. и социальной систем общества, в которых выделяются прямые и обратные гомеостатич. взаимодействия системы с внешней средой. В социально-политич. системах велика роль человеческого фактора (риск ошибок и др.) при принятии решений, поскольку гиперактивное воздействие на среду и её изменение формируются самими социально-политич. агентами. Соответственно в этой сфере велика функциональная роль обществ.
В макроэкономике гомеостатич. подход лёг в основу теории общего экономич. равновесия (кейнсианская и неоклассическая модели). Принцип Г. применяется в социальной экологии, которая рассматривает природную среду как дифференцированную систему динамич.
Механизмы гомеостаза | Кинезиолог
Сазонов В.Ф. Механизмы гомеостаза
Гомеостаз в биологии – это поддержание постоянства внутренней среды организма.
В основе гомеостаза лежит чувствительность организма к отклонению определённых параметров (гомеостатических констант) от заданного значения. Пределы допустимых колебаний гомеостатического параметра (гомеостатической константы) могут быть широкими и узкими. Узкие пределы имеют: температура тела, рН крови, содержание глюкозы в крови. Широкие пределы имеют: давление крови, масса тела, концентрация аминокислот в крови.
Специальные внутриорганизменные рецепторы (интерорецепторы) реагируют на отклонение гомеостатических параметров от заданных пределов. Такие интерорецепторы имеются внутри таламуса, гипоталамуса, в сосудах и в органах. В ответ на отклонение параметров они запускают восстановительные гомеостатические реакции.
В ответ на отклонение параметров они запускают восстановительные гомеостатические реакции.
Регуляция по отклонению – основной механизм гомеостаза, при котором сверхпороговое изменение оптимального уровня регулируемого показателя инициирует деятельность соответствующих систем, приводящую к восстановлению его значения до исходного уровня.
Общий механизм нейроэндокринных гомеостатических реакций для внутренней регуляции гомеостаза
Параметры гомеостатической константы отклоняются, интерорецепторы возбуждаются, затем возбуждаются соответствующие центры гипоталамуса, они стимулируют выброс гипоталамусом соответствующих либеринов. В ответ на действие либеринов происходит выброс гормонов гипофизом, а затем под их действием идёт выброс гормонов других эндокринных желёз. Гормоны, выделившись из желёз внутренней секреции в кровь, изменяют обмен веществ и режим работы органов и тканей. В итоге установившийся новый режим работы органов и тканей смещает изменившиеся параметры в сторону прежнего заданного значения и восстанавливает величину гомеостатической константы. Таков общий принцип восстановления гомеостатических констант при их отклонении.
Таков общий принцип восстановления гомеостатических констант при их отклонении.
Примеры
Мочеобразование и выведение мочи. Дыхание: чувствительность к избытку СО2 заставляет дышать чаще и восстанавливать тем самым стандартную концентрацию СО2. Теплообмен.
Механизм гомеостаза первого порядка
Гомеостаз поддерживается механизмами нескольких уровней, как это обычно свойственно иерархическим системам. При отклонении избранного параметра от средней линии в сторону верхнего или нижнего предела сразу же включаются «ближайшие» компенсационные механизмы, которые гасят это отклонение. Собственно это и будет называться регуляцией гомеостаза как устойчивого состояния, а поскольку процессы автоматизированы за счёт отрицательных обратных связей, то данное явление можно назвать саморегуляцией.
Итак, колебание гомеостатической константы допустимо в определённых пределах. За счет автоматического гашения отклонений гомеостатический параметр настойчиво возвращается к средней линии. В идеале данный механизм стремится миминизировать колебания гомеостатического параметра вокруг средней линии. Чем лучше работает этот механизм, тем меньше будут колебания. Можно назвать это первым гомеостатическим механизмом, он является базовым. Например, именно так работают различные буферные системы, компенсирующие небольшие отклонения, в частности отклонения в рН среды.
В идеале данный механизм стремится миминизировать колебания гомеостатического параметра вокруг средней линии. Чем лучше работает этот механизм, тем меньше будут колебания. Можно назвать это первым гомеостатическим механизмом, он является базовым. Например, именно так работают различные буферные системы, компенсирующие небольшие отклонения, в частности отклонения в рН среды.
Механизм гомеостаза второго порядка
Это регуляция гомеостаза второго порядка, которая накладывается на первый механизм поддержания гомеостаза. При выходе гомеостатического параметра за определённый верхний или нижний предел допустимых колебаний включается гомеостатический механизм второго порядка и возвращает параметр в заданные пределы. Если происходят какие-либо более мощные изменения и достигаются более амплитудные пределы, то подключаются механизмы гомеостаза следующего уровня и так далее.
Например, при недостатке кислорода в крови (гипоксии) в качестве первого механизма гомеостаза будет запускаться усиление частоты и глубины дыхания. Но при длительном недостатке кислорода в крови, который не устраняется учащённым дыханием, запускается второй механизм гомеостаза: увеличение количества эритроцитов в крови. Для этого будет активироваться процесс выработки эритроцитов (эритропоэз). В результате количество эритроцитов в крови возрастёт, и соответственно повысится концентрация кислорода в крови. Таким образом, данная гомеостатическая константа начнёт возвращаться к норме.
Источник изображения: Зинчук, В В. Нормальная физиология. Краткий курс : учеб. пособие / В. В. Зинчук, О.А. Балбатун, Ю.М. Емельянчик; под ред. В.В. Зинчука. – 3-е изд., стер. – Минск : Вышэйшая школа, 2014. – 431 с.
Механизм гомеостаза третьего порядка
При длительном или постоянном смещении гомеостатического параметра от средней линии к одному из пределов (верхнему или нижнему) включаются компенсационные процессы по гашению процесса смещения, предназначенные для возврате параметра к средней линии. Система этого вида гомеостаза чувствительна к длительному общему смещению средней линии вверх или вниз. Механизмы компенсации должны быть другими по сравнению с гомеостазом первого порядка.
В качестве примера можно представить себе нормально работающий холодильник, в котором вдруг начали непрерывно открывать и закрывать дверцу. Обычных механизмов поддержания холода при этом становится уже недостаточно.
Признаки холодильника с хорошим гомеостазом:
1. Не допускает значительных отклонений от средней линии.
2. Быстро компенсирует появившиеся отклонения
3. Имеет способность к самообучению – в процессе адаптации размах колебания уменьшается.
В отличие от искусственных автоматических систем гомеостатические живые системы, обладают уникальным свойством пластичности. Они перестраивают свою деятельность в результате постоянной нагрузки определённого рода. Самый впечатляющий пример биологической пластичности — это смена знака при управлении обратной связью с «-» на «+» и наоборот.
Гомеостатические реакции, противодействующие отклонению гомеостатической константы
1. Обратимые кратковременные функциональные изменения.
Пример:
При повышении температуры окружающей среды повышается температура тела. В ответ на это начинается кратковременное усиление потоотделения. За счёт этого усиливается испарение и, как следствие — происходит охлаждение организма. После нормализации температуры тела потоотделение возвращается к норме. Таким образом, это было обратимое кратковременное функциональное изменение.
2. Обратимые долговременные функциональные изменения.
Пример:
При воздействии…
3. Необратимые долговременные функциональные изменения.
4. Обратимые кратковременные структурные изменения.
5. Обратимые долговременные структурные изменения.
Пример:
При воздействии на организм интенсивного солнечного света возникают повреждения глубоких слоёв кожи. В ответ на это начинается усиление выработки пигмента меланина клетками кожи. В результате появляется загар, который препятствует глубокому проникновению солнечных лучей под кожу. Процесс повреждения клеток солнечным светом прекращается. После прекращения действия солнечного света через длительный срок пигмент исчезает, и кожные покровы возвращаются к первоначальному состоянию. Таким образом, это был пример обратимого долговременного структурного изменения.
6. Необратимые долговременные структурные изменения.
7. Кратковременные поведенческие реакции.
8. Долговременные поведенческие изменения.
9. Наследственные изменения.
10. Изменения образа жизни.
11. Изменения популяции.
12. Изменение биологического вида.
Переключение регуляции гомеостаза с внутренних нейрогуморальных механизмов на внешние поведенческие
Для понимания того, как просиходит переключение регуляции гомеостаза с внутренних механизмов на внешние, необходимо рассмотреть представления о потребности и мотивации с точки зрения психофизиологии.
1. Сенсорные интерорецепторы, реагирующие на изменение гомеостатических констант организма, через афферентные нейроны передают возникающее в них сенсорное возбуждение в соответствующие функциональные нервные центры продолговатого мозга, среднего мозга и гипоталамуса, которые можно назвать центрами биорегуляции.
2. В этих функциональных нервных центрах определяется отклонение данных констант от нормы. Отклонение констант в заданных пределах устраняется за счёт регуляторных возможностей самих функциональных центров.
3. Однако при отклонении любой гомеостатической константы выше или ниже допустимых пределов функциональные центры передают возбуждение выше: в «потребностные центры» гипоталамуса. Это необходимо для того, чтобы переключиться с внутренней нейрогуморальной регуляции гомеостаза на внешнюю — поведенческую.
4. Возбуждение того или иного потребностного центра гипоталамуса формирует соответствующее ему функциональное состояние, которое субъективно переживается как потребность в чём-то: пище, воде, тепле, холоде или сексе. Возникает активирующее и побуждающее к действию психоэмоциональное состояние неудовлетворённости.
5. Для организации целенаправленного поведения необходимо выбрать только одну из потребностей в качестве первоочередной и создать для её удовлетворения рабочую доминанту. Считается, что главную роль в этом играют миндалины мозга (Сorpus amygdoloideum). Получается, что на основе одной из потребностей, которые формирует гипоталамус, миндалина создаёт ведущую мотивацию, организующую целенаправленное поведение для удовлетворения только одной этой избранной потребности.
6. Следующим этапом можно считать запуск подготовительного поведения, или драйв-рефлекса, который должен повысить вероятность для запуска исполнительного рефлекса в ответ на пусковой стимул. Драйв-рефлекс побуждает организм к созданию такой ситуации, в которой будет повышена вероятность обнаружения объекта, подходящего для удовлетворения текущей потребности. Это может быть, например, перемещение в место, богатое пищей, или водой, или сексульными партнёрами, в зависимости от ведущей потребности. Когда же в достигнутой ситуации обнаруживается конкретный объект, подходящий для удовлетворения данной доминантной потребности, то он запускает исполнительное рефлекторное поведение, направленное на удовлетворение потребности с помощью именно этого объекта.
© 2014-2020 Сазонов В.Ф. © 2014-2020 kineziolog.bodhy.ru. © 2016-2020 kineziolog.su.
Системы гомеостаза — подробный образовательный ресурс по гомеостазу.
Понятие гомеостаза | Кинезиолог
Сазонов В.Ф. Понятие гомеостаза
Введение
Одним из важнейших свойств живых организмов является постоянство их внутренней среды, которая может существенно отличаться по своему составу от внешней среды, окружающей организм. Например, рыбы, живущие в солёной морской воде, имеют менее солёную внутреннюю среду, и наоборот, рыбы, живущие в пресной воде, сохраняют свою внутреннюю среду более солёной по сравнению с окружающей их водой. И такое отличие состава своей внутренней среды от внешней организм поддерживает постоянно.
Представление о постоянстве внутренней среды организма было введено в физиологию и медицину французским физиологом Клодом Бернаром. В 1878 г. он сформулировал гипотезу об относительном постоянстве внутренней среды живых организмов. И только в 1929 (1932) г. американский физиолог Уолтер Кэннон предложил для обозначения постоянства внутренней среды организма термин “гомеостаз”. Он показал, что способность организма к поддержанию гомеостаза обеспечивается специальными системами регуляции, которые можно рассматривать по-отдельности как связанные между собой «системы гомеостаза».
Гипоталамус (нижний отдел промежуточного мозга) — главная нервная структура, поддерживающая гомеостаз организма. Для этого у него существует своя рецептивная система и нервные центры гомеостатической регуляции.
Определение понятия
Гомеостаз в широком значении — это поддержание постоянного уравновешенного состояния открытой динамической системы вопреки возмущающим воздействиям на неё.
способности различных открытых систем сохранять постоянство своего внутреннего состояния при наличии возмущающих воздействийИсточник: https://vikent.ru/enc/595/
способности различных открытых систем сохранять постоянство своего внутреннего состояния при наличии возмущающих воздействийИсточник: https://vikent.ru/enc/595/
Гомеостаз в биологическом значении — это поддержание постоянства внутренней среды организма.
Гомеостаз в общем смысле — это сохранение динамического равновесия системы в борьбе с отклонениями её значимых констант от заданных параметров. © 2013-2020 Сазонов В.Ф. © 2016-2020 kineziolog.su.
Гомеокинез — это совокупность процессов, обеспечивающих гомеостаз.
Смысл гомеокинеза — поддержание гомеостатических констант в заданных пределах.
Способ поддержания гомеостатических констант в заданных пределах — противодействие отклонениям гомеостатических констант от физиологической нормы.
Примеры гомеостатического контроля заданных параметров (гомеостатических констант):
- на уровне организма — артериальное давление (АД), базальная температура тела, объём циркулирующей крови и множество других параметров;
- на уровне межклеточного пространства (на примере плазмы крови) — содержание кислорода, углекислоты, глюкозы, K+, Na+, Ca2+, Н+ и множество других;
- на уровне клеток — объём клеток и их органоидов, концентрация ионов (например, K+, Na+ и Ca2+), а также макроэргических соединений (например, АТФ).
«Болевой гомеостаз» — это равновесное состояние между болевыми и антиболевыми сигналами, как нервными, так и химическими (веществом Р и эндорфинами). В какую бы сторону ни сдвинулось это равновесие, сразу же появляется сила, действующая в противоположном направлении (Теппермен Дж., Теппермен Х. Физиология обмена веществ и эндокринной системы.Вводный курс: Пер. с англ. — М.: Мир, 1989. — 656 с.). Болезненная ломка наркоманов ярко иллюстрирует это положение о болевом гомеостазе.
Изменяемая проницаемость мембран почечных канальцев контролирует верхнюю границу концентрации для многих веществ, растворённых в крови. Например, при искусственном повышении их концентрации в организме почки усиливают их выведение из крови в мочу за счёт повышения проницаемости к ним канальцевых мембран. В целом мембранная проницаемость в почках обеспечивает поддержание постоянствп верхней границы концентрационных констант многих веществ крови.
Эндокринные железы контролируют величину гомеостатических констант и обеспечивают её поддержание в заданных пределах. В частности, хорошо известна система гормональной регуляции концентрации глюкозы в крови с помощью инсулина и глюкагона, секретируемых поджелудочной железой в ответ на изменение «глюкозной гомеостатической константы».
Изменения гомеостатических констант
Значения гомеостатических констант не являются абсолютной нормой. Они могут изменяться под воздействием условий окружающей среды и выходить на уровень нового «коридора нормы».
Так, например, считается, что количество сахара (глюкозы) в крови может колебаться в пределах 80-100 мг/дл (это норма), а низкое содержание сахара в крови (менее 60-70 мг/дл) — это уже гипогликемия.
Концентрация сахара (глюкозы) в крови — это системообразующий фактор углеводного гомеостаза. Его концентрация должна поддерживаться на постоянном уровне в довольно узких пределах в качестве «гомеостатической константы». Значительное снижение его содержания в крови приводит к гипогликемической коме — потере сознания из-за недостаточного снабжения клеток мозга глюкозой.
Но вот массовое обследование пришлого населения в условиях Азиатского Севера показало, что содержание сахара (глюкозы) в крови у этих людей находится на нижней границе нормы. У отдельных лиц, постоянно сталкивающихся с экстремальными факторами высоких широт (например, работающих на открытом воздухе в зимних условиях), количество сахара в крови может снижаться до 45-50 мг/дл. Кроме низкого содержания сахара, никаких признаков гипогликемии нет, организм здоров и работоспособен. Такие случаи наблюдались в Антарктиде.
Анализ причин этого явления показал, что в условиях хронического напряжения снижается почечный барьер для сахара, т.е. тот уровень его в крови, при превышении которого сахар начинает появляться в моче. Также в крови оказалось несколько повышено содержание молочной и в меньшей степени пировиноградной кислоты. Выявлены изменения со стороны ключевых ферментов углеводного обмена.
В таких случаях можно сказать, что данная детерминантная система перешла на новый уровень гомеостаза.
Содержание липидных фракций в крови в условиях Заполярья оказалось выше, чем в средних широтах. Повышенным оказалось содержание общих липидов, свободных жирных кислот, липопротеидов различных классов и др. На Севере жиры начинают использоваться в большей степени, а углеводы — в меньшей степени как энергетический материал.
Можно сделать важный вывод о том, что углеводный гомеостаз работает как соподчиненная система белее высокого уровня системной организации — энергетического гомеостаза.
Несмотря на повышенное содержание липидов в крови, при этом не создается условий, благоприятствующих развитию атеросклероза.
Это связано с тем, что у пришлого населения Заполярья в большей степени увеличено содержание ЛПВП (неатерогенной фракции липидов), чем ЛПНП и ЛПОНП (атерогенной фракции). Выше активность липопротеиновой липазы — фермента, превращающего ЛПОНП в ЛПВП, т.е. в неопасные фракции липидов.
Итак, обмен углеводов и липидов в организме следует рассматривать как единое целое, точнее как взаимосвязанные составные части энергетического обмена.
На Крайнем Севере выше артериовенозная разность для свободных жирных кислот (суммарной фракции ЛПНП и ЛПОНП) и ниже для сахара, чем в условиях средних широт. Все это дает основание считать, что при длительном напряжении организма реализуется новая программа действия: переключение энергетического обмена с углеводного типа на липидный.
Отмечено также снижение содержания в крови водорастворимых витаминов, таких как В1, В2 и С. Для них, как и для сахара крови, почечный барьер также ниже, чем в средних широтах. Дополнительное введение витаминов в виде различных фармакологических препаратов (витаминных драже, напитков и др.) не приводит к стойкому повышению содержания их в крови, т.е. выше уровня почечного барьера. Такой гиповитаминоз на Севере не патология, а местная норма. Мы можем считать его местной гомеостатической константой. Несмотря на пониженное содержание водорастворимых витаминов в крови, активность ферментов, в которые эти витамины входят как кофакторы, такая же, как в средних широтах.
В целом отмеченные изменения гомеостатических констант носят адаптационный характер и целесообразны для конкретного климата, для конкретной местности.
Источник: Панин Л. Е. Системные представления о гомеостазе // Бюллетень СО РАМН. 2007. №5. URL: https://cyberleninka.ru/article/n/sistemnye-predstavleniya-o-gomeostaze (дата обращения: 12.11.2018).
Механизмы поддержания гомеостаза
Системы гомеостаза — подробный образовательный ресурс по гомеостазу.
Гомеостаз — это… Что такое Гомеостаз?
Гомеоста́з (др.-греч. ὁμοιοστάσις от ὁμοιος — одинаковый, подобный и στάσις — стояние, неподвижность) — саморегуляция, способность открытой системы сохранять постоянство своего внутреннего состояния посредством скоординированных реакций, направленных на поддержание динамического равновесия. Стремление системы воспроизводить себя, восстанавливать утраченное равновесие, преодолевать сопротивление внешней среды.
Гомеостаз популяции — способность популяции поддерживать определённую численность своих особей длительное время.
Американский физиолог Уолтер Кеннон (Walter B. Cannon) в 1932 году в своей книге «The Wisdom of the Body» («Мудрость тела») предложил этот термин как название для «координированных физиологических процессов, которые поддерживают большинство устойчивых состояний организма». В дальнейшем этот термин распространился на способность динамически сохранять постоянство своего внутреннего состояния любой открытой системы. Однако представление о постоянстве внутренней среды было сформулировано ещё в 1878 году французским учёным Клодом Бернаром.
Общие сведения
Термин «гомеостаз» чаще всего применяется в биологии. Многоклеточным организмам для существования необходимо сохранять постоянство внутренней среды. Многие экологи убеждены, что этот принцип применим также и к внешней среде. Если система неспособна восстановить свой баланс, она может в итоге перестать функционировать.
Комплексные системы — например, организм человека — должны обладать гомеостазом, чтобы сохранять стабильность и существовать. Эти системы не только должны стремиться выжить, им также приходится адаптироваться к изменениям среды и развиваться.
Свойства гомеостаза
Гомеостатические системы обладают следующими свойствами:
- Нестабильность системы: тестирует, каким образом ей лучше приспособиться.
- Стремление к равновесию: вся внутренняя, структурная и функциональная организация систем способствует сохранению баланса.
- Непредсказуемость: результирующий эффект от определённого действия зачастую может отличаться от того, который ожидался.
Примеры гомеостаза у млекопитающих:
Важно отметить, что, хотя организм находится в равновесии, его физиологическое состояние может быть динамическим. Во многих организмах наблюдаются эндогенные изменения в форме циркадного, ультрадианного и инфрадианного ритмов. Так, даже находясь в гомеостазе, температура тела, кровяное давление, частота сердечных сокращений и большинство метаболических индикаторов не всегда находятся на постоянном уровне, но изменяются в течение времени.
Механизмы гомеостаза: обратная связь
Когда происходит изменение в переменных, наблюдаются два основных типа обратной связи, на которые реагирует система:
- Отрицательная обратная связь, выражающаяся в реакции, при которой система отвечает так, чтобы изменить направление изменения на противоположное. Так как обратная связь служит сохранению постоянства системы, это позволяет соблюдать гомеостаз.
- Например, когда концентрация углекислого газа в организме человека увеличивается, лёгким приходит сигнал к увеличению их активности и выдыханию большего количество углекислого газа.
- Терморегуляция — другой пример отрицательной обратной связи. Когда температура тела повышается (или понижается) терморецепторы в коже и гипоталамусе регистрируют изменение, вызывая сигнал из мозга. Данный сигнал, в свою очередь, вызывает ответ — понижение температуры (или повышение).
- Положительная обратная связь, которая выражается в усилении изменения переменной. Она оказывает дестабилизирующий эффект, поэтому не приводит к гомеостазу. Положительная обратная связь реже встречается в естественных системах, но также имеет своё применение.
- Например, в нервах пороговый электрический потенциал вызывает генерацию намного большего потенциала действия. Свёртывание крови и события при рождении можно привести в качестве других примеров положительной обратной связи.
Устойчивым системам необходимы комбинации из обоих типов обратной связи. Тогда как отрицательная обратная связь позволяет вернуться к гомеостатическому состоянию, положительная обратная связь используется для перехода к совершенно новому (и, вполне может быть, менее желанному) состоянию гомеостаза, — такая ситуация называется «метастабильность». Такие катастрофические изменения могут происходить, например, с увеличением питательных веществ в реках с прозрачной водой, что приводит к гомеостатическому состоянию высокой эвтрофикации (зарастание русла водорослями) и замутнению.
Экологический гомеостаз
Экологический гомеостаз наблюдается в климаксовых сообществах с максимально возможным биоразнообразием при благоприятных условиях среды.
В нарушенных экосистемах, или субклимаксовых биологических сообществах — как, например, остров Кракатау, после сильного извержения вулкана в 1883 — состояние гомеостаза предыдущей лесной климаксовой экосистемы было уничтожено, как и вся жизнь на этом острове. Кракатау за годы после извержения прошёл цепь экологических изменений, в которых новые виды растений и животных сменяли друг друга, что привело к биологической вариативности и в результате климаксовому сообществу. Экологическая сукцессия на Кракатау осуществилась за несколько этапов. Полная цепь сукцессий, приведшая к климаксу, называется присерией. В примере с Кракатау на этом острове образовалось климаксовое сообщество с восемью тысячами различных видов, зарегистрированных в 1983, спустя сто лет с того времени, как извержение уничтожило на нём жизнь. Данные подтверждают, что положение сохраняется в гомеостазе в течение некоторого времени, при этом появление новых видов очень быстро приводит к быстрому исчезновению старых.
Случай с Кракатау и другими нарушенными или нетронутыми экосистемами показывает, что первоначальная колонизация пионерными видами осуществляется через стратегии воспроизведения, основанные на положительной обратной связи, при которых виды расселяются, производя на свет как можно больше потомства, но при этом практически не вкладываясь в успех каждого отдельного. В таких видах наблюдается стремительное развитие и столь же стремительный крах (например, через эпидемию). Когда экосистема приближается к климаксу, такие виды заменяются более сложными климаксовыми видами, которые через отрицательную обратную связь адаптируются к специфическим условиям окружающей их среды. Эти виды тщательно контролируются потенциальной ёмкостью экосистемы и следуют иной стратегии — произведению на свет меньшего потомства, в репродуктивный успех которого в условиях микросреды его специфической экологической ниши вкладывается больше энергии.
Развитие начинается с пионер-сообщества и заканчивается на климаксовом сообществе. Это климаксовое сообщество образуется, когда флора и фауна пришла в баланс с местной средой.
Подобные экосистемы формируют гетерархии, в которых гомеостаз на одном уровне способствует гомеостатическим процессам на другом комплексном уровне. К примеру, потеря листьев у зрелого тропического дерева даёт место для новой поросли и обогащает почву. В равной степени тропическое дерево уменьшает доступ света на низшие уровни и помогает предотвратить инвазию других видов. Но и деревья падают на землю и развитие леса зависит от постоянной смены деревьев, круговорота питательных веществ, осуществляемого бактериями, насекомыми, грибами. Схожим образом такие леса способствуют экологическим процессам — таким, как регуляция микроклиматов или гидрологических циклов экосистемы, а несколько разных экосистем могут взаимодействовать для поддержания гомеостаза речного дренажа в рамках биологического региона. Вариативность биорегионов так же играет роль в гомеостатической стабильности биологического региона, или биома.
Биологический гомеостаз
Гомеостаз выступает в роли фундаментальной характеристики живых организмов и понимается как поддержание внутренней среды в допустимых пределах.
Внутренняя среда организма включает в себя организменные жидкости — плазму крови, лимфу, межклеточное вещество и цереброспинальную жидкость. Сохранение стабильности этих жидкостей жизненно важно для организмов, тогда как её отсутствие приводит к повреждению генетического материала.
В отношении любого параметра организмы делятся на конформационные и регуляторные. Регуляторные организмы сохраняют параметр на постоянном уровне, независимо от того, что происходит в среде. Конформационные организмы позволяют окружающей среде определять параметр. Например, теплокровные животные сохраняют постоянную температуру тела, тогда как холоднокровные демонстрируют широкий диапазон температур.
Речь не идёт о том, что конформационные организмы не обладают поведенческими приспособлениями, позволяющими им в некоторой степени регулировать взятый параметр. Рептилии, к примеру, часто сидят на нагретых камнях утром, чтобы повысить температуру тела.
Преимущество гомеостатической регуляции состоит в том, что она позволяет организму функционировать более эффективно. Например, холоднокровные животные, как правило, становятся вялыми при низких температурах, тогда как теплокровные почти так же активны, как и всегда. С другой стороны, регуляция требует энергии. Причина, почему некоторые змеи могут есть только раз в неделю, состоит в том, что они тратят намного меньше энергии для поддержания гомеостаза, чем млекопитающие.
Клеточный гомеостаз
Регуляция химической деятельности клетки достигается с помощью ряда процессов, среди которых особое значение имеет изменение структуры самой цитоплазмы, а также структуры и активности ферментов. Авторегуляция зависит от температуры, степени кислотности, концентрации субстрата, присутствия некоторых макро- и микроэлементов.
Гомеостаз в организме человека
Разные факторы влияют на способность жидкостей организма поддерживать жизнь. В их числе такие параметры, как температура, солёность, кислотность и концентрация питательных веществ — глюкозы, различных ионов, кислорода, и отходов — углекислого газа и мочи. Так как эти параметры влияют на химические реакции, которые сохраняют организм живым, существуют встроенные физиологические механизмы для поддержания их на необходимом уровне.
Гомеостаз нельзя считать причиной процессов этих бессознательных адаптаций. Его следует воспринимать как общую характеристику многих нормальных процессов, действующих совместно, а не как их первопричину. Более того, существует множество биологических явлений, которые не подходят под эту модель — например, анаболизм.
Другие сферы
Понятие «гомеостаз» используется также и в других сферах.
Актуарий может говорить о рисковом гомеостазе, при котором, к примеру, люди, у которых на машине установлены незаклинивающие тормоза, не находятся в более безопасном положении по сравнению с теми, у кого они не установлены, потому что эти люди бессознательно компенсируют более безопасный автомобиль рискованной ездой. Это происходит потому, что некоторые удерживающие механизмы — например, страх — перестают действовать.
Социологи и психологи могут говорить о стрессовом гомеостазе — стремлении популяции или индивида оставаться на определённом стрессовом уровне, зачастую искусственно вызывая стресс, если «естественного» уровня стресса недостаточно.
Примеры
Многие из этих органов контролируются гормонами гипоталамо-гипофизарной системы.
См. также
Роль микробиоты кишечника в поддержании метаболического гомеостаза uMEDp
Микробиота кишечника – совокупность микроорганизмов, находящихся в тесной функциональной связи с организмом. Интеграция метаболизма микробиоты в обмен веществ организма-хозяина достигается посредством регуляции синтеза биологически активных субстанций, таких как свободные жирные кислоты, желчные кислоты, эндоканнабиоидов, модуляции проницаемости кишечной стенки и вовлечения центральных механизмов контроля аппетита.Пробиотические препараты воздействуют на микробиоту организма-хозяина, корректируя дисбиоз и улучшая метаболические показатели, что свидетельствует о потенциальной возможности их использования для профилактики и лечения ожирения, а также ассоциированных с ним заболеваний, наряду с выполнением рекомендаций по диете, модификацией образа жизни и базовой терапией.
Вклад микробиоты в регуляцию энергетического обмена
Роль штаммов, входящих в состав пробиотиков, в коррекции метаболического синдрома
Микробиота кишечника человека представляет собой сложную систему микроорганизмов c колоссальным видовым разнообразием и внушительными количественными характеристиками [1]. В кишечнике насчитывается более 500–1000 видов микроорганизмов. Они составляют 1–3% общей массы организма и являются огромнейшим резервуаром генетического материала [2, 3].
Значимую долю микробиоты составляют, в частности, представители филотипов Firmicutes, Bacteroidetes, Proteobacteria, Verrucomicrobia, Actinobacteria [3].
В настоящее время внимание исследователей обращено к изучению микробиоты не только как важнейшего фактора пищеварения, но и как невидимого органа, ответственного за поддержание метаболического и иммунологического гомеостаза макроорганизма. Учитывая огромный вклад бактерий кишечника в процессы расщепления пищевых компонентов, их участие в синтезе и всасывании веществ, важных для регуляции липидного, углеводного и белкового обменов, поддержании иммунологической толерантности и барьерной функции, а также формировании противоинфекционной защиты, становится очевидна роль нарушений микробиоты в каскаде патологических реакций, лежащих в основе развития метаболических заболеваний. Большая часть отечественных и зарубежных работ в этой области посвящена вопросам функциональной роли микроорганизмов в развитии социально значимых патологий, таких как ожирение, сахарный диабет, неалкогольная жировая болезнь печени, а также канцерогенезе.
Распространенность ожирения достигла размеров эпидемии. Основной вклад в его развитие вносят особенности питания и низкая физическая активность. Однако существует еще ряд факторов, лидирующие позиции среди которых наряду со стрессами занимает состояние микробиома кишечника. Он реализует значимую часть патологических реакций, связанных с нарушением регуляции адипогенеза и аппетита [4–8]. К сожалению, интерпретация результатов исследований, демонстрирующих различие в составе кишечной микробиоты у тучных и индивидов с нормальной массой тела, усложняется подтверждением причинно-следственных связей между первичными факторами, приводящими к регистрируемым дисбалансам жирового обмена, и дисбиозом. Однако достоверно показано, что колонизация желудочно-кишечного тракта здоровых мышей микроорганизмами, полученными из дистальных отделов кишечника тучных особей, вызывала прирост жировой ткани на 60% и быстрое развитие инсулинорезистентности [5]. Транслокация микробиоты лиц с ожирением в кишечник стерильных животных также сопровождалась увеличением общей и жировой массы, несмотря на низкокалорийное питание [9].
Безусловно, ожирение – ключевой, а в ряде случаев и пусковой фактор прогрессирования нарушений углеводного и липидного обменов. Следовательно, очевидна связь между составом микробиоты и метаболическим дисбалансом. Биотехнологический и физико-технический прогресс, наблюдающийся последнее десятилетие, позволил предпринять активные попытки по расшифровке функциональной связи микроорганизмов и факторов поддержания метаболического гомеостаза.
Согласно данным литературы, симбиоз макроорганизма и микробиоты в отношении липидного и углеводного обменов реализуется благодаря нескольким основным механизмам. Одна из гипотез – вовлечение микроорганизмов в синтез и метаболизм короткоцепочечных жирных кислот (КЖК), а именно масляной, уксусной и пропионовой (бутирата, ацетата и пропионата соответственно). КЖК являются производными продуктов расщепления растворимых полисахаридов. Они стимулируют рост и пролиферацию энтероцитов, выполняют роль основного энергетического субстрата для клеток кишечника [10]. Механизм влияния бутирата на регуляцию энергопотребления и углеводный обмен базируется на способности активировать специфические рецепторы, сопряженные с G-белками, или серпентины, широко представленные в адипоцитах, иммунных и эпителиальных клетках кишечника [11]. Активация данных рецепторов индуцирует секрецию пептида YY в подвздошной и толстой кишке, способствующего снижению перистальтики кишечника и оказывающего влияние на центральную регуляцию аппетита [12, 13]. Активация серпентиновых рецепторов также приводит к стимуляции секреции глюкагоноподобного пептида 1 (ГПП-1). Функциональные возможности ГПП-1 подробно описаны в литературе: стимулирование постпрандиального выброса инсулина, снижение секреции глюкагона, ингибирование моторики кишечника. Это обусловило широкое применение агонистов рецепторов ГПП-1 в лечении ожирения и сахарного диабета 2-го типа [14]. Непосредственная роль рецепторов, сопряженных с G-белками, в регуляции энергетического обмена продемонстрирована на мышиных моделях. Так, у мышей с дефицитом вышеуказанных рецепторов развивалось ожирение, несмотря на нормокалорийное питание. В то же время у животных с гиперэкспрессией серпентинов не наблюдалось изменения массы тела на фоне диеты с высоким содержанием жиров. Вклад микробиоты в активацию данных рецепторов – изменение концентрации свободных жирных кислот. Ее роль в указанном процессе также подтверждается тем, что в условиях стерильности кишечника или после антибактериальной терапии вне зависимости от выраженности экспрессии серпентинов у мышей была зафиксирована нормальная масса тела [11].
Короткоцепочечные жирные кислоты также вовлечены в регуляцию кишечного глюконеогенеза. Так, бутират и пропионат активируют экспрессию генов, ответственных за реализацию синтеза глюкозы в слизистой оболочке кишечника через циклический аденозинмонофосфат-зависимый механизм и активацию оси «кишечник – головной мозг» посредством рецептора жирных кислот 3. В эксперименте у здоровых особей это оказывало положительный эффект на массу тела и контроль глюкозы. При этом у особей, нокаутированных по гену, кодирующему один из ключевых ферментов глюконеогенеза, аналогичных изменений не наблюдалось, несмотря на модификацию кишечной микробиоты [15, 16]. Таким образом, метаболически позитивное влияние короткоцепочечных жирных кислот реализуется только в условиях нормального кишечного глюконеогенеза.
Протективное действие КЖК в отношении поддержания барьерной функции эндотелия кишечника заключается в снижении проницаемости интестинального барьера. Механизм последнего не установлен, однако экспериментально показано, что при добавлении бутирата и пропионата к бокаловидным клеткам, с одной стороны, происходила стимуляция экспрессии мРНК гена муцина, с другой – повышение транскрипции белка клаудина 1, одного из важнейших компонентов замыкающих контактов, устанавливающего параклеточный барьер, который контролирует потоки молекул в эпителиальном межклеточном пространстве [17, 18].
В качестве еще одного механизма вовлечения микробиоты в энергетический гомеостаз следует рассматривать систему интеграции микроорганизмов в реализацию иммунного ответа и системного воспаления. Результаты ряда последних исследований свидетельствуют о роли КЖК в активации местного иммунитета посредством регуляции дифференцировки Т-клеток [19]. Одним из факторов, посредством которого микроорганизмы участвуют в регуляции иммунного статуса, является повышение концентрации ранее упомянутых короткоцепочечных жирных кислот. Данные метаболиты модулируют синтез и дифференциацию Т-клеток, секретирующих интерлейкины (ИЛ), которые выполняют важнейшую роль в регуляции аутоиммунитета, оказывая протективное воздействие на бета-клетки поджелудочной железы, и способствуют улучшению глюкозного и липидного метаболизма, а также снижению инсулинорезистентности [20]. Проанализирована связь между дисбиозом кишечника и снижением уровня ИЛ-18 в условиях подавления генов, ответственных за формирование протеинов, входящих в комплекс инфламмасом, – ключевых факторов, способствующих трансформации провоспалительных цитокинов в активные формы. Другими факторами микробиотической жизнедеятельности, вовлеченными в процесс иммунного ответа, считаются липополисахариды (ЛПС). Они являются основными компонентами клеточной стенки грамотрицательных бактерий и играют ключевую роль в развитии острой и хронической форм воспаления [21]. Избыточное содержание липополисахаридов в плазме, или эндотоксемия, впервые охарактеризовано P.D. Cani и соавт. как фактор развития дислипидемии на фоне диеты с высоким содержанием жиров. У особей, которым длительное время подкожно вводили ЛПС, отмечалось снижение численности бактерий типа Bacteroides, повышение тощакового уровня глюкозы и развитие инсулинорезистентности [21]. Аналогичные результаты получены M. Lassenius и соавт. В ходе исследования у пациентов с сахарным диабетом 1-го типа была выявлена ассоциация между уровнем ЛПС в крови и повышением уровня триглицеридов, диастолических показателей артериального давления. В другом исследовании, включившем более тысячи пациентов, уровень эндотоксемии был значимо выше в группе сахарного диабета [22].
Воздействие микробиоты на энергетический гомеостаз реализуется посредством влияния микроорганизмов на качественный и количественный состав желчных кислот. Известно, что пропорциональные характеристики последних значимо отличаются у здоровых особей после применения антибиотиков [23]. В настоящее время значение желчных кислот в регуляции энергетического обмена до конца не определено, однако установлено, что они являются лигандами для ряда важнейших рецепторов, таких как фарнезоидный Х-рецептор и мембранный рецептор желчных кислот, активация которых модифицирует глюкозный, липидный и энергетический обмены [24].
Одной из причин развития ожирения и ассоциированных с ним заболеваний являются особенности пищевого поведения. В данном контексте перспективным представляется изучение взаимосвязи микробиоты с эндоканнабиоидной системой. Данная система представлена каннабиоидными (КБ) рецепторами 1 и 2, эндогенными каннабиоидами и ферментами, участвующими в процессе их биосинтеза и деградации. Установлено, что эндоканнабиоиды влияют на энергообмен как с помощью центральных орексигенных механизмов на уровне лимбической системы, гипоталамуса, так и с помощью регуляции липогенеза [25].
Модификация микробиоты кишечника способствует снижению массы тела у тучных особей и экспрессии мРНК рецептора КБ-1. Применение антагонистов данного рецептора вызывало значимое снижение проницаемости стенки кишечника и уровня липополисахаридов в плазме [26]. Среди других биологически активных субстанций, вовлеченных в регуляцию углеводного обмена, а именно секреции инсулина и функционирование бета-клеток, выделяют гамма-аминомасляную кислоту (ГАМК). В экспериментах на клетках продемонстрировано усиление пролиферации бета-клеток и секреции инсулина при добавлении лакто- и бифидобактерий, способных к синтезу ГАМК (рис.) [27, 28].
Учитывая тесную связь между микробиотой и развитием ожирения, инсулинорезистентности, нарушением углеводного и липидного обменов, предпринимались попытки выявить виды или филотипы микроорганизмов, ответственных за нарушение метаболического гомеостаза.
Некоторые авторы связывают развитие ожирения и ассоциированных с ним заболеваний с изменением качественного состава микробиоты. Благодаря суммарному анализу всей совокупности генетического материала микроорганизмов кишечника удалось выделить две группы индивидов в зависимости от распределения и количества бактериальных генов. Первая группа характеризуется достаточным биологическим разнообразием (высокий геномный индекс) и преобладанием Actinobacteria и Euryarchaeota, а также противовоспалительных видов Faecalibacterium prausnitzii. Вторая – низким геномным индексом и распространенностью условно провоспалительных видов Bacteroidetes, Ruminococcus gnavus, ассоциированных с язвенным колитом и болезнью Крона. Кроме того, у данной группы отмечались значимо более высокие уровни лептина, адипонектина, триглицеридов, свободных жирных кислот, С-реактивного белка. Несмотря на очевидную связь качественного состава микробиоты и показателей энергообмена, четкая корреляция геномного индекса и ожирения в обеих группах отсутствовала, что, вероятно, обусловлено влиянием собственного генома на развитие метаболических нарушений [29]. N. Larsen и соавт. установили, что, несмотря на отсутствие различий в суммарной совокупности бактериального пула тонкой кишки, доля филотипа Firmicutes у пациентов с избыточной массой тела и сахарным диабетом 2-го типа ниже, чем у лиц без нарушений углеводного обмена [30]. В более масштабном исследовании, проведенном J. Qin и соавт., у пациентов с сахарным диабетом 2-го типа наблюдались дисбиотические нарушения – снижение численности бутират-продуцирующих бактерий при незначительном повышении количества условно патогенных микроорганизмов [31].
Другая гипотеза, также основывающаяся на видовой дифференциации микробиоты у лиц с ожирением и нарушениями разных видов обмена, связана с дисбиозом кишечника, при котором при неизменном потреблении пищевых компонентов микробиота обладает большей способностью к энергоизвлечению [32, 33]. Показано, что у мышей ob/ob с генетически детерминированным ожирением значимо преобладало количество коротких бактериальных ДНК-последовательностей, кодирующих ферменты, вовлеченные в расщепление полисахаридов и крахмала, а также генов, ответственных за синтез белков-переносчиков, обеспечивающих мембранный транспорт продуктов гидролиза (свидетельство высокой интенсивности обмена). В ранее упомянутом исследовании J. Qin и соавт. также было выявлено повышение трансмембранного транспорта сахаров, уровня метаболизации метана и деградации ксенобиотиков, что поддерживало концепцию микробиотического усиления энергоизвлечения из нутриентов у лиц с избыточной массой тела [31].
Микробиота также участвует в синтезе важнейших витаминов – кофакторов, необходимых для реализации метаболического конвейера по преобразованию энергии пищи в главный источник энергетических запасов организма аденозинтрифосфат [34]. Так, тиамин (витамин В1) выполняет роль кофактора в ферментативных реакциях расщепления пировиноградной кислоты, рибофлавин (витамин В12), ниацин (витамин В3), пантотеновая кислота (витамин В5) участвуют в окислительно-восстановительных биохимических процессах цикла Кребса. Биотин (витамин В7) является кофактором при метаболизме жирных кислот, лейцина и глюконеогенезе. Некоторые штаммы лактобактерий способны к синтезу фолатов, в связи с чем вызывают огромный интерес как альтернатива синтезированной фолиевой кислоте [35]. Микробиота участвует в метаболизме и синтезе 25(ОН)D, а также его производных посредством изменения концентрации фактора роста фибробластов 23 [36].
Учитывая вышесказанное, представляется логичным поиск потенциальных факторов, модулирующих количественный и качественный состав бактерий кишечника, с целью максимального извлечения преимуществ из симбиоза микроорганизмов и макроорганизма. Одним из таких факторов являются пробиотики. Согласно определению Всемирной организации здравоохранения, к пробиотикам следует относить «живые микроорганизмы», при назначении которых в корректных количествах наблюдаются положительные эффекты на организм.
Биологические и клинические эффекты пробиотиков основаны на коррекции качественного состава микробиоты с восстановлением пропорционального количества «метаболически полезных» бактерий. Наиболее перспективными в отношении влияния на метаболизм человека являются пробиотики, которые содержат лактобактерии и бифидобактерии.
Биологическое действие пробиотиков достигается путем активации или коррекции механизмов, посредством которых микробиота регулирует метаболический обмен и иммунный статус.
В первую очередь пробиотики и входящие в их состав бактерии характеризуются высокой способностью продуцировать КЖК. Это достигается путем ферментации конечных продуктов расщепления углеводов [37]. Показано, что лактобактерии способны к синтезу всего спектра КЖК (пропионата, бутирата, ацетата), бифидобактерии – лактата и ацетата.
Влияние пробиотиков на концентрацию конечных продуктов распада углеводов продемонстрировано при проведении экспериментов в специализированной системе – функциональном аналоге пищеварительного тракта человека (Simulator Human Microbial Ecosystem – SHIME) [38]. Механизм позитивного влияния короткоцепочечных жирных кислот на углеводный и липидный обмены описан ранее.
Пробиотики также воздействуют на синтез и секрецию интестинальных пептидов, участвующих в регуляции углеводного обмена. В настоящее время выделены виды лактобактерий, способных секретировать ГПП-1. У мышей с диабетом при добавлении в пищу вышеуказанных микроорганизмов отмечалось значимое улучшение гликемических показателей за счет повышения уровня инсулина [39]. Это может послужить основанием для изучения факторов, лежащих в основе профилактики нарушений углеводного обмена, и возможностей использования пробиотиков в качестве антидиабетических препаратов. Как указывалось ранее, лактобактерии способны синтезировать ГАМК и модулировать эндоканнабиоидную систему.
В настоящее время представлены результаты более 50 исследований, посвященных влиянию пробиотиков на микробиоту кишечника, липидный и углеводный обмены, артериальное давление. Анализ данных литературы о применении и эффектах пробиотиков свидетельствует, что в значимом количестве рандомизированных клинических исследований (РКИ) подтверждено положительное влияние модификации микробиоты на течение сахарного диабета 1-го и 2-го типов, ожирение [40–45]. Так, в 2016 г. C. Li и соавт. провели метаанализ, по результатам которого продемонстрировано значимое снижение уровня глюкозы плазмы натощак при применении пробиотиков (стандартизованная разность средних (СРС) 0,61 ммоль/л), уровня холестерина высокой плотности (СРС 0,42 ммоль/л) без значимых различий в показателях триглицеридов, гликированного гемоглобина, индекса инсулинорезистентности, холестерина низкой плотности [46]. В том же году аналогичные результаты представлены Q. Zhang и L. Hua. Исследователи отметили более выраженные эффекты пробиотических препаратов в отношении снижения тощаковых уровней глюкозы и повышения инсулина [47]. H. Jun и соавт. установили, что пробиотики по сравнению с плацебо значимо снижали как лабораторные показатели липидного обмена (общего холестерина (СРС 0,57 мг/дл), липопротеинов низкой плотности (СРС 0,4 мг/дл), триглицеридов (СРС 0,66 мг/дл)) при повышении уровня липопротеинов высокой плотности (СРС 0,38 мг/дл), так и значения артериального давления [48], что в совокупности свидетельствовало о роли пробиотиков в уменьшении риска развития сердечно-сосудистых осложнений у данной категории пациентов. Таким образом, хотелось бы подчеркнуть, что пробиотики, которые ранее продемонстрировали эффективность в лечении заболеваний желудочно-кишечного тракта, согласно современным данным, обладают широким спектром положительных свойств в отношении коррекции компонентов метаболического синдрома (например, штаммы, указанные в таблице).
В настоящее время накоплено значительное количество данных, свидетельствующих о колоссальной роли микроорганизмов кишечника в регуляции энергетического гомеостаза. В большей степени влияние микробиоты на систему углеводного и жирового обменов реализуется посредством модуляции синтеза свободных жирных кислот, желчных кислот, ряда системных биоактивных молекул, клеточного звена иммунной системы и проницаемости кишечной стенки для липополисахаридов.
Поддержание корректного количественного и качественного состава микробиоты, в том числе с использованием пробиотических препаратов, необходимо для нормального функционирования не только желудочно-кишечного тракта, но и системы энергетического гомеостаза. Суммированные данные систематических обзоров, включивших рандомизированные клинические исследования, свидетельствуют о широких перспективах в отношении применения пробиотиков как для профилактики нарушений углеводного обмена, так и для коррекции уже имеющихся заболеваний, в частности сахарного диабета 2-го типа и дислипидемии.
При поддержке компании ООО «Пфайзер Инновации»
Роль микробиоты в регуляции гомеостаза организма человека при инфекции | Бухарин
Взаимодействия «паразит–хозяин» микробов и человека весьма разнообразны и нередко нарушают гомеостаз хозяина, т.е. стабильное внутреннее равновесие функционирующих систем организма. С другой стороны, имеется немало примеров, когда микробные клетки оказываются полезны для сохранения здоровья человека. Однако в этом «союзе, неотделимом от вражды», который длится уже много веков, есть свой «микробный орган» — микробиом, которым Природа наделила человека, защищая все его биотопы. Как же это осуществляется и что в «копилке» исследователей?
Метаболическая интеграция и сигнальные молекулы
Обилие разнообразных сигнальных молекул и метаболитов в кишечнике позволяет микробиоте осуществлять влияние на состояние организма хозяина, формирование его гомеостаза и управление поведением. Регуляторные метаболиты микроорганизмов включают короткоцепочечные жирные кислоты, гамма-аминомасляную кислоту, биотин, витамин К, путресцин, спермидин, спермин, таурин, кадаверин, триптофан и др. [1][2][3][4].
Была обоснована интеграция метаболизма человека и его микробиоты на основе обобщения результатов исследований с участием микробных метаболитов в развитии критических состояний [5], где было показано, что в сложившейся системе человек–микробиом присутствуют все необходимые объективные условия для формирования метаболической интеграции. Особого внимания заслуживает группа микробных экзометаболитов, имеющих ароматическое строение. Их анализ выявил около 50 ароматических соединений в кишечнике здорового человека, в количественном отношении преобладали такие метаболиты, как фенилуксусная кислота, гидроксифенилуксусная кислота, фенилпировиноградная кислота и др. В сыворотке крови здоровых людей обнаружено присутствие большинства этих ароматических аминокислот с преобладанием гидроксифенилуксусной кислоты. Изменение соотношения ароматических аминокислот в крови авторы связывают с их избирательной утилизацией клетками тканевых барьеров, хотя это не исключает потребности в метаболитах кишечной доминантной микрофлоры.
Имеются данные, подтверждающие роль опиатов в инфекционном процессе [6][7]. Экспериментальные материалы показывают, что при стрессе у лабораторных животных появляются опиаты в просвете кишечника, что сопровождается активацией вирулентности кишечной палочки и нарушением барьерной функции кишечного эпителия [8]. Оказалось, что динорфин — представитель группы опиатов, не увеличивает ростовые свойства, но усиливает продукцию пиоцианина у псевдомонад. А это является еще одним доказательством влияния эукариотических «сигнальных молекул» на физиологию прокариот без изменения их роста/размножения [9].
В ряде работ также описано снижение вирулентности микроорганизмов под действием сигнальных молекул иммунной системы, посредством нарушения микробного кворума, как это было показано на примере динорфина и интерферона (INF)-γ [8][9][10]. В работе M.W. Bader и соавт. [11] представлены данные о влиянии пептидных гормонов, имеющих структурную гомологию к антибактериальным пептидам, на микроорганизмы. Предполагается, что эти молекулы обладают вторичной антимикробной активностью, помимо их мишень-специфического взаимодействия с клетками эукариот.
Натрийуретические пептиды в настоящее время также рассматриваются как пептиды с антимикробным действием, которые могут оказывать влияние на микробиоту при инфекционном процессе [10]. В пользу этого свидетельствуют данные о формировании пор в мембране бактерий под действием С-типа натрийуретического пептида и увеличение концентрации мозгового натрийуретического пептида при септическом шоке. В ряде работ установлено, что натрийуретические гормоны типа В и С стимулируют вирулентные свойства псевдомонад, не влияя на их ростовые характеристики, но изменяя внутриклеточную концентрацию цАМФ. Считается, что механизм данного действия натрийуретических гормонов опосредован белком Vfr, связывающим цАМФ и контролирующим выработку различных факторов вирулентности у Pseudomonas aeruginosa. Исследования штаммов псевдомонад показали наличие рецепторов к разным подтипам натрийуретических гормонов, действующих как через цАМФ, так и через цГМФ [12][13][14].
Гипоталамические нонапептиды
Всеобщий исследовательский интерес к окситоцину и вазопрессину не случаен. Являясь продуктом гипоталамо-гипофизарной нейросекреторной системы (ГГНС) головного мозга, его супраоптического и паравентрикулярного ядер, окситоцин, как и вазопрессин, обладает широтой физиологических действий и принимает непосредственное участие в регуляции адаптационных реакций организма человека [15]. Особенно наглядно это выявляется при инфекции, когда окситоцин защищает хозяина от возбудителя. Ранее считали, что препарат не оказывает защитный эффект непосредственно, хотя и была обнаружена его способность усиливать антимикробное действие антибиотиков, применяемых в комбинации с окситоцином [16].
Так в чем же секрет защитного эффекта при инфекционной патологии? Что он делает с возбудителем инфекции? Для этого вернемся к ранним работам по регуляции персистентного потенциала бактерий О.Л. Черновой (1989) [17], которая, изучая влияние различных антисептических фармакологических средств на антилизоцимную активность золотистого и эпидермального стафилококков, показала, что лидером в десятке изученных антисептиков — препаратов, подавляющих антилизоцимную активность бактерий, — оказался окситоцин, что позволило обратить внимание на ингибирование этого персистентного признака микроорганизмов. В последующем Д.А. Кириллов (2004) [18] методом клонального анализа популяций различных возбудителей инфекции показал, что окситоцин перестраивает персистентный потенциал клонов популяции возбудителя вплоть до его элиминации из организма хозяина.
Эти работы «проторили дорожку» в XXI в. — век инфектологии (от микробиологии и иммунологии), изучающей взаимоотношения паразит–хозяин при инфекции на основе симбиотической платформы и клонального анализа персистентного потенциала популяции возбудителя. Таким образом, клональная перестройка популяции возбудителя болезни, снижающая его адаптационные возможности, — это существенный механизм защиты хозяина, реализуемый при помощи окситоцина.
Не исключено, что данный механизм защиты хозяина от инфекций — еще одна иллюстрация из разряда «природоподобных технологий», которые нам еще предстоит освоить. Но это дело времени и смелости ума. А основания для этого есть.
Посильную лепту в изучение защитного эффекта окситоцина внесли и иммунологи, описавшие другие механизмы опосредованного защитного действия окситоцина при инфекции: фагоцитарную функцию макрофагов, усиление бластной трансформации лимфоцитов, ингибирование биопленкообразования патогенов [19].
Обсуждая эту проблему, нельзя не упомянуть инсулиноподобный эффект окситоцина, базирующийся на усилении синтеза гликогена из глюкозы. Хирурги хорошо пользуются этим приемом, применяя окситоцин на фоне сахарного диабета пациентов при гнойно-некротических поражениях стоп и гнойно-воспалительных заболеваниях мягких тканей [19][20][21].
Универсальные регуляторные эффекты нонапептидов нашли свое применение и при панкреонекрозе, деструктивном панкреатите и системных поражениях поджелудочной железы [22][23].
Исследователи не теряют интереса к фундаментальным проблемам медицины. Это в полной мере относится к проблеме гомеостаза организма. Как и чем мы можем помочь пациенту?
Лучшее свидетельство тому — исследование, выполненное в «школе» академика Ю.В. Наточина, где был выявлен новый механизм работы многоцелевого регулятора гомеостаза — окситоцина, определивший новую функциональную роль нонапептида — его участие в осморегуляции организма. При изучении регуляции водовыделительной функции почек отмечено, что после водной нагрузки при гипергидратации у крыс возрастала секреция окситоцина ГГНС, что способствовало усилению водного диуреза и приводило к более быстрому выделению воды почкой и восстановлению осмотического гомеостаза [24].
Адипокины и цитокины
Многочисленные исследования механизмов взаимосвязи метаболических нарушений и воспалительных процессов привели к признанию факта, что жировая ткань функционирует как эндокринный орган, выделяя различные биологически активные вещества (адипокины). Изучение адипокинов показало, что дисбаланс данных про- и противовоспалительных медиаторов приводит к различным метаболическим дисфункциям, что свидетельствует о роли адипокинов в формировании гомеостаза хозяина [25]. Принимая во внимание участие адипокинов в воспалении, эти пептиды были разделены на провоспалительные (лептин, резистин, интерлейкин (IL)-6, фактор некроза опухоли (TNF)-α) и противовоспалительные (адипонектин, антагонист рецептора IL-1, IL-10) [26].
Лептин обладает множественным действием и влияет на гипоталамус, осуществляя гормональную регуляцию, связанную с поступлением питательных веществ и энергетическим метаболизмом, а также влияет на метаболизм глюкозы, липидов и другие функции человека [27]. Одной из важных функций лептина является регуляция иммунного ответа, что предполагает роль данных пептидов в интегративных механизмах ассоциативного симбиоза человека и микроорганизмов.
В настоящее время этот вопрос активно изучается. Еще не выяснена защитная роль адипокинов при инфекции толстого кишечника с участием лептина, индуцирующего продукцию муцина за счет стимуляции эпителиальных клеток ободочной кишки и таким образом обеспечивающего статический внешний барьер против патогенов. Однако при этом бактериальная инвазия Salmonella typhimurium, наличие эндотоксина сальмонелл или кишечной палочки не влияли на уровень лептина в крови. Напротив, введение токсина Clostridioides difficile лабораторным животным вызывало значительное повышение уровня лептина в плазме крови и усиливало экспрессию рецепторов к лептину на клетках слизистого эпителия, что предполагает прямой провоспалительный эффект лептина в кишечнике [28][29].
Известно, что диарея, возникающая при бактериальной кишечной инфекции, связана с воздействием микробных липополисахаридов на иммунную систему и нарушением моторики желудочно-кишечного тракта. В экспериментах in vivo выявлено, что у мышей, получавших грелин, нарушение моторики, вызванное присутствием эндотоксина в крови, корректировалось за счет ингибирования уровня оксида азота в желудочно-кишечном тракте и уменьшения продукции провоспалительных цитокинов IL-1β и TNF-α, а также за счет индукции противовоспалительного цитокина IL-10 [30, 31].
Интеграция микробиоты с организмом хозяина может осуществляться при помощи сигнальных молекул иммунной системы человека — цитокинов, баланс которых является одним из условий формирования гомеостаза человека, поскольку цитокины принимают непосредственное участие в регуляции иммунного ответа при инфекции [32]. При этом изменение цитокинового баланса происходит не только за счет взаимодействия микробиоты с клетками иммунитета, но и при непосредственном влиянии бактерий на цитокины (антипептидная активность). Также известно влияние самих цитокинов на биологические свойства микроорганизмов. В экспериментах in vitro показана стимуляция ростовых свойств бактерий под действием IL-1, IL-2, IL-6, INF-γ, TNF-α. У культуры Yersinia pestis обнаружены мембранные рецепторы (антиген сборки капсулы F1), связывающие IL-1β, а у Р. aeruginosa — белок, специфически связывающийся с INF-γ, что приводило к активации механизмов «quorum sensing» [33][34][35][36][37].
Получены данные о ферментах бактерий, расщепляющих многие виды органических макромолекул, включая цитокины IL-2, INF-γ [38], которые могут свидетельствовать о том, что инактивация цитокинов, являющихся продуктом иммунных клеток (лимфоцитов, макрофагов и др.), может привести к нарушениям механизмов как врожденного, так и адаптивного иммунитета. Подтверждением модификации цитокинов и их рецепторов служат материалы, свидетельствующие, что ауреолизин золотистого стафилококка, являющийся металлопротеазой, может вызвать деградацию рецепторов к IL-6 на клетках, цистеиновая протеаза Streptococcus pyogenes разрушает IL-1β, а сериновая протеаза — IL-8. Кроме того, цистеиновая протеаза Porphyromonas gingivalis может вызвать деградацию целой группы цитокинов, включая IL-8, -1β, -6, -12, INF-γ, TNF-α, а металлопротеаза Р. aeruginosa способна разрушать IL-2, IL-6 [39][40].
Таким образом, полученные «находки» иллюстрируют способность микробиоты не только влиять на продукцию цитокинов иммунными клетками, но и использовать определенные цитокины в качестве ростовых факторов и медиаторов «чувства кворума», а также проявлять антипептидную активность, внося свой вклад в формирование цитокинового баланса в организме человека.
Несомненно, что взаимодействие микробных сигнальных метаболитов и иммунной системы человека представляет интерес с позиции интеграции молекулярных систем про- и эукариот при ассоциативном симбиозе человека. В ряде работ показано, что различные гомологи ацилгомосерин-лактонов (АГЛ) ускоряют апоптоз макрофагов и нейтрофилов, ингибируют пролиферацию лимфоцитов и выработку TNF-α и IL-12, тормозят Т-клеточный ответ, индуцируют апоптоз в дендритных клетках и CD4+ Т-лимфоцитах. Роль АГЛ подтверждается наличием механизмов, ограничивающих количество сигнальных молекул в среде, что названо «тушением кворума». Снижение концентрации АГЛ контролируется со стороны как микробиоты, так и хозяина. К примеру, бациллы в ответ на увеличение количества АГЛ продуцируют фермент, инактивирующий широкий спектр АГЛ путем расщепления лактонового кольца. Клетки дыхательного эпителия млекопитающих также производят АГЛ-инактивирующие ферменты (параоксоназы), деградирующие АГЛ синегнойной палочки. В другой работе показано существование трех семейств параоксоназ, которые у млекопитающих расположены преимущественно в печени и могут инактивировать разнообразные АГЛ [41][42][43][44][45][46].
При изучении механизмов интеграции микробиоты и хозяина интерес также представляют сигнальные молекулы микроорганизмов. Их влияние на иммунитет человека было показано на примере гомологов алкилоксибензолов, когда под влиянием метилрезорцина изменялись функциональная активность и субстратная специфичность лизоцима [47].
Очевидно, что в механизмах интеграции микробиоты и человека задействованы сигнальные молекулы: со стороны микробиоты — низкомолекулярные метаболиты, молекулы «quorum sensing» и пр., а со стороны хозяина — гормоны и медиаторы иммунитета. По-видимому, в условиях симбиотических взаимоотношений микробиота–хозяин формируется единая регуляторная среда, в которой наблюдается многообразие создающихся связей: от непосредственных (прямых) взаимодействий — разрушения сигнальных молекул (инактивации молекул кворума, разрушения антимикробных факторов иммунитета), индукции физиологических функций за счет наличия схожих рецепторов к лигандам и, наконец, модификации сигнальных молекул (расширения спектра имеющихся антимикробных ферментов, появления антимикробной активности у пептидов, ранее не имевших данного свойства) — до косвенных воздействий, опосредованных активацией и регуляцией системы иммунитета через цитокиновую сеть и систему адипокинов. По-видимому, сочетание этого многообразия механизмов интеграции в единой регуляторной среде приводит к формированию гомеостаза, означающего динамическое равновесие сигнальных систем микробиоты и человека в условиях ассоциативного симбиоза [46].
Бифидофлора кишечного биотопа — «форпост» здоровья человека
Роль микробного «органа» (микробиома) трудно переоценить, и уж если он создан Природой и сосуществует с хозяином много веков, то остается лишь понять его физиологическое назначение. Наличие в организме млекопитающих универсального и древнего «центра управления» — гипоталамо-гипофизарной системы, продуцирующей нонапептидные нейросекреторные гормоны (вазопрессин и окситоцин), предполагает, что они не могут остаться без работы [48][49].
Оказалось, что кишечная микрофлора, стимулируя иммунную защиту хозяина, защищает организм от раневой инфекции. С одной стороны, эта защита может осуществляться за счет транслокации полезной микрофлоры хозяина, как это было показано на примере бацилл [50]. С другой стороны, микробные компоненты (клетки и метаболиты), формируя кишечно-мозговую ось, могут влиять на выработку гипоталамического гормона — окситоцина. Работы по изучению влияния бактерий на секрецию окситоцина малочисленны и проведены на модели лактобактерий. Установлено, что лактобациллы стимулируют продукцию окситоцина, что благотворно отражается на заживлении инфицированных ран в эксперименте [51]. Также отмечено, что Lactobacillus spp. стимулируют окситоцин, который регулирует экспрессию INF-γ и CD25 для иммунной толерантности. Все эти усилия предупреждают избыточную реактивность как своих, так и внешних факторов среды, которые способствуют преждевременному старению организма. На моделях мышей показана эффективность индуцированных лактобациллами и их клеточными лизатами T-reg при участии нейропептидного гормона окситоцина [51–53].
Приведенные материалы вкупе с описанными нашими данными свидетельствуют, что микробиом усиливает регуляцию окситоцина, тем самым улучшая течение раневой инфекции, способствуя быстрейшему заживлению ран [19].
В свете обсуждаемой проблемы определенный интерес представляют данные оренбургских исследователей ИКВС УрО РАН, проводящих изучение биологических характеристик бифидофлоры в качестве ключевого регулятора здоровья человека.
Систематическое изучение микросимбиоценоза кишечного биотопа у человека позволило выявить феномен микробного распознавания свой– чужой в условиях взаимодействия доминантных (бифидофлора) и ассоциативных микросимбионтов [46].
Известно, что, независимо от уровня сложности, любые живые организмы (от прокариот до высших эукариот) имеют различные механизмы защиты от чужеродной информации, поскольку концепция «своего» тесно связана с самоидентификацией и саморегуляцией любой биологической системы [54].
Микробное распознавание и механизмы самоидентификации бактерий активно изучаются. L.M. Wenren с соавт. [55] в результате исследования роста культур Proteus mirabilis на поверхности агаровых сред отметили, что взаимоотношения микроорганизмов в бульонной культуре могут отличаться от таковых в модели «агаровой среды», поскольку в этом процессе имеют значение микробные метаболиты. A.E. Shank и соавт. [56] связывали регуляторные взаимодействия микроорганизмов с наличием в супернатанте сигнальных молекул. Очевидно, что изменение фенотипа микробных популяций при межмикробном взаимодействии осуществляется с помощью различных молекул, далее использующихся микробиотой в качестве индукторов новых метаболитов-посредников, что в конечном итоге оказывает влияние на формирование антагонистических либо синергидных связей между микроорганизмами [46].
С использованием приема индукции микробных метаболитов в условиях пары доминант–ассоциант был выявлен феномен оппозитного (усиление/подавление) влияния микросимбионтов на их биологические свойства (антагонистический, персистентный потенциал и способность к формированию биопленок), позволяющий реализовать принцип «свой–чужой» в условиях микросимбиоценоза. Дальнейшее развитие исследований по определению «чужеродности» штаммов микроорганизмов позволило определить биосовместимость бактерий в микробной композиции и оценить эффективность пробиотических препаратов [46].
Используя симбиотический подход на платформе нового направления «инфекционная симбиология», было определено, что не только организм хозяина, посредством различных механизмов врожденного и адаптивного иммунитета, выявляет и уничтожает «чужеродные» штаммы бактерий и грибов, но и сами микроорганизмы (представители доминантной микробиоты) способны определять «свои» и «чужие» виды микросимбионтов в составе микросимбиоценоза. Таким образом, своеобразная перестройка «микробного органа» человека позволяет микробиоте сформировать симбиотические связи для поддержания стабильного функционирования микросимбиоценоза на оптимальном уровне с целью выживания нормофлоры в той экологической нише человека, которую она занимает.
Заключение
Оценивая ретроспективу рассмотренного вопроса, можно сделать вывод о целесообразности продолжения накопления материала по выяснению механизмов защиты организма хозяина при помощи микробных клеток и их продуктов. Активно изучается возможность использования в качестве регуляторов гомеостаза организма человека сигнальных молекул, гормонов и цитокинов. Это очень интересная многообещающая тематика по выявлению новых «природоподобных» технологий, которые нам предстоит еще открыть, но учиться у Природы не зазорно.
Появление в третьем тысячелетии науки инфектологии значительно расширило рамки изучения отношений «паразит–хозяин» с включением симбиотического подхода на организменном и клональном уровнях персистентного потенциала патогенов.
Это позволило выявить роль кишечной микробиоты в регуляции гомеостаза хозяина через треугольник «кишечная микробиота–ГГНС–окситоцин» [19]. К этому можно присовокупить материалы, уточняющие биоэффекты данного универсального ключевого регулятора гомеостаза:
1) более быстрое заживление ран;
2) поддержание костно-мышечной массы тела человека;
3) улучшение ментального здоровья;
4) психотропное действие, регуляция социальной памяти и когнитивных функций;
5) пониженный риск ожирения;
6) усиление репродуктивной активности и др.
Описанный треугольник «микробиота–ГГНС–окситоцин» подтвержден экспериментально-клиническими материалами и органично вписывается в концепцию «кишечно-мозговой оси», характеризующую ряд важнейших физиологических функций хозяина, существенно дополняя их.
1. Zheng X., Xie G., Zhao A., Zhao L., Yao C., Chiu N.H., et al. The footprints of gut microbial-mammalian cometabolism. J. Proteome Res. 2011; 10(12): 5512-22. https://doi.org/10.1021/pr2007945
2. Le Gall G., Noor S.O., Ridgway K. Metabolomics of fecal extracts detects altered metabolic activity of gut microbiota in ulcerative colitis and irritable bowel syndrome. J. Proteome Res. 2011; 10(9): 4208-18. https://doi.org/10.1021/pr2003598
3. Beloborodova N.V, Olenin A.Y., Fedotcheva N.I., Shubina V, Teplova V.V. Effect of phenolic acids originating from microbes on mitochondria and neutrophils. Crit. Care. 2012; 16(Suppl. 3): 26.
4. Matsumoto M., Kibe R., Ooga T., Aiba Y, Kurihara S., Sawaki E., et al. Impact of intestinal microbiota on intestinal luminal metabolome. Sci. Re: 2012; 2: 233. https://doi.org/10.1038/srep00233
5. Белобородова Н.В. Интеграция метаболизма человека и его микробиома при критических состояниях. Общая реаниматология. 2012; 8(4): 42-54.
6. Hooi D.S., Bycroft B.W., Chhabra S.R., Williams P., Pritchard D.I. Differential immune modulatory activity of Pseudomonas aeruginosa quorum-sensing signal molecules. Infect. Immun. 2004; 72(11): 6463-70. https://doi.org/10.1128/iai.72.11.6463-6470.2004
7. Pritchard D.I. Immune modulation by Pseudomonas aeruginosa quorum-sensing signal molecules. Int. J. Med. Microbiol. 2006; 296(2-3): 111-6. https://doi.org/10.1016/j.ijmm.2006.01.037
8. Wu L., Holbrook C., Zaborina O., Ploplys E., Rocha F., Pel¬ham D., et al. Pseudomonas aeruginosa expresses a lethal virulence determinant, the PA-I lectin/adhesin, in the intestinal tract of a stressed host: the role of epithelia cell contact and mole¬cules of the quorum sensing signaling system. Ann. Surg. 2003; 238(5): 754-64. https://doi.org/10.1097/01.sla.0000094551.88143.f8
9. Zaborina O., Lepine F., Xiao G., Valuckaite V, Chen Y, Li T., et al. Dynorphin activates quorum sensing quinolone signaling in Pseudomonas aeruginosa. PLoS Pathog. 2007; 3(3): 1-15. https://doi.org/10.1371/journal.ppat.0030035
10. Lesouhaitier O., Veron W., Chapalain A., Madi A., Blier A.S., Dagorn A., et al. Gram-negative bacterial sensors for eukaryotic signal molecules. Sensors (Basel). 2009; 9(9): 6967-90. https://doi.org/10.3390/s90906967
11. Bader M.W., Sanowar S., Daley M.E., Schneider A.R., Cho U., Xu W., et al. Recognition ofantimicrobial peptides by a bacterialsensor kinase. Cell. 2005; 122(3): 461-72. https://doi.org/10.1016/j.cell.2005.05.030
12. Vila G., Resl M., Stelzeneder D., Struck J., Maier C., Riedl M., et al. Plasma NT-proBNP increases in response to LPS administration in healthy men. J. Appl. Physiol. (1985). 2008; (105): 1741-5. https://doi.org/10.1152/japplphysiol.90442.2008
13. Veron W., Lesouhaitier O., Pennanec X., Rehel K., Leroux P., Orange N., et al. Natriuretic peptides affect Pseudomonas aeru-ginosa and specifically modify lipopolysaccharide biosynthesis. FEBS J. 2007; 274(22): 5852-64. https://doi.org/10.1111/j.1742-4658.2007.06109.x
14. Veron W., Orange N., Feuilloley M.G., Lesouhaitier O. Natri-uretic peptides modify Pseudomonas fluorescens cytotoxicity by regulating cyclic nucleotides and modifying LPS structure. BMC Microbiol. 2008; 8: 114. https://doi.org/10.1186/1471-2180-8-114
15. Стадников А.А., Бухарин О.В. Гипоталамическая нейросекреция и структурно-функциональный гомеостаз прои эукариот. Оренбург; 2012.
16. Стадников А.А. Роль гипоталамических нейропептидов во взаимодействии прои эукариот (структурно-функциональные аспекты). Екатеринбург; 2001.
17. Чернова О.Л. Антилизоцимная активность стафилококков, выделенных при бактерионосительстве: автореф. дисс. … канд. биол. наук. Челябинск; 1989. 17 с.
18. Кириллов Д.А. Лекарственная регуляция персистентных свойств микроорганизмов: автореф. дисс. … канд. мед. наук. Оренбург; 2004. 22 с.
19. Бухарин О.В., Стадников А.А., Перунова Н.Б. Роль окситоцина и микробиоты в регуляции взаимодействий прои эукариот при инфекции. Екатеринбург; 2018.
20. Widmaier U., Shah P.R., Lee G. Interactions between oxytocin, glucagon and streptozotocin induced diabetic rats. Regul. Pept. 1991; 34(3): 235-49. https://doi.org/10.1016/0167-0115(91)90182-g
21. Бухарин О.В., Курлаев П.П., Перунова Н.Б., Скоробогатых Ю.И. Экспериментальное изучение комбинации ципрофлоксацина с окситоцином на образование биоплёнок условно-патогенными бактериями. Журнал микробиологии, эпидемиологии и иммунобиологии. 2010; (6): 3-7.
22. Костюченко А.Л., Филин В.И. Неотложная панкреатология. СПб.: Деан; 2000.
23. Демидов В.М., Демидов С.М. Перспективы интрабурсального применения даларгина и сандостатина при лапароскопических вмешательствах у больных с панкреонекрозами. Анналы хирургической гепатологии. 2002; 7(1): 200.
24. Наточин Ю.В., Голосова Д.Р., Шахматова Е.И. Новая функциональная роль окситоцина участие в осморегуляции. Доклады Академии наук. 2018; 479(6): 712-5. https://doi.org/10.7868/S0869565218120228
25. Greenberg A.S., Obin M.S. Obesity and the role of adipose tis-sue in inflammation and metabolism. Am. J. Clin. Nutr 2006; 83(2): 461-5. https://doi.org/10.1093/ajcn/83.2.461s
26. Toussirot E., Streit G., Wendling D. The contribution of adipose tissue and adipokines to inflammation in joint diseases. Curr Med. Chem. 2007; 14(10): 1095-100. https://doi.org/10.2174/092986707780362826
27. El Homsi M., Ducroc R., Claustre J. et al. Leptin modulates the expression of secreted and membrane-associated mucins in colonic epithelial cells by targeting PKC, PI3K, and MAPK path-ways. Am J. Physiol. Gastrointest. Liver Physiol. 2007; 293(1): G365-73. https://doi.org/10.1152/ajpgi.00091.2007
28. Jenkins N.L., Turner J.L., Dritz S.S., Durham S.K., Minton J.E. Changes in circulating insulinlike growth factor-I, insulinlike growth factor binding proteins, and leptin in weaned pigs in-fected with Salmonella enterica serovar Typhimurium. Domest. Anim. Endocrinol. 2004; 26(1): 49-60. https://doi.org/10.1016/j.domaniend.2003.09.001
29. Mykoniatis A., Anton M., Wilk M., Wang C.C., Ungsunan L., Bluher S., et al. Leptin mediates Clostridium difficile toxin A-induced enteritis in mice. Gastroenterology. 2003; 124(3): 683-91. https://doi.org/10.1053/gast.2003.50101
30. Chen Y.T., Tsai S.H., Sheu S.Y. et al. Ghrelin improves LPS-induced gastrointestinal motility disturbances: roles of NO and prostaglandin E2. Shock. 2010; (33): 205-212.
31. Waseem T., Duxbury M., Ito H., Tsai L.H. Exogenous ghrelin modulates release of pro-inflammatory and anti-inflammatory cytokines in LPS-stimulated macrophages through distinct sig-naling pathways. Surgery. 2008; 33(2): 205-12. https://doi.org/10.1097/shk.0b013e3181ae841b
32. Lambert G. Stress-induced gastrointestinal barrier dysfunction and its inflammatory effects. J. Anim. Sci. 2009; 87(14 Suppl.): E101-8. https://doi.org/10.2527/jas.2008-1339
33. Zav’yalov V, Chernovskaya T.V., Navolotskaya E.V., Karlyshev A.V., MacIntyre S., Vasiliev A.M., et al. Specific high af-finity binding of human interleukin 1 beta by Caf1A usher pro-tein of Yersinia pestis. FEBS Lett. 1995; 371(1): 65-8. https://doi.org/10.1016/0014-5793(95)00878-d
34. Wu L., Holbrook C., Zaborina O., Ploplys E., Rocha F., Pelham D., et al. Pseudomonas aeruginosa expresses a lethal virulence determinant, the PA-I lectin/adhesin, in the intestinal tract of a stressed host: the role of epithelia cell contact and molecules of the quorum sensing signaling system. Ann. Surg. 2003; 238(5): 754-64. https://doi.org/10.1097/01.sla.0000094551.88143.f8
35. Романова Ю.М., Алексеева Н.В., Степанова Т.В., Разумихин М.В., Томова А.С., Шилов И.А. и др. Влияние фактора некроза опухоли на размножение вегетативных и некультивируемых форм сальмонелл. Журнал микробиологии, эпидемиологии и иммунобиологии. 2002; (4): 20-5.
36. Kanangat S., Meduri Gu., Tolley E.F., Patterson D.R., Meduri C.U., Pak C., et al. Effects of cytokines and endotoxin on the intracellular growth of bacteria. Infect. Immun. 1999; 67(6): 2834-40. https://doi.org/10.1128/iai.67.6.2834-2840.1999
37. Wilson M., Seymour R., Henderson B. Bacterial perturbation of cytokine networks. Infect. Immun. 1998; 66(6): 2401-9. https://doi.org/10.1128/iai.66.6.2401-2409.1998
38. Potempa J., Pike R.N. Corruption of innate immunity by bacterial proteases. J. Innate. Immun. 2009; 1(2): 70-87. https://doi.org/10.1159/000181144
39. Sheets S.M., Robles-Price A.G., McKenzie R.M., Casiano C.A., Fletcher H.M., et al. Gingipain-dependent interactions with the host are important for survival of Porphyromonas gingivalis. Front. Biosci. 2008; 13: 3215-38. https://doi.org/10.2741/2922
40. Leidal K.G., Munson K.L., Johnson M.C., Denning G.M. Metalloproteases from Pseudomonas aeruginosa degrade hyman RANTES, MCP-1, and ENA-78. J. Interferon. Cytokine. Res. 2003; 23(6): 307-18. https://doi.org/10.1089/107999003766628151
41. Tateda K., Ishii Y, Horikawa M., Matsumoto T., Miyairi S., Pechere J.C., et al. The Pseudomonas aeruginosa autoinducer N-3-oxododecanoyl homoserine lactone accelerates apoptosis in macrophages and neutrophils. Infect. Immun. 2003; 71(10): 5785-93. https://doi.org/10.1128/iai.71.10.5785-5793.2003
42. Telford G., Wheeler D., Williams P., Tomkins P.T., Appleby P., Sewell H., et al. The Pseudomonas aeruginosa quorum-sensing signal molecule N-(3-oxododecanoyl)-L-homoserine lactone has immunomodulatory activity. Infect. Immun. 1998; 66(1): 36-42. https://doi.org/10.1128/iai.66.L36-42.1998
43. Boontham P., Robins A., Chandran P., Pritchard D., Camara M., Williams P., et al. Significant immunomodulatory effects of Pseudomonas aeruginosa quorum-sensing signal molecules: possible link in human sepsis. Clin. Sci. (Lond.) 2008; 115(11): 343-51. https://doi.org/10.1042/cs20080018
44. Stoltz D.A., Ozer E.A., Ng C.J., Yu J.M., Reddy S.T., Lusis A.J., et al. Paraoxonase-2 deficiency enhances Pseudomonas aeruginosa quorum sensing in murine tracheal epithelia. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2007; 292(4): L852-60. https://doi.org/10.1152/ajplung.00370.2006
45. Bar-Rogovsky H., Hugenmatter A., Tawfik D.S. The evolution-ary origins of detoxifying enzymes: the mammalian serum paraoxonases (PONs) relate to bacterial homoserine lactonases. J. Biol. Chem. 2013; 288(33): 23914-27. https://doi.org/10.1074/jbc.m112.427922
46. Бухарин О.В., Перунова Н.Б. Микросимбиоценоз. Екатеринбург; 2014.
47. Евдокименко А.Ю., Досадина Э.Э., Эль Регистан Г.И., Белов А.А. Влияние алкилоксибензолов на ферментативную активность некоторых гидролаз при различных условиях. Успехи в химии и химической технологии. 2016; 30(9): 10-2.
48. Gordon J.I. Honor thy gut symbionts redux. Science. 2012; 336(6086): 1251-3. https://doi.org/10.1126/science.1224686
49. Бухарин О.В. Адаптивные стратегии взаимодействия возбудителя и хозяина при инфекции. Вестник Российской академии наук. 2018; 88(7): 637-43. https://doi.org/10.31857/S086958730000087-3
50. Тарасенко В.С., Фадеев С.Б., Бухарин О.В. Хирургическая инфекция мягких тканей (клинико-микробиологический аспект). Екатеринбург; 2015.
51. Poutahidis T., Kearney S.M., Levkovich T., Qi P., Varian B.J., Lakritz J.R., et al. Microbial symbionts accelerate wound healing via the neuropeptide hormone oxytocin. PLoS One. 2013; 8(10): e78898. https://doi.org/10.1371/journal.pone.0078898
52. Poutahidis T., Springer A., Levkovich T., Qi P., Varian B.J., Lakritz J.R., et al. Probiotic microbes sustain youthful serum testosterone levels and testicular size in aging mice. PLoS One. 2014; 9(1): e84877. https://doi.org/10.1371/journal.pone.0084877
53. Poutahidis T., Kleinewietfeld M., Smillie C., Levkovich T., Perrotta A., Bhela S., et al. Microbial reprogramming inhibits Western diet-associated obesity. PLoS One. 2013; 8(7): e68596. https://doi.org/10.1371/journal.pone.0068596
54. Lopez-Larrea C., eds. Self and Nonself. New York: Springer; 2012.
55. Wenren L.M., Sullivan N.L., Cardarelli L., Septer A.N., Gibbs K.A. Two independent pathways for self-recognition in proteus mirabilis are linked by type VI-dependent export. mBio. 2013; 4(4): e00374-13. https://doi.org/10.1128/mbio.00374-13
56. Shank A.E., Kolter R. New developments in microbial inter-species signaling. Curr Opin. Microbiol. 2009; 12(2): 205-14. https://doi.org/10.1016/j.mib.2009.01.003
Что такое гомеостаз? | Живая наука
Гомеостаз — это способность поддерживать относительно стабильное внутреннее состояние, которое сохраняется, несмотря на изменения во внешнем мире. Все живые организмы, от растений до щенков и людей, должны регулировать свою внутреннюю среду, чтобы обрабатывать энергию и в конечном итоге выжить. Например, если у вас резко подскакивает кровяное давление или резко падает температура тела, ваши системы органов могут с трудом выполнять свою работу и в конечном итоге выходить из строя.
Почему важен гомеостаз
Физиолог Уолтер Кэннон ввел термин «гомеостаз» в 1920-е годы, расширив предыдущие работы покойного физиолога Клода Бернара.В 1870-х годах Бернар описал, как сложные организмы должны поддерживать баланс в своей внутренней среде, или «внутренней среде , », , чтобы вести «свободную и независимую жизнь» в мире за ее пределами. Кэннон отточил эту концепцию и представил гомеостаз широкой аудитории в своей книге «Мудрость тела» (Британский медицинский журнал, 1932). [7 величайших тайн человеческого тела]
Провозглашенное основным принципом физиологии, основное определение гомеостаза, данное Кэнноном, используется и сегодня.Термин происходит от греческих корней, означающих «подобный» и «состояние стабильности». Приставка «гомео» подчеркивает, что гомеостаз не работает как термостат или круиз-контроль в автомобиле, фиксированный на одной точной температуре или скорости. Вместо этого, согласно обзору в журнале Appetite, гомеостаз удерживает важные физиологические факторы в пределах допустимого диапазона значений.
Человеческое тело, например, регулирует внутреннюю концентрацию водорода, кальция, калия и натрия, заряженных частиц, на которые клетки полагаются для нормального функционирования.Согласно обзору 2015 года, опубликованному в журнале Advances in Physiology Education, гомеостатические процессы также поддерживают уровни воды, кислорода, pH и сахара в крови, а также температуру тела.
Согласно Scientific American, в здоровых организмах гомеостатические процессы происходят постоянно и автоматически. Несколько систем часто работают в тандеме, чтобы поддерживать один физиологический фактор, например температуру тела. Если эти меры не сработают или потерпят неудачу, организм может умереть от болезни или даже смерти.
Как поддерживается гомеостаз
Многие гомеостатические системы прислушиваются к сигналам бедствия от тела, чтобы узнать, когда ключевые переменные выходят за пределы допустимого диапазона.Нервная система обнаруживает эти отклонения и сообщает в центр управления, часто расположенный в головном мозге. Затем центр управления направляет мышцы, органы и железы на исправление нарушения. Согласно онлайн-учебнику «Анатомия и физиология», непрерывный цикл нарушения и приспособления известен как «отрицательная обратная связь».
Например, человеческое тело поддерживает внутреннюю температуру около 98,6 градусов по Фаренгейту (37 градусов по Цельсию). При перегреве термодатчики на коже и в мозге издают сигнал тревоги, инициируя цепную реакцию, которая заставляет тело вспотеть и покраснеть.При охлаждении тело реагирует дрожью и сокращает кровообращение к коже. Точно так же, когда уровень натрия резко повышается, организм сигнализирует почкам, чтобы они сберегали воду и выводили излишки соли с концентрированной мочой.
Животные также изменят свое поведение в ответ на отрицательный отзыв. Например, при перегреве мы можем сбросить слой одежды, уйти в тень или выпить стакан холодной воды.
Современные модели гомеостаза
Концепция отрицательной обратной связи восходит к описанию гомеостаза Кэнноном в 1920-х годах и была первым объяснением того, как работает гомеостаз.Но в последние десятилетия многие ученые утверждают, что организмы способны предвидеть возможные нарушения гомеостаза, а не только реагировать на них постфактум.
Согласно статье 2015 года в Psychological Review, эта альтернативная модель гомеостаза, известная как аллостаз, подразумевает, что идеальная уставка для конкретной переменной может смещаться в ответ на временные изменения окружающей среды. Точка может смещаться под влиянием циркадных ритмов, менструальных циклов или суточных колебаний температуры тела.Согласно обзору 2015 года, опубликованному в журнале Advances in Physiology Education, уставки также могут изменяться в ответ на физиологические явления, такие как лихорадка, или для компенсации нескольких гомеостатических процессов, происходящих одновременно.
«Сами по себе заданные значения не являются фиксированными, но могут демонстрировать адаптивную пластичность», — сказал Арт Вудс, биолог из Университета Монтаны в Миссуле. «Эта модель позволяет упреждающе реагировать на предстоящие потенциальные нарушения в заданных точках».
Например, согласно обзору 2007 года в Appetite, в ожидании еды организм выделяет дополнительный инсулин, грелин и другие гормоны.Эта превентивная мера подготавливает организм к поступающему потоку калорий, а не пытается контролировать уровень сахара в крови и запасы энергии.
Возможность смещения заданных значений позволяет животным адаптироваться к краткосрочным стрессовым факторам, но они могут потерпеть неудачу перед лицом долгосрочных проблем, таких как изменение климата.
«Активация гомеостатических систем ответа может быть хорошей на короткие периоды времени», — сказал Вудс. Но они не рассчитаны на длительный срок службы. «Гомеостатические системы могут катастрофически выйти из строя, если их зайти слишком далеко; поэтому, хотя системы могут быть в состоянии справиться с краткосрочными новыми климатами, они могут быть не в состоянии справиться с большими изменениями в течение более длительных периодов времени.»
Гомеостатические точки могут быть адаптивными. Например, в ожидании еды организм выделяет дополнительный инсулин, грелин и другие гормоны, чтобы подготовить тело к поступающему потоку калорий, а не бороться за контроль над уровнем сахара в крови и запасами энергии. (Изображение предоставлено Shutterstock)Поддержание потока информации
Гомеостатические системы, возможно, в первую очередь развивались, чтобы помочь организмам поддерживать оптимальное функционирование в различных средах и ситуациях. Но, согласно эссе 2013 года в журнале Trends in Ecology & Evolution, некоторые ученые предполагают, что гомеостаз в первую очередь обеспечивает «тихий фон» для взаимодействия клеток, тканей и органов друг с другом.Теория утверждает, что гомеостаз облегчает организмам извлечение важной информации из окружающей среды и передачу сигналов между частями тела.
Независимо от своего эволюционного предназначения, гомеостаз уже почти столетие формирует исследования в области наук о жизни. Хотя гомеостатические процессы в основном обсуждаются в контексте физиологии животных, они также позволяют растениям управлять запасами энергии, питать клетки и реагировать на проблемы окружающей среды. Помимо биологии, социальные науки, кибернетика, информатика и инженерия используют гомеостаз в качестве основы для понимания того, как люди и машины поддерживают стабильность, несмотря на сбои.
Дополнительно r Источники:
- Ознакомьтесь с полезными графиками о гомеостазе из Академии Хана.
- Узнайте, как гомеостаз влияет на физиологию человека, с помощью ускоренного курса.
- Посмотрите это видео от сестер Амеба, чтобы узнать больше об отрицательных отзывах.
Гомеостаз | Анатомия и физиология
Введение в гомеостаз
Гомеостаз означает способность организма поддерживать стабильную внутреннюю среду (регулирующие гормоны, температура тела., водный баланс и др.). Поддержание гомеостаза требует, чтобы организм постоянно следил за своими внутренними условиями. От температуры тела до артериального давления и уровней определенных питательных веществ каждое физиологическое состояние имеет определенную уставку. Уставка — это физиологическое значение, вокруг которого колеблется нормальный диапазон. Нормальный диапазон — это ограниченный набор значений, который является оптимально здоровым и стабильным. Например, уставка нормальной температуры человеческого тела составляет примерно 37 ° C (98.6 ° F) Физиологические параметры, такие как температура тела и артериальное давление, имеют тенденцию колебаться в пределах нормы на несколько градусов выше и ниже этой точки. Центры управления в головном мозге играют роль в регулировании физиологических параметров и поддержании их в пределах нормы. Поскольку организм работает над поддержанием гомеостаза, любому значительному отклонению от нормального диапазона будет сопротивляться, и гомеостаз будет восстановлен посредством процесса, называемого петлей обратной связи .
Цепь обратной связи состоит из трех основных компонентов (рисунок 1.10а). Датчик , также известный как рецептор , является компонентом системы обратной связи, которая отслеживает физиологическое значение. Он отвечает за обнаружение изменений в окружающей среде. Это значение сообщается в центр управления. Центр управления — это компонент системы обратной связи, который сравнивает значение с нормальным диапазоном. Если значение слишком сильно отклоняется от уставки, центр управления активирует исполнительный элемент. Эффектор — это компонент в системе обратной связи, который вызывает изменение, обращающее ситуацию вспять и возвращающее значение в нормальный диапазон.Эффекторы — мышцы и железы.
Два типа петель обратной связи: отрицательная и положительная
Отрицательная обратная связь — это механизм, в котором эффект реакции на стимул заключается в отключении исходного стимула или уменьшении его интенсивности. Петли отрицательной обратной связи — наиболее распространенные механизмы организма, используемые для поддержания гомеостаза. Поддержание гомеостаза с помощью отрицательной обратной связи происходит во всем теле постоянно, и поэтому понимание отрицательной обратной связи является фундаментальным для понимания физиологии человека.
Рисунок 1.10. Цепь отрицательной обратной связи В петле отрицательной обратной связи стимулу — отклонению от заданного значения — сопротивляется физиологический процесс, который возвращает тело к гомеостазу. (а) Петля отрицательной обратной связи состоит из четырех основных частей. (б) Температура тела регулируется отрицательной обратной связью.Чтобы привести систему в движение, стимул изменяет внутреннюю среду за пределы ее нормального диапазона (то есть за пределы гомеостаза).Этот стимул улавливается определенным рецептором. Например, при контроле уровня глюкозы в крови определенные эндокринные клетки поджелудочной железы обнаруживают избыток глюкозы (стимул) в кровотоке. Эти бета-клетки поджелудочной железы реагируют на повышенный уровень глюкозы в крови, высвобождая гормон инсулин в кровоток. Инсулин сигнализирует волокнам скелетных мышц, жировым клеткам (адипоцитам) и клеткам печени, чтобы они поглощали избыток глюкозы, удаляя ее из кровотока. Когда концентрация глюкозы в кровотоке падает, снижение концентрации — фактическая отрицательная обратная связь — обнаруживается альфа-клетками поджелудочной железы, и высвобождение инсулина прекращается.Это предотвращает дальнейшее падение уровня сахара в крови ниже нормального диапазона.
У людей есть аналогичная система обратной связи по регулированию температуры, которая работает, способствуя либо потере тепла, либо притоку тепла (рис. 1.10b). Когда центр регуляции температуры мозга получает данные от датчиков, указывающие, что температура тела превышает нормальный диапазон, он стимулирует скопление клеток мозга, называемое «центром потери тепла». Эта стимуляция имеет три основных эффекта:
- Кровеносные сосуды кожи начинают расширяться, позволяя большему количеству крови из сердцевины тела течь к поверхности кожи, позволяя теплу излучаться в окружающую среду.
- Когда приток крови к коже увеличивается, потовые железы активируются, чтобы увеличить свою продукцию. Когда пот испаряется с поверхности кожи в окружающий воздух, он уносит с собой тепло.
- Глубина дыхания увеличивается, и человек может дышать через открытый рот, а не через носовые проходы. Это еще больше увеличивает потерю тепла из легких.
Напротив, активация центра накопления тепла в головном мозге воздействием холода снижает приток крови к коже, и кровь, возвращающаяся от конечностей, отводится в сеть глубоких вен.Такое расположение улавливает тепло ближе к сердцевине корпуса и ограничивает теплопотери. Если потеря тепла велика, мозг вызывает усиление случайных сигналов к скелетным мышцам, заставляя их сокращаться и вызывать дрожь. Сокращения мышц при дрожании высвобождают тепло при использовании АТФ. Мозг заставляет щитовидную железу в эндокринной системе выделять гормон щитовидной железы, который увеличивает метаболическую активность и производство тепла в клетках по всему телу. Мозг также сигнализирует надпочечникам о высвобождении адреналина (адреналина), гормона, который вызывает расщепление гликогена на глюкозу, которая может использоваться в качестве источника энергии.Распад гликогена на глюкозу также приводит к усилению метаболизма и выработке тепла.
Концентрация воды в организме имеет решающее значение для правильного функционирования. Тело человека сохраняет очень жесткий контроль над уровнем воды без сознательного контроля со стороны человека. Посмотрите это видео, чтобы узнать больше о концентрации воды в организме. Какой орган в первую очередь контролирует количество воды в организме?
Положительная обратная связь усиливает изменение физиологического состояния организма, а не обращает его вспять.Отклонение от нормального диапазона приводит к большим изменениям, и система уходит дальше от нормального диапазона. Положительная обратная связь в организме нормальна только тогда, когда есть определенная конечная точка. Роды и реакция организма на кровопотерю — два примера положительных петель обратной связи, которые нормальны, но активируются только при необходимости.
Доношенные роды — это пример ситуации, в которой сохранение существующего состояния организма нежелательно. Для изгнания ребенка в конце беременности требуются колоссальные изменения в организме матери.И события родов, однажды начавшись, должны быстро развиваться до завершения, иначе жизнь матери и ребенка окажется под угрозой. Чрезвычайная мускульная работа во время родов и родоразрешения является результатом системы положительной обратной связи (рис. 1.11).
Рисунок 1.11. Цепь положительной обратной связи Нормальные роды происходят благодаря положительной обратной связи. Положительная обратная связь приводит к изменению статуса тела, а не к возвращению к гомеостазу.Первые схватки (раздражитель) подталкивают ребенка к шейке матки (самой нижней части матки).Шейка матки содержит чувствительные к растяжению нервные клетки, которые контролируют степень растяжения (датчики). Эти нервные клетки отправляют сообщения в мозг, который, в свою очередь, заставляет гипофиз в основании мозга выделять гормон окситоцин в кровоток. Окситоцин вызывает более сильное сокращение гладких мышц матки (эффекторов), толкая ребенка дальше по родовым путям. Это вызывает еще большее растяжение шейки матки. Цикл растяжения, высвобождения окситоцина и все более сильных сокращений прекращается только после рождения ребенка.На этом этапе растяжение шейки матки прекращается, останавливая выброс окситоцина.
Второй пример положительной обратной связи сосредоточен на обращении вспять крайних повреждений тела. После проникающей раны наиболее непосредственной угрозой является чрезмерная кровопотеря. Меньшая циркуляция крови означает снижение артериального давления и уменьшение перфузии (проникновения крови) в мозг и другие жизненно важные органы. Если перфузия сильно снижена, жизненно важные органы отключатся, и человек умрет. Организм реагирует на эту потенциальную катастрофу, выделяя в поврежденную стенку кровеносного сосуда вещества, которые запускают процесс свертывания крови.По мере того, как происходит каждый этап свертывания, он стимулирует высвобождение большего количества свертывающихся веществ. Это ускоряет процессы свертывания и закрытия поврежденного участка. Свертывание ограничивается определенной областью, основанной на строго контролируемой доступности белков свертывания. Это адаптивный, спасающий жизнь каскад событий.
Интегрирующие системы
Каждая система органов выполняет определенные функции для организма, и каждая система органов обычно изучается независимо. Однако системы органов также работают вместе, чтобы помочь телу поддерживать гомеостаз.
Например, сердечно-сосудистая, мочевыделительная и лимфатическая системы помогают организму контролировать водный баланс. Сердечно-сосудистая и лимфатическая системы транспортируют жидкости по всему телу и помогают определять уровни растворенных веществ и воды, а также регулировать давление. Если уровень воды становится слишком высоким, мочевыделительная система производит более разбавленную мочу (мочу с более высоким содержанием воды), чтобы помочь устранить избыток воды. Если уровень воды становится слишком низким, образуется более концентрированная моча, чтобы сохранить воду.Пищеварительная система также играет роль с переменным водопоглощением. Вода может быть потеряна через покровную и дыхательную системы, но эта потеря не связана напрямую с поддержанием жидкости в организме и обычно связана с другими гомеостатическими механизмами.
Точно так же сердечно-сосудистая, покровная, дыхательная и мышечная системы работают вместе, чтобы помочь телу поддерживать стабильную внутреннюю температуру. Если температура тела повышается, кровеносные сосуды кожи расширяются, позволяя большему количеству крови течь к поверхности кожи.Это позволяет теплу рассеиваться через кожу в окружающий воздух. Кожа также может выделять пот, если тело становится слишком горячим; когда пот испаряется, это помогает охладить тело. Ускоренное дыхание также может помочь телу избавиться от избыточного тепла. В совокупности эти реакции на повышение температуры тела объясняют, почему вы потеете, тяжело дышите и краснетесь на лице, когда вы тренируетесь. (Тяжелое дыхание во время упражнений также является одним из способов, которым организм получает больше кислорода к вашим мышцам и избавляется от лишнего углекислого газа, производимого мышцами.)
И наоборот, если ваше тело слишком холодное, кровеносные сосуды кожи сокращаются, и кровоток к конечностям (рукам и ногам) замедляется. Мышцы быстро сокращаются и расслабляются, благодаря чему выделяется тепло, чтобы согреться. Волосы на коже поднимаются вверх, задерживая около кожи больше воздуха, который является хорошим изолятором. Эти реакции на снижение температуры тела объясняют, почему вы дрожите, получаете «мурашки по коже» и холодные, бледные конечности, когда вам холодно.
Щелкните эту ссылку и переместите ползунок, чтобы увидеть моделирование гомеостатического контроля температуры.Поддержание гомеостаза | Биология для майоров II
Результаты обучения
- Объясните, как разные системы органов связаны друг с другом для поддержания гомеостаза
Каждая система органов выполняет определенные функции для организма, и каждая система органов обычно изучается независимо. Однако системы органов также работают вместе, чтобы помочь телу поддерживать гомеостаз.
Уровни воды
Например, сердечно-сосудистая, мочевыделительная и лимфатическая системы помогают организму контролировать водный баланс.Сердечно-сосудистая и лимфатическая системы транспортируют жидкости по всему телу и помогают определять уровни растворенных веществ и воды, а также регулировать давление. Если уровень воды становится слишком высоким, мочевыделительная система производит более разбавленную мочу (мочу с более высоким содержанием воды), чтобы помочь устранить избыток воды. Если уровень воды становится слишком низким, образуется более концентрированная моча, чтобы сохранить воду.
Внутренние температуры
Точно так же сердечно-сосудистая, покровная (кожа и связанные с ней структуры), дыхательная и мышечная системы работают вместе, помогая телу поддерживать стабильную внутреннюю температуру.Если температура тела повышается, кровеносные сосуды кожи расширяются, позволяя большему количеству крови течь к поверхности кожи. Это позволяет теплу рассеиваться через кожу в окружающий воздух. Кожа также может выделять пот, если тело становится слишком горячим; когда пот испаряется, это помогает охладить тело. Ускоренное дыхание также может помочь телу избавиться от избыточного тепла. В совокупности эти реакции на повышение температуры тела объясняют, почему вы потеете, тяжело дышите и краснетесь на лице, когда вы тренируетесь.(Тяжелое дыхание во время упражнений также является одним из способов, которым организм получает больше кислорода к вашим мышцам и избавляется от лишнего углекислого газа, вырабатываемого мышцами.)
И наоборот, если ваше тело слишком холодное, кровеносные сосуды кожи сокращаются, и кровоток к конечностям (рукам и ногам) замедляется. Мышцы быстро сокращаются и расслабляются, благодаря чему выделяется тепло, чтобы согреться. Волосы на коже поднимаются вверх, задерживая около кожи больше воздуха, который является хорошим изолятором. Эти реакции на снижение температуры тела объясняют, почему вы дрожите, получаете «мурашки по коже» и холодные, бледные конечности, когда вам холодно.
Пример: лихорадка
Так что же происходит, когда у вас жар? Означает ли это, что ваше тело не может поддерживать свой гомеостаз, точно так же, как в вашем доме станет слишком жарко, если ваш кондиционер сломается?
В крайних случаях жар может потребовать неотложной медицинской помощи; но лихорадка — это адаптивный физиологический ответ нашего организма на определенные инфекционные агенты. Определенные химические вещества, называемые пирогенами, заставят ваш гипоталамус сместить заданное значение на более высокое значение. Это больше похоже на программирование термостата в вашем доме на более высокую температуру для экономии энергии в жаркий день, когда вы не собираетесь быть дома в течение дня.Эти пирогены могут поступать от заражающих вас микроорганизмов или вырабатываться клетками вашего тела в ответ на какую-либо инфекцию.
Практические вопросы
- По мере того, как уровень пирогенов в крови увеличивается, а заданное значение сбрасывается выше, хеморецепторы, которые теперь стимулируют гипоталамус, реагируют на ________ как переменную, а не терморецепторы, реагирующие на температуру тела как переменную.
- температура
- пирогены
- пульс
- артериальное давление
Вариант b верен.Увеличение количества пирогенных химических веществ в крови стимулирует рецепторы, которые сбрасывают верхний предел температуры для фебрильной реакции. Температура является переменной при нормальном регулировании температуры тела, но не в этом сценарии. В крови содержится химическое вещество, которое стимулирует фебрильную реакцию, но частота сердечных сокращений напрямую не стимулирует этот рецептор. В крови содержится химическое вещество, которое стимулирует фебрильную реакцию, но артериальное давление не стимулирует напрямую этот рецептор.
- Центр управления _________.
- скелетная мышца
- потовые железы
- сосуды
- гипоталамус
Ответ d правильный. Гипоталамус является центром контроля как гомеостаза нормальной температуры тела, так и фебрильной реакции. Скелетные мышцы, потовые железы и кровеносные сосуды — все это эффекторы.
- Поскольку уставка была увеличена, теперь вам холодно, даже если температура вашего тела обычно находится в пределах здорового диапазона.Это вызывает «озноб», который вы чувствуете при повышении температуры тела. В ответ гипоталус будет работать на повышение температуры тела. Какой ответ сделает это?
- Гипоталамус будет стимулировать потовые железы и расширять кровеносные сосуды в качестве эффекторов для охлаждения тела.
- Гипоталамус стимулирует скелетные мышцы дрожать и сужать кровеносные сосуды.
Вариант b верен. Это повысит температуру тела. Вариант А снизит температуру тела.
Хотя доказательства носят косвенный характер, считается, что лихорадка усиливает иммунный ответ организма. Повышенная температура может фактически нарушить репликацию инфекционных бактерий и вирусов, которые лучше всего приспособлены к выживанию в нормальном диапазоне гомеостатических температур тела. Это может дать вашим иммунным клеткам шанс уничтожить микроорганизмы, прежде чем они начнут быстро размножаться и распространяться в организме. Есть также некоторые косвенные доказательства того, что повышение температуры тела незначительно изменяет некоторые метаболические реакции, что также позволяет иммунной системе функционировать более эффективно.
Практические вопросы
- Когда достигается новое более высокое заданное значение, терморецепторы стимулируют _________ как центр управления.
- скелетная мышца
- потовые железы
- сосуды
- гипоталамус
Вариант d верен. Гипоталамус является центром контроля как гомеостаза нормальной температуры тела, так и фебрильной реакции. Мышцы, потовые железы и кровеносные сосуды являются эффекторами; они не служат центром управления.
- В ответ потовые железы и кровеносные сосуды (эффекторы) стимулируются на _________.
- выделяют пот для испарения и расширяют сосуды для увеличения потери тепла кровью вблизи поверхности кожи.
- дрожит, чтобы создать тепло, и сужает сосуды, чтобы сохранить тепло, удерживая кровь подальше от поверхности кожи.
Вариант а правильный. Это охладит тело. Вариант б согреет тело.
К сожалению, во время некоторых инфекций уровень пирогенов колеблется.”Это регулирует заданное значение температуры вверх и вниз. Когда уровень пирогена падает, вы получаете другую часть лихорадки: «пот» и чувство покраснения. Пока уровни пирогена продолжают увеличиваться и уменьшаться, вы будете чувствовать, как будто вы раскачиваетесь взад и вперед.
Практический вопрос
- После того, как уровень пирогена снизится из-за того, что инфекция находится под контролем, ________ (центр управления) сбросит более высокое заданное значение до нормального.
- терморецепторы
- хеморецепторов
- гипоталамус
Вариант c верен.Гипоталамус по-прежнему является центром управления, который реагирует на стимул от какого-либо рецептора. Терморецепторы и хеморецепторы стимулируют центр управления в ответ на изменение переменной, которую они контролируют, в данном случае температуры тела.
Ваше тело будет продолжать колебаться между верхним и нижним пределом температуры тела, но поскольку теперь оно находится в пределах вашего «нормального» диапазона температур, вы, вероятно, даже не заметите, что ваше тело все еще работает, поддерживая гомеостаз этой переменной.
Практический вопрос
- У пациентов после операции часто поднимается температура. Что из следующего, , а не , может быть разумной причиной такого ответа?
- Травма ткани в результате операции стимулировала клетки тела к высвобождению пирогенов.
- Несмотря на меры предосторожности, некоторые бактерии заразили человека во время операции.
- Операция повредила терморецепторы
- Послеоперационные лекарства повлияли на иммунную систему, вызвав выброс пирогенов.
Вариант c верен. Терморецепторы расположены по всему телу, поэтому маловероятно, что операция может напрямую повредить все рецепторы. Все остальные варианты могли стать причиной послеоперационной лихорадки.
Гомеостаз ионов
Функции организма, такие как регуляция сердцебиения, сокращение мышц, активация ферментов и клеточная связь, требуют строго регулируемого уровня кальция. Обычно мы получаем много кальция с пищей.Тонкий кишечник усваивает кальций из переваренной пищи.
Эндокринная система — это центр управления гомеостазом кальция в крови. Паращитовидные и щитовидные железы содержат рецепторы, которые реагируют на уровень кальция в крови. В этой системе обратной связи уровень кальция в крови является переменной величиной, потому что он изменяется в зависимости от окружающей среды. Изменения уровня кальция в крови имеют следующие эффекты:
- При низком уровне кальция в крови паращитовидная железа выделяет паратироидный гормон .Этот гормон заставляет эффекторные органы (почки и кости) реагировать на повышение уровня кальция. Почки препятствуют выведению кальция с мочой. Остеокласты в костях реабсорбируют костную ткань и выделяют кальций.
- Когда уровень кальция в крови высок, щитовидная железа выделяет кальцитонин . Кальцитонин заставляет почки реабсорбировать меньше кальция из фильтрата, что позволяет удалить избыток кальция из организма с мочой. Кальцитонин также подавляет образование активного витамина D в почках; без витамина D тонкий кишечник не усваивает так много кальция, поступающего с пищей.Остеобласты, стимулируемые кальцитонином, используют кальций в крови для добавления в костную ткань.
Практические вопросы
Основываясь на приведенном выше описании гомеостаза кальция, попробуйте ответить на следующие вопросы:
|
|
- Вариант d верен: кальций — переменная.Правильный уровень кальция важен для многих функций организма.
- Вариант b верен: эндокринная система является рецептором. Эндокринная система регулирует многое.
- Вариант б верен: эндокринная система — это центр управления. Эндокринная система может как определять, так и регулировать уровень кальция. Эффектором являются паратироидный гормон и кальцитонин.
- Вариант c правильный: паратироидный гормон и кальцитонин являются эффекторами; они изменяют функцию почек и костей для поддержания гомеостаза кальция.
Нарушение баланса кальция в крови может привести к болезни или даже смерти. Гипокальциемия означает низкий уровень кальция в крови. Признаки гипокальциемии включают мышечные спазмы и сердечные нарушения. Гиперкальциемия возникает, когда уровень кальция в крови выше нормы. Гиперкальциемия также может вызвать нарушение работы сердца, а также мышечную слабость и образование камней в почках.
Практический вопрос
Какие проблемы связаны с дисфункцией гомеостаза кальция?
- порок сердца
- Болезнь костей
- оба
- ни
Вариант c верен.Сердце часто страдает от сильных краткосрочных изменений кальция, а кости часто страдают небольшими долгосрочными изменениями кальция. Нарушение гомеостаза кальция также может влиять на функцию мышц и приводить к образованию камней в почках.
Посмотрите это видео, чтобы еще раз обсудить гомеостаз и системы органов:
Внесите свой вклад!
У вас была идея улучшить этот контент? Нам очень понравится ваш вклад.
Улучшить эту страницуПодробнее
10.7. Гомеостаз и обратная связь — Biology LibreTexts
Твердая на ходу
Это устройство выглядит простым, но оно управляет сложной системой, которая поддерживает постоянную температуру в доме. Устройство представляет собой устаревший термостат. Циферблат показывает текущую температуру в комнате, а также позволяет жителю установить термостат на желаемую температуру. Термостат — это часто упоминаемая модель того, как живые системы, включая человеческое тело, поддерживают устойчивое состояние, называемое гомеостазом.
Рисунок \ (\ PageIndex {1} \): Термостат для зданияЧто такое гомеостаз?
Гомеостаз — это состояние, при котором такая система, как человеческое тело, поддерживается в более или менее устойчивом состоянии. Работа клеток, тканей, органов и систем органов по всему телу состоит в том, чтобы поддерживать множество различных переменных в узких пределах, совместимых с жизнью. Поддержание стабильной внутренней среды требует постоянного мониторинга внутренней среды и постоянного внесения корректировок для поддержания баланса.
Уставка и нормальный диапазон
Для любой заданной переменной, такой как температура тела или уровень глюкозы в крови, существует конкретная уставка , которая является физиологическим оптимальным значением. Например, заданная температура человеческого тела составляет около 37 ºC (98,6 ºF). Поскольку организм работает над поддержанием гомеостаза температуры или любой другой внутренней переменной, значение обычно колеблется около заданного значения. Такие колебания нормальны до тех пор, пока они не становятся слишком сильными.Разброс значений, в пределах которого такие колебания считаются незначительными, называется нормальным диапазоном . В случае температуры тела, например, нормальный диапазон для взрослого составляет от 36,5 до 37,5 ºC (от 97,7 до 99,5 ºF).
Поддержание гомеостаза
Гомеостаз в организме человека обычно поддерживается чрезвычайно сложным уравновешивающим действием. Независимо от того, поддерживается ли переменная в пределах своего нормального диапазона, для поддержания гомеостаза требуется по крайней мере четыре взаимодействующих компонента: стимул, сенсор, центр управления и эффектор.
- Стимул обеспечивается регулируемой переменной. Как правило, стимул указывает на то, что значение переменной отклонилось от заданного значения или вышло за пределы нормального диапазона.
- Датчик отслеживает значения переменной и отправляет данные о них в центр управления.
- Центр управления сравнивает данные с нормальными значениями. Если значение не соответствует заданному значению или находится за пределами нормального диапазона, центр управления отправляет сигнал на исполнительный элемент.
- Эффектор — это орган, железа, мышца или другая структура, которая воздействует на сигнал из центра управления, чтобы переместить переменную обратно к заданному значению.
Каждый из этих компонентов показан на рисунке \ (\ PageIndex {2} \). Диаграмма слева — это общая модель, показывающая, как компоненты взаимодействуют для поддержания гомеостаза. Стимул активирует сенсор. Датчик активирует систему управления, регулирующую эффектор. На диаграмме справа показан пример температуры тела.Из диаграмм вы можете видеть, что поддержание гомеостаза включает в себя обратную связь, то есть данные, которые используются для управления ответной реакцией. Высокая температура тела может стимулировать центр регулирования температуры мозга, чтобы активировать потовые железы, чтобы снизить температуру тела. Когда температура тела достигает нормального диапазона, это действует как отрицательная обратная связь, чтобы остановить процесс. Отзыв может быть как положительным, так и отрицательным. Все механизмы обратной связи, поддерживающие гомеостаз, используют отрицательную обратную связь. Биологические примеры положительной обратной связи встречаются гораздо реже.
Рисунок \ (\ PageIndex {2} \): Для поддержания гомеостаза посредством обратной связи требуется стимул, датчик, центр управления и эффектор.Отрицательная обратная связь
В контуре отрицательной обратной связи обратная связь служит для уменьшения чрезмерного отклика и удержания переменной в нормальном диапазоне. Примеры процессов, контролируемых отрицательной обратной связью, включают регулирование температуры тела и контроль уровня глюкозы в крови.
Температура тела
Регулирование температуры тела включает отрицательную обратную связь, снижает ли она температуру или повышает ее (Рисунок \ (\ PageIndex {3} \)).
Охлаждение
Центром регулирования температуры человеческого тела является гипоталамус головного мозга. Когда гипоталамус получает данные от датчиков в коже и головном мозге о том, что температура тела выше заданного значения, он приводит в действие следующие реакции:
- Кровеносные сосуды в коже расширяются (вазодилатация), позволяя большему количеству крови из теплого ядра тела течь близко к поверхности тела, поэтому тепло может излучаться в окружающую среду.
- Когда приток крови к коже увеличивается, потовые железы в коже активируются, чтобы увеличить выделение пота (потоотделение).Когда пот испаряется с поверхности кожи в окружающий воздух, он уносит с собой тепло.
- Дыхание становится глубже, и человек может дышать через рот, а не через носовые проходы. Это увеличивает потерю тепла из легких.
Нагревание
Когда центр регулирования температуры мозга получает данные о том, что температура тела ниже заданного значения, он приводит в действие следующие реакции:
- Кровеносные сосуды кожи сужаются (сужение сосудов), чтобы кровь не могла течь близко к поверхности тела.Это снижает потери тепла с поверхности.
- Когда температура падает, скелетным мышцам подаются случайные сигналы, заставляющие их сокращаться. Это вызывает дрожь, которая выделяет небольшое количество тепла.
- Мозг может стимулировать щитовидную железу (через гипофиз) на выработку большего количества гормонов щитовидной железы. Этот гормон увеличивает метаболическую активность и выработку тепла в клетках по всему телу.
- Надпочечники также могут быть стимулированы к секреции гормона адреналина.Этот гормон вызывает расщепление гликогена (углевода, используемого для хранения энергии у животных) до глюкозы, которая может использоваться в качестве источника энергии. Этот катаболический химический процесс является экзотермическим или выделяющим тепло.
Глюкоза крови
В процессе контроля уровня глюкозы в крови определенные эндокринные клетки поджелудочной железы, называемые альфа- и бета-клетками, определяют уровень глюкозы в крови. Затем они реагируют соответствующим образом, чтобы поддерживать уровень глюкозы в крови в пределах нормы.
- Если уровень глюкозы в крови поднимается выше нормы, бета-клетки поджелудочной железы выделяют гормон инсулин в кровоток. Инсулин дает клеткам сигнал забрать избыток глюкозы из крови до тех пор, пока уровень глюкозы в крови не снизится до нормального диапазона.
- Если уровень глюкозы в крови падает ниже нормы, альфа-клетки поджелудочной железы выделяют гормон глюкагон в кровоток. Глюкагон дает клеткам сигнал расщеплять накопленный гликоген до глюкозы и высвобождать глюкозу в кровь до тех пор, пока уровень глюкозы в крови не повысится до нормального диапазона.
Положительный отзыв
В контуре положительной обратной связи обратная связь служит для усиления ответа до тех пор, пока не будет достигнута конечная точка. Примеры процессов, контролируемых положительной обратной связью в организме человека, включают свертывание крови и роды.
Свертывание крови
Когда рана вызывает кровотечение, организм реагирует положительной петлей обратной связи, чтобы свернуть кровь и остановить кровопотерю. Вещества, выделяемые поврежденной стенкой кровеносного сосуда, запускают процесс свертывания крови.Тромбоциты в крови начинают цепляться за поврежденный участок и выделять химические вещества, которые привлекают дополнительные тромбоциты. По мере того, как тромбоциты продолжают накапливаться, выделяется больше химикатов и больше тромбоцитов привлекается к месту образования сгустка. Положительная обратная связь ускоряет процесс свертывания до тех пор, пока сгусток не станет достаточно большим, чтобы остановить кровотечение.
Рисунок \ (\ PageIndex {4} \): Нормальные роды обусловлены положительной обратной связью. Положительная обратная связь вызывает возрастающее отклонение от нормального состояния к фиксированной конечной точке, а не возврат к нормальному заданному значению, как при гомеостазеРоды
На рисунке \ (\ PageIndex {4} \) показан цикл положительной обратной связи, который контролирует роды.Обычно этот процесс начинается, когда голова младенца прижимается к шейке матки. Это стимулирует нервные импульсы, которые проходят от шейки матки к гипоталамусу головного мозга. В ответ гипоталамус отправляет гормон окситоцин в гипофиз, который выделяет его в кровоток, чтобы он мог попасть в матку. Окситоцин стимулирует сокращения матки, которые сильнее прижимают ребенка к шейке матки. В ответ шейка матки начинает расширяться, готовясь к выходу ребенка.Этот цикл положительной обратной связи продолжается с повышением уровня окситоцина, более сильными сокращениями матки и более широким расширением шейки матки, пока ребенок не будет вытолкнут через родовые пути и выйдет из тела. В этот момент шейка матки больше не стимулируется, чтобы посылать нервные импульсы в мозг, и весь процесс останавливается.
Когда гомеостаз не работает
Гомеостатические механизмы работают непрерывно, чтобы поддерживать стабильные условия в организме человека. Однако иногда механизмы выходят из строя.Когда они это сделают, это может привести к гомеостатическому дисбалансу , в котором клетки могут не получить все, что им нужно, или токсичные отходы могут накапливаться в организме. Если гомеостаз не восстанавливается, дисбаланс может привести к болезни или даже смерти. Диабет — это пример заболевания, вызванного дисбалансом гомеостаза. В случае диабета уровень глюкозы в крови больше не регулируется и может быть опасно высоким. Медицинское вмешательство может помочь восстановить гомеостаз и, возможно, предотвратить необратимое повреждение организма.
Характеристика: Мое человеческое тело
Диабет диагностируется у людей, у которых наблюдается аномально высокий уровень глюкозы в крови после голодания в течение не менее 12 часов. Уровень глюкозы в крови натощак ниже 100 — это нормально. Уровень от 100 до 125 относит вас к категории предиабета, а уровень выше 125 указывает на диагноз диабета.
Из двух типов диабета наиболее распространенным является диабет 2 типа, на который приходится около 90 процентов всех случаев диабета в Соединенных Штатах.Диабет 2 типа обычно начинается после 40 лет. Однако из-за резкого роста ожирения в последние десятилетия среди молодых людей возраст, в котором диагностируется диабет 2 типа, снизился. Даже детям сейчас ставят диагноз сахарный диабет 2 типа. Сегодня около 30 миллионов американцев страдают диабетом 2 типа, а еще 90 миллионов — предиабетом.
Скорее всего, вам нужно будет проверить уровень глюкозы в крови во время обычного медицинского осмотра. Если уровень глюкозы в крови указывает на то, что у вас диабет, это может стать для вас шоком, потому что у вас может не быть никаких симптомов заболевания.Вы не одиноки, потому что каждый четвертый диабетик не знает, что у них есть болезнь. Как только диагноз диабета утвердится, новости могут вас расстроить. Диабет может привести к сердечным приступам, инсультам, слепоте, почечной недостаточности и потере пальцев ног или стоп. Риск смерти у взрослых с диабетом на 50 процентов выше, чем у взрослых без диабета, и диабет является седьмой по значимости причиной смерти среди взрослых. Кроме того, контроль диабета обычно требует частого измерения уровня глюкозы в крови, наблюдения за тем, что и когда вы едите, приема лекарств или даже инъекций инсулина.Все это может показаться ошеломляющим.
Хорошая новость в том, что изменение образа жизни может остановить прогрессирование диабета 2 типа или даже обратить его вспять. Вот как:
- Похудеть. Любое похудение приносит пользу. Потеря всего лишь семи процентов вашего веса может быть всем, что нужно, чтобы остановить диабет на его пути. Особенно важно избавиться от лишнего веса в области талии.
- Регулярно выполняйте физические упражнения. Вы должны стараться заниматься спортом пять дней в неделю не менее 30 минут.Это не только снизит уровень сахара в крови, но и улучшит работу инсулина; это также снизит ваше кровяное давление и улучшит здоровье сердца. Еще одним преимуществом упражнений является то, что они помогут вам похудеть за счет увеличения скорости основного обмена.
- Примите здоровую диету. Уменьшите потребление рафинированных углеводов, таких как сладости и сладкие напитки. Увеличьте потребление продуктов, богатых клетчаткой, таких как фрукты, овощи и цельнозерновые продукты. Примерно четверть каждого приема пищи должна состоять из продуктов с высоким содержанием белка, таких как рыба, курица, молочные продукты, бобовые или орехи.
- Контрольный стресс. Стресс может повысить уровень глюкозы в крови, а также повысить кровяное давление и риск сердечных заболеваний. Когда вы чувствуете стресс, делайте дыхательные упражнения, совершайте быструю прогулку или бег трусцой. Кроме того, постарайтесь заменить стрессовые мысли более успокаивающими.
- Создать систему поддержки. Заручитесь помощью и поддержкой близких, а также медицинских работников, таких как диетолог и инструктор по диабету. Наличие системы поддержки поможет убедиться, что вы на пути к хорошему самочувствию и сможете придерживаться своего плана.
Обзор
- Что такое гомеостаз?
- Определите уставку и нормальный диапазон для физиологических измерений.
- Идентифицировать и определить четыре взаимодействующих компонента, которые поддерживают гомеостаз в петлях обратной связи.
- Сравните и сопоставьте контуры отрицательной и положительной обратной связи.
- Объясните, как отрицательная обратная связь влияет на температуру тела.
- Приведите два примера физиологических процессов, которые контролируются контурами положительной обратной связи.
- Петля отрицательной обратной связи:
- возвращает уровень переменной в нормальный диапазон
- может понижать, но не повышать температуру тела
- — это тип обратной связи, участвующий в свертывании крови
- A и B
- Во время кормления грудью ребенок начинает сосать сосок, и он увеличивает количество молока, производимого матерью. Чем больше сосет, тем обычно вырабатывается больше молока.
- Это пример отрицательной или положительной обратной связи? Поясните свой ответ.
- В чем, по вашему мнению, может быть эволюционная выгода от механизма регулирования производства молока, описанного в части а?
- Объясните, почему гомеостаз регулируется петлями отрицательной обратной связи, а не петлями положительной обратной связи.
- Уставка обычно:
- верхняя граница нормального диапазона
- нижняя граница нормального диапазона
- в середине нормального диапазона
- точка, в которой изменения больше не могут происходить
- Уровень полового гормона тестостерона (Т) контролируется отрицательной обратной связью.Другой гормон, гонадотропин-рилизинг-гормон (ГнРГ), выделяется гипоталамусом головного мозга, который заставляет гипофиз высвобождать лютеинизирующий гормон (ЛГ). ЛГ стимулирует гонады к выработке Т. Когда в кровотоке слишком много Т, он возвращается к гипоталамусу, заставляя его производить меньше ГнРГ. Хотя это не описывает все петли обратной связи, участвующие в регулировании T, ответьте на следующие вопросы об этой конкретной петле обратной связи.
- Что является стимулом в этой системе? Поясните свой ответ.
- Что такое центр управления в этой системе? Поясните свой ответ.
- Что считается в этой системе гипофизом: стимулом, сенсором, центром управления или эффектором? Поясните свой ответ.
Что такое гомеостаз?
Что такое гомеостаз?
Гомеостаз относится к потребности организма в достижении и поддержании определенного состояния равновесия. Этот термин впервые был придуман психологом Уолтером Кэнноном в 1926 году.В частности, гомеостаз — это тенденция организма контролировать и поддерживать внутренние состояния, такие как температура и уровень сахара в крови, на довольно постоянных и стабильных уровнях.
Гомеостаз означает способность организма регулировать различные физиологические процессы, чтобы поддерживать внутреннее состояние стабильным и сбалансированным. Эти процессы происходят в основном без нашего осознания.
Как обслуживается?
У вашего тела есть определенные точки для различных состояний, включая температуру, вес, сон, жажду и голод.Когда уровень отключен (в любом направлении, слишком много или слишком мало), гомеостаз исправит его. Например, чтобы регулировать температуру, вы будете потеть, когда вам станет слишком жарко, или дрожите, когда вам станет слишком холодно.
Одна известная теория мотивации человека, известная как теория уменьшения влечения, предполагает, что гомеостатический дисбаланс создает потребности. Эти потребности в восстановлении баланса побуждают людей выполнять действия, которые вернут тело в его идеальное состояние.
Другой способ думать об этом — это как о термостате в вашем доме.После установки в определенный момент он работает, чтобы поддерживать внутреннее состояние на этом уровне. Когда температура в вашем доме упадет, ваша печь включится и нагреет вещи до заданной температуры.
Точно так же, если что-то не сбалансировано в вашем теле, физиологическая реакция будет активирована до тех пор, пока уставка не будет снова достигнута. Вот как работают основные компоненты гомеостаза:
- Стимул : Стимул от изменения окружающей среды выбивает что-то из равновесия в теле.
- Приемник : Приемник реагирует на изменение, сообщая блоку управления.
- Блок управления : Затем блок управления сообщает об изменении, необходимом для восстановления баланса тела.
- Эффектор : Эффектор получает эту информацию и выполняет необходимые изменения.
Петля отрицательной обратной связи будет работать, чтобы уменьшить эффект стимула, а петля положительной обратной связи — увеличить его. В гомеостазе наиболее распространены петли отрицательной обратной связи, поскольку организм обычно пытается уменьшить эффект стимула, чтобы вернуть тело в равновесие.
Типы гомеостатической регуляции
В организме происходит три основных типа гомеостатической регуляции. Хотя их имена могут быть вам незнакомы, вы, вероятно, сталкиваетесь с ними каждый день.
Терморегуляция
Когда вы думаете о гомеостазе, в первую очередь на ум приходит температура. Это одна из самых важных и очевидных гомеостатических систем. Регулирование температуры тела называется терморегуляцией.
Все организмы, от крупных млекопитающих до крошечных бактерий, должны поддерживать идеальную температуру, чтобы выжить.Некоторые факторы, влияющие на эту способность поддерживать стабильную температуру тела, включают в себя то, как эти системы регулируются, а также общий размер организма.
- Эндотермы : Некоторые существа, известные как эндотермы или «теплокровные» животные, достигают этого посредством внутренних физиологических процессов. Птицы и млекопитающие (включая человека) — эндотермы.
- Ectotherms : Другие существа — эктотермы (также известные как «хладнокровные») и полагаются на внешние источники для регулирования температуры своего тела.И рептилии, и земноводные — эктотермы.
Разговорные термины «теплокровный» и «хладнокровный» на самом деле не означают, что у этих организмов разная температура крови. Эти термины просто относятся к , как эти существа поддерживают внутреннюю температуру тела.
На терморегуляцию также влияет размер организма или, более конкретно, отношение поверхности к объему.
- Крупные организмы : Более крупные существа имеют гораздо больший объем тела, что заставляет их выделять больше тепла.
- Мелкие организмы : С другой стороны, более мелкие животные производят меньше тепла, но также имеют более высокое отношение поверхности к объему. Они теряют больше тепла, чем производят, поэтому их внутренние системы должны работать намного усерднее, чтобы поддерживать постоянную температуру тела. Это касается даже младенцев, особенно недоношенных.
Осморегуляция
Осморегуляция стремится поддерживать необходимое количество воды и электролитов внутри и вне клеток организма.Баланс соли и воды на мембранах играет важную роль, как и в случае осмоса, отсюда и название «осморегуляция». В этом процессе почки несут ответственность за избавление от лишней жидкости, шлаков или электролитов. Осморегуляция также влияет на артериальное давление.
Химический регламент
Ваше тело регулирует и другие химические механизмы, чтобы поддерживать баланс систем. Они используют гормоны в качестве химических сигналов, например, в случае уровня сахара в крови.В этой ситуации поджелудочная железа будет выделять либо инсулин, когда уровень сахара в крови высокий, либо глюкагон, когда уровень сахара в крови низкий, для поддержания гомеостаза.
Влияние гомеостаза
Гомеостаз включает в себя как физиологические, так и поведенческие реакции. Что касается поведения, вы можете поискать теплую одежду или участок солнечного света, если вам станет холодно. Вы также можете согнуть свое тело внутрь и держать руки близко к телу, чтобы сохранить тепло.
Как эндотермы, у людей также есть ряд внутренних систем, которые помогают регулировать температуру тела.Когда температура вашего тела опускается ниже нормы, возникает ряд физиологических реакций, которые помогают восстановить равновесие. Кровеносные сосуды конечностей тела сужаются, чтобы предотвратить потерю тепла. Дрожь также помогает телу вырабатывать больше тепла.
Организм также реагирует, когда температура поднимается выше нормы. Вы когда-нибудь замечали, как ваша кожа краснеет, когда вам очень тепло? Это ваше тело пытается восстановить температурный баланс. Когда вам слишком тепло, ваши кровеносные сосуды расширяются, чтобы отдавать больше тепла телу.Потоотделение — еще один распространенный способ уменьшить тепло тела, поэтому в очень жаркий день вы часто покраснетесь и потеете.
Поддержание гомеостаза путем принятия решений
Abstract
Живым организмам необходимо поддерживать энергетический гомеостаз. Для многих видов это подразумевает действия с отсроченными последствиями. Например, людям, возможно, придется выбирать между поиском высококалорийной, но труднодоступной пищи и низкокалорийной, но легкодоступной пищи под угрозой голода.Гомеостатические принципы предписывают решения, которые максимизируют вероятность поддержания соответствующих уровней энергии на всей траектории поиска пищи. Здесь предсказания, основанные на биологических принципах, контрастируют с предсказаниями моделей принятия экономических решений, основанных на максимизации полезности конечного результата выбора. Для эмпирического арбитража между предсказаниями биологических и экономических моделей для принятия решений отдельными людьми мы разработали виртуальную задачу по добыче пищи, в которой игроки неоднократно выбирали между двумя средами кормодобывания, теряли энергию с течением времени и получали энергию вероятностно в соответствии со статистикой. среды, которую они выбрали.Достижение нулевой энергии было оформлено как голод. Мы использовали математику случайных блужданий, чтобы получить конечные распределения результатов выборов. Это также обеспечивало эквивалентные лотереи, представленные в чисто экономической, подобной казино, рамке, в которой голод соответствовал ничему выигрышу. Сравнение байесовских моделей показало, что как в рамках поиска пищи, так и в рамках казино выбор участников зависел совместно от вероятности голода и ожидаемой конечной ценности результата, но не мог быть объяснен экономическими моделями, основанными на комбинациях статистических моментов или полезность, зависящая от ранга.Это означает, что при точно определенных ограничениях биологические принципы лучше подходят для объяснения принятия решений человеком, чем экономические модели, основанные на максимизации конечной полезности.
Сведения об авторе
Общие модели принятия решений возникают на прочных аксиоматических основах, но не учитывают различные эмпирически наблюдаемые модели выбора, такие как отношение к риску перед лицом серьезных событий. Здесь мы утверждаем, что одна из причин такого несоответствия между теорией и данными заключается в пренебрежении основными биологическими принципами, такими как метаболический гомеостаз.Мы используем сравнение байесовских моделей, чтобы показать, что модели, основанные на гомеостатических соображениях, лучше объясняют человеческие решения, чем классические экономические модели — как в новой задаче виртуального поиска пищи, так и в стандартных экономических играх. В частности, мы показываем, что в соответствии с принципом гомеостаза человеческий выбор сводит к минимуму вероятность достижения нижней границы. Наши результаты подчеркивают, что предсказания, основанные на биологических принципах, обеспечивают простые, проверяемые и экологически рациональные объяснения очевидных предубеждений при принятии решений.
Образец цитирования: Korn CW, Bach DR (2015) Поддержание гомеостаза посредством принятия решений. PLoS Comput Biol 11 (5): e1004301. https://doi.org/10.1371/journal.pcbi.1004301
Редактор: Джилл Х. О’Рейли, Оксфордский университет, ВЕЛИКОБРИТАНИЯ
Поступила: 20 января 2015 г .; Принята к печати: 27 апреля 2015 г .; Опубликовано: 29 мая 2015 г.
Авторские права: © 2015 Korn, Bach. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника
Доступность данных: Все соответствующие данные находятся в пределах документ и вспомогательные информационные файлы к нему.Набор данных S1 содержит данные о поведении всех субъектов.
Финансирование: Эта работа финансировалась Цюрихским университетом. Центр Wellcome Trust для нейровизуализации поддерживается за счет основного финансирования Wellcome Trust [091593 / Z / 10 / Z]. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Гомеостаз имеет первостепенное значение для всех живых организмов [1].Проще говоря, организмы должны поддерживать свою внутреннюю среду в определенных границах, чтобы избежать смерти. Этот принцип гомеостаза отражается на уровнях молекулярных взаимодействий [2], петель гормональной обратной связи [3,4], нейронных цепей [5] и психофизиологических процессов [6]. Помимо необходимости немедленной регуляции, многие виды сталкиваются со сложными решениями с отсроченными и вероятными последствиями для долгосрочного метаболического гомеостаза. Здесь мы выдвигаем гипотезу о том, что гомеостатические требования определяют решения людей о кормлении.Например, охота на оленя дает большой прирост энергии с низкой вероятностью ее получения, а сбор ягод дает небольшой прирост энергии с высокой вероятностью. Чтобы свести к минимуму вероятность голода, агенты-люди должны интегрировать статистику доступных вариантов с их текущими уровнями энергии и их временным горизонтом.
Классические представления о гомеостазе [1–7] часто иллюстрируются термостатом, который определяет разницу между заданным значением температуры и текущей температурой.Это значение отклонения вызывает изменение уровней нагрева. Таким образом, термостат должен ретроспективно компенсировать уже проявившиеся отклонения. Напротив, мы предлагаем, чтобы лица, принимающие решения, могли предвидеть возможные отклонения и заранее минимизировать вероятность достижения запретительной границы, такой как голод. Это расширяет устоявшиеся представления о гомеостазе в (психо) физиологии [1–7] и согласуется с недавними теоретическими представлениями о гомеостазе как принципе, объясняющем принятие решений в здоровых и психиатрических популяциях [8–10].
В применении к индивидуальному принятию решений прогнозы этой модели противоречат экономическим моделям, которые прочно опираются на аксиоматические основы [11] и изящно объясняют многие типы денежных решений [12]. Эти модели постулируют, что лица, принимающие решения, основывают свой выбор на полезности, присвоенной конечному результату выбора, независимо от траектории к этой конечной точке [12–14].
В моделях риск-доходность и их вариантах [13,15] конечная полезность вычисляется с помощью статистических моментов распределения результатов, обычно ожидаемого значения, дисперсии, а в некоторых моделях также асимметрии.В обширной литературе есть обобщенные модели риска и доходности, которые включают субъективные преобразования статистических моментов [15,16]. В поведенческой экономике варианты моделей риска-доходности широко используются для описания того, как люди выбирают между денежными играми [17–19], как они оценивают события реальной жизни [16] и как животные выбирают основные подкрепления [20,21]. ]. Теория ожидаемой полезности и ее производные составляют еще один класс моделей, в которых значения возможных результатов трансформируются во внутреннюю меру полезности с помощью индивидуальной функции полезности лица, принимающего решения [12,15,22].Зависимые от ранга полезные модели дополнительно дополняют нелинейное взвешивание вероятностей исхода опциона [23,24]. Подобно моделям риск-доходность, полезные модели, зависящие от ранга, широко использовались для описания эмпирических данных как в лаборатории, так и на местах [15,23,24].
Эмпирически наблюдаемые отклонения от прогнозов этих микроэкономических моделей часто оформляются как иррациональные предубеждения, и дополнительные параметры включаются для поглощения таких предубеждений, но часто без принципиальных предположений о том, почему эти влияния должны возникать в первую очередь [25].Здесь мы представляем принципиальную биологическую причину отклонений от экономических принципов. Важно отметить, что максимизация полезности конечного результата данного набора вариантов игнорирует катастрофические последствия, если траектория этого результата достигает нижней границы внутренней среды. Мы стремились показать, что даже в безопасной лабораторной среде модель принятия решений, основанная на принципах гомеостаза, может объяснить решения о кормлении лучше, чем экономические модели. Кроме того, мы выдвинули гипотезу, что соображения гомеостаза также будут определять принятие человеческими решениями для просто структурированных лотерей без какой-либо ссылки на добычу пищи, как это часто используется в поведенческой экономике [12–14,26].
Чтобы проверить эти гипотезы, мы разработали виртуальную задачу по поиску пищи. В каждом испытании участники-люди выбирали одну из двух «кормовых сред» с различным возможным приростом «энергии» и соответствующими вероятностями, в которых они будут кормиться до трех последовательных «дней» (см. Рис. 1A для иллюстрации). В момент выбора полоса энергии отображала текущее внутреннее состояние участников. Участников проинструктировали, что каждый день сбора пищи они теряют одно очко энергии и получают энергию в соответствии со статистикой выбранной среды.Последовательные дни выполнения задания требовали интеграции рисков от нескольких попыток поиска пищи. Для каждого испытания участники принимали одно решение между двумя средами кормодобывания в течение указанного количества дней. Потеря всех энергетических очков в любой день определенного периода кормодобывания была оформлена как «голодная смерть», но не наказывалась явно. Каждое испытание было независимым. Мы не давали отзывов об исходах их выбора или промежуточных состояниях в последовательности поиска пищи. В конце эксперимента участники были вознаграждены за конечный результат поиска пищи в двух случайно выбранных испытаниях.Голод означал, что участники ничего не выиграли от испытания. Мы предположили, что участники будут вычислять вероятность голодной смерти для кормодобывающей среды и основывать свои решения на этой метрике.
Рис. 1. Виртуальная задача по поиску пищи и вывод азартных игр.
(A) Рамка для сбора корма: В каждом испытании участники видели энергетическую шкалу, отображающую их начальные энергетические точки. Участникам приходилось выбирать между двумя вариантами кормодобывания, которые были изображены в виде круговых диаграмм с двумя секторами: голубые секторы соответствовали неудачному поиску пищи (т.е., выигрышу в ноль баллов), синие секторы соответствовали успешному поиску пищи (т. е. увеличению количества очков, написанных над секторами). В каждом испытании участники принимали единственное решение, и выбранный вариант поиска пищи разыгрывался в течение указанного количества «дней». Для каждого дня определялся вероятностный результат выбранного варианта кормодобывания, и соответствующие приросты добавлялись к шкале энергии. Точная стоимость одного балла вычиталась каждый день, чтобы отразить потребление энергии.Если в какой-либо день полоса энергии достигла нуля, участник в этом испытании умирал от голода. Обозначим эту вероятность как p голодать . Таким образом, выбор в системе сбора пищи был между двумя последовательностями азартных игр, где количество дней указывало на количество игр в каждой последовательности. Участники не видели результатов своего выбора, но им были представлены примеры в письменных инструкциях. (B) Рамка казино: Численно идентичные азартные игры, как в рамке сбора пищи, были представлены в виде круговых диаграмм, подобных азартным играм с вращением колеса в казино.Размер каждого сектора обозначал вероятность выигрыша суммы, написанной рядом с ним. В кадре казино p starve был непосредственно виден как размер сектора при нулевом исходе. Таким образом, выбор в рамках казино был между двумя одношаговыми играми. (C) Иллюстрация логики, лежащей в основе математики случайных блужданий, которую мы использовали для вывода азартных игр (математические подробности см. В тексте S1). Древовидная иллюстрация изображает случайное блуждание, используемое для вычисления переменных правильных азартных игр в A и B.Случайное блуждание начинается с позиции «2», что соответствует начальному количеству энергетических баллов. На каждом шаге («день») агент идет вправо с вероятностью p (соответствует успешному поиску пищи; темно-синий сектор круговой диаграммы в A) или влево с вероятностью 1-p (соответствует неудачному поиску пищи. ; голубой сектор круговой диаграммы в A). Размер шага «влево» соответствует расчетной стоимости и всегда равен единице. Размер шага вправо зависит от количества очков энергии, которое нужно получить (здесь +3 очка, и, следовательно, размер шага равен +2, потому что точная стоимость должна быть вычтена).Ноль — это поглощающая граница, поэтому стрелки не начинаются с нуля. Возможные результаты видны непосредственно в рамке казино (здесь 0, 2, 5 и 8 баллов). Чтобы определить вероятность определенного исхода, нужно проследить все возможные пути вдоль ветвей дерева, ведущих к этому результату, и просуммировать их вероятности. Вероятности конкретного «пути по ветвям» определяются путем умножения вероятностей всех стрелок на этом пути. В текущем примере вероятность результата равна нулю (т.е.е., p голодать ) составляет q 2 , а вероятность исхода 2 равна 2pq 2 . (D) Распределение переменных для случайного блуждания, показанного на C, по всем значениям p (т. Е. Вероятность успешного поиска пищи или правильного движения в случайном блуждании). В примере правильной игры, показанной на A и B, p было выбрано равным 40%, а пересечение черных вертикальных линий с зелеными линиями дает переменные для текущего примера (т. Е. Для начального уровня энергии в 2 балла энергии ).
https://doi.org/10.1371/journal.pcbi.1004301.g001
Мы использовали математику случайных блужданий для аналитического получения распределений конечных результатов периода кормодобывания и вероятностей голода во время кормодобывания (см. Рис. 1C для графической иллюстрации, Рис. 1D для примера переменных, полученных с помощью этой процедуры, Таблица 1 для обзора азартных игр и Методы и текст S1 для математических деталей). Поскольку участники были вознаграждены только за конечные результаты, гомеостатические принципы не имеют отношения к максимальному увеличению полезности, но наша задача предполагала их использование.Следовательно, мы проверили, влияют ли такие принципы на решения, если они не задействованы рамкой задачи. Мы представили участникам чисто экономические азартные игры, оформленные как лотереи казино с вращающимся колесом, без какого-либо упоминания о собирательстве (см. Рис. 1B). Эти лотереи имели те же конечные результаты, что и среда сбора пищи. Вероятность голодной смерти в фрейме сбора пищи соответствовала вероятности ничего не выиграть в фрейме казино. Как и в случае с фуражировкой, участники не получали отзывов о результатах своего выбора.В дополнение к инструкции, два фрейма различались тем, что варианты в фрейме поиска были представлены как последовательности азартных игр, когда количество дней было больше единицы, тогда как варианты в фрейме казино всегда были одношаговыми играми. Чтобы инструкции по поиску пищи не влияли на поведение в фрейме казино, фрейм казино предшествовал фрейму фуражировки для всех участников.
Результаты
Общее сравнение семейств моделей на обеих рамах
Сначала мы спросили, объясняют ли модели, основанные на принципах гомеостаза, выбор лучше, чем стандартные экономические модели — как в рамках поиска пищи, так и в рамках казино.Мы объединили варианты из обеих рамок и сравнили три семейства формальных моделей принятия решений. Первые два семейства моделей включали вариации двух типов экономических моделей, а третье семейство — модели, основанные на гомеостатических соображениях (подробности см. В методе , и таблице 2).
В соответствии с нашей гипотезой о том, что решения участников должны учитывать вероятность голода (стр. голодать ), семейство гомеостатических моделей обеспечило значительно лучшее соответствие.При предположении, что разные участники могут использовать разные модели (анализ случайных эффектов), вероятность превышения того, что семейство гомеостатических моделей является наиболее частым в популяции, составила 0,9403 (таблица 3). При предположении, что все участники используют одну и ту же модель (анализ фиксированных эффектов), победившая модель принадлежала к гомеостатическому семейству (см. Рис. 2A и таблицу 2 для лог-групповых байесовских факторов на основе байесовского информационного критерия (BIC) по сравнению с простейшая модель; см. Раздел 7 текста S2 и Таблицу S1 и Таблицу S2 для результатов, основанных на информационном критерии Акаике, AIC; см. Таблицу S3 для соответствия различных моделей для каждого отдельного участника).Таким образом, общее сравнение модельных семейств подтвердило нашу гипотезу о том, что вероятность голодания и, следовательно, гомеостатические принципы обеспечивают объяснительную силу в объяснении поведения участников, помимо экономических переменных, и хотя они не имеют отношения к максимизации полезности в лаборатории.
Рис 2. Результаты сравнения моделей.
(A) Лог-групповые байесовские факторы (чем меньше, тем лучше) по сравнению с простейшими моделями (Модель 1) на основе BIC для девяти протестированных моделей ( левая часть ) и гистограммы наиболее эффективных моделей на участника ( правая часть ).Модели принадлежали трем семействам. В первом семействе модели основывались на различных комбинациях весовых параметров для первых трех статистических моментов (т.е. EV, Var и Skw). Второе семейство включает две ранг-зависимые полезные модели, в которых вероятности взвешивались в соответствии с нелинейными весовыми функциями (Prelec-I и Prelec-II). В третьем семействе модели основывались на гомеостатическом принципе минимизации p голодания в дополнение к комбинациям весовых параметров для статистических моментов.В обоих кадрах третье семейство моделей лучше всего соответствовало данным. Для удобства первые две модели не изображены, потому что их логарифмические байесовские факторы были далеки от других моделей. Гистограмма показывает, что у 11 из 22 участников самая эффективная модель принадлежала к третьему семейству моделей. Обратите внимание, что анализ фиксированных эффектов не учитывает возможные выбросы, в то время как анализ случайных эффектов учитывает. Меньшие коэффициенты Байеса лог-группы указывают на большее количество доказательств для соответствующей модели по сравнению с базовой моделью.См. Также Таблицу 2. (B) Оценки параметров из адаптации модели общего выигрыша для кормодобывания-p голодать (ξ добывать пищу ) и казино-p голодать (ξ казино ) для отдельных участников. Как и ожидалось из представления о том, что участники должны минимизировать p голодать , большинство оценок параметров были отрицательными (то есть в нижнем левом квадранте). Кроме того, для большинства участников foraging-p голодать было более отрицательно, чем у casino-p голодать (т.е.д., большинство точек лежит ниже линии тождества). См. Также S1 Fig для данных о выборе сгруппированных значений.
https://doi.org/10.1371/journal.pcbi.1004301.g002
Сравнение семейств моделей по типу рамы
Затем мы отдельно проанализировали варианты выбора в фрейме сбора пищи и в чисто денежном контексте фрейма казино, сравнивая три семейства моделей в каждом фрейме. Возникла та же общая картина. Гомеостатическое семейство, в которое включены модели p голодает , имело самые высокие вероятности превышения в обоих кадрах независимо (поиск пищи: 0.9869; казино: 0,8675; Таблица 3). Кроме того, модели, победившие в анализе фиксированных эффектов, принадлежали к семейству гомеостатических (см. Таблицу 2 для лог-групповых байесовских факторов на основе BIC). Далее, когда мы подогнали модели отдельно для первого и второго блоков фреймов фуражиров и казино, во всех анализах прослеживалась одна и та же картина. Гомеостатическое семейство имело самые высокие вероятности превышения, и модели, принадлежащие к этому семейству, выиграли анализ фиксированных эффектов (см. Таблицы 2 и 3; см. Раздел 7 текста S2, Таблицу S1 и Таблицу S2 для результатов, основанных на AIC).
Сравнение в семействе моделей-победителей
В рамках семейства моделей-победителей мы проанализировали, какая конкретная модель лучше всего объясняет выбор. При анализе случайных эффектов вероятность превышения простейшей гомеостатической модели составила 0,9786 для обеих рамок (таблица 4). Аналогичные результаты были получены при подгонке моделей отдельно в двух фреймах или отдельно для первого и второго блоков (таблица 4; см. Раздел 7 текста S2 и таблицу S4 для результатов, основанных на AIC; см. Рис. S1 для данных выбора с разбивкой по ячейкам).В этой выигрышной модели переменная решения представляла собой линейную комбинацию разницы в вероятности голодания (p голодать ), взвешенной с помощью параметра ξ, и разницы в ожидаемом значении (EV). Эта переменная решения была преобразована в вероятность решения сигмовидной функцией с другим параметром β.
Сравнение параметров в адаптированной версии победившей модели
Предыдущий анализ показал, что участники последовательно использовали модели, основанные на принципах гомеостаза в обеих рамках.Это оставляет открытым вопрос, как участники использовали p голодать и отличалось ли это между двумя рамками. Следовательно, мы добавили свободные параметры, зависящие от кадра, в модель-победитель, а затем протестировали оценки параметров среди участников.
В частности, мы адаптировали модель-победитель (Модель 7), которая включала два свободных параметра: параметр β для шума решения и параметр ξ для количественной оценки воздействия p голодания на решение участников. Этот параметр ξ был заменен двумя весовыми параметрами, зависящими от кадра (ξ для поиска пищи и ξ для казино ).Мы добавили эту модель (Модель 10) к первоначальному набору из трех моделей третьего семейства. Несмотря на наказание за дополнительный свободный параметр, он объяснил выбор лучше, чем три другие рассматриваемые модели. Вероятность его превышения составила 0,9983 в анализе случайных эффектов, и он имел наименьший лог-групповой байесовский фактор в анализе фиксированных эффектов (таблица 5; см. Раздел 7 текста S2 и таблицу S5 для результатов, основанных на AIC). Это указывает на то, что участники взвешивали p , голодающие по-разному в двух кадрах.
Важно отметить, что наш прогноз о том, что выбор участников сводит к минимуму p голодать , требует, чтобы весовые параметры p голодать были отрицательными. Действительно, параметры для конкретных кадров ξ , собирая пищу и ξ , казино , были значительно меньше нуля для всех участников (знаковый тест параметров в общей модели выигрыша: ξ , добыча пищи : p <0,001; и ξ ). казино : p <0,005; рис. 2B). То есть участники выбирали игры с меньшим p голодать и, таким образом, минимизировали p голодать .Кроме того, для всех участников ξ , добывающий корм, был меньше, чем ξ , казино (тест знаков, сравнивающий ξ , собирающий корм, и ξ , казино : p <0,05).
В соответствии с вышеприведенным анализом, подтверждающий анализ показал, что p голодать играет большую роль, чем EV в фуражировке, в то время как в фрейме казино EV играет большую роль, чем p голодать , для объяснения выбора (см. Текст S2 Раздел 1 и Таблица S6 для подробностей). Кроме того, мы разработали дополнительную модель, чтобы проверить, приводит ли различное количество дней кормодобывания к разному весу p голодать , но не нашли доказательств, подтверждающих эту идею (текст S2, раздел 2 и таблица S7).Мы также не нашли доказательств, подтверждающих гипотезу о том, что различные комбинации уровней энергии и дней кормодобывания приводят к разному весу p голодать (S2 Text Раздел 4). Дополнительный анализ показал, что участники не включали ошибочно значения ниже нуля в свою оценку распределений результатов в кормодобывающем хозяйстве (раздел 5 текста S2 и таблица S8). В ходе исследовательского анализа мы также не нашли доказательств связи параметров модели с метакогнитивными оценками участников по шкале отношения к риску, зависящей от предметной области (S2 Text Раздел 8).
В совокупности участники последовательно минимизировали p голодание в обоих кадрах, и делали это больше в поисках пищи, чем в кадре казино.
Анализ времени реакции
Может ли время реакции (RT) как предварительная мера трудности выбора дать нам дополнительные доказательства актуальности гомеостатических принципов? Поскольку наше сравнение моделей показало, что EV и p от голода объяснили выбор участников, мы проверили, связаны ли EV и p с голоду с RT.То есть мы проверили, были ли RT быстрее для больших абсолютных различий между двумя вариантами в EV и p голодать . Это действительно имело место, как показывает модель линейных смешанных эффектов для логарифмически преобразованных RT (EV: t = -4,98, p <0,001; p голодание : t = -2,62, p <0,05; уровни значимости были определены с помощью тестов логарифмического правдоподобия, сравнивая полную модель с моделью без соответствующего фактора). Взаимодействие EV и p голодать было значительным и связано с более медленными RT (t = 3.52, р <0,005). См. S1 Fig для бинированных данных RT. В целом, комбинация EV и p голодать была связана с трудностью выбора, индексированной RT, что подтверждает, что при выборе участников руководствовались принципами гомеостаза.
Обсуждение
В этом исследовании рассматривается вопрос о том, объясняют ли принципы гомеостатики принятие решений человеком по сравнению с ранее описанными экономическими моделями, основанными на максимизации полезности конечной точки. Мы обнаружили, что человеческие решения сводят к минимуму вероятность достижения нижней гомеостатической границы на траектории их конечных результатов, несмотря на то, что наши задачи не повлекли за собой каких-либо явных негативных последствий достижения этой границы.Это было очевидно как в задаче виртуального сбора пищи, в которой возможность голода была важной особенностью задачи, так и в рамках, подобной казино, в которой явно указывались только конечные результаты азартных игр и связанные с ними вероятности. Наше детальное сравнение моделей доказало, что переменная решения в самой экономной модели была основана на линейной комбинации вероятности голода и ожидаемого значения конечной точки (EV), что превосходит стандартные экономические модели.
Максимизация конечной точки EV лежит в основе многих вариантов аксиоматически выведенных микроэкономических моделей. Однако ни варианты моделей риска-доходности, ни варианты теории ожидаемой полезности не предсказывают, что решения минимизируют вероятность достижения нижней гомеостатической границы до того, как эта конечная точка будет реализована. Семейство моделей-победителей включало вероятность нулевых результатов, хотя мы действительно побуждали участников избегать их, и хотя нулевые результаты уже включены в расчет статистических моментов и полезностей.Для описания поведения мы могли бы использовать очень специфическую форму функции полезности, которая приписывает высокую отрицательную полезность нулевому результату и положительную полезность соседнему положительному результату, в отличие от типичных функций полезности в экономической литературе. Однако такая модель не была бы более скупой, чем наша, и не предлагала бы никаких дополнительных объяснительных возможностей.
Отметим, что в модели наилучшего соответствия переменная решения представляла собой линейную смесь переменных результата и, таким образом, не отличалась от предыдущих моделей риска и доходности по своей математической структуре.Важно отметить, что минимизация вероятности нулевого исхода обеспечила большую объяснительную силу, чем отношение к риску, основанное на дисперсии или асимметрии. Таким образом, наши результаты согласуются с предыдущими отчетами, требующими более детальных и, возможно, контекстно-зависимых показателей в рамках моделей риска-доходности [19,27]. Кроме того, наша модель делает осмысленные прогнозы только тогда, когда вероятность угроз гомеостазу отлична от нуля, и, таким образом, наш подход имеет желаемую особенность, заключающуюся в том, что объем модели находится в строго определенных ограничениях.
Мы предоставляем доказательства того, что гомеостатический принцип избегания нижней границы энергетических уровней пронизывает процесс принятия решений человеком. Классические описания часто связывают гомеостатические процессы с действием термостата [6,7]. Пример термостата лучше всего подходит для физиологических переменных с узким гомеостатическим диапазоном, для которого этот диапазон может быть аппроксимирован заданным значением (например, pH крови) [7]. Для других переменных гомеостатический диапазон больше. В случае метаболического гомеостаза гликогеновые и жировые буферы увеличивают гомеостатический диапазон, и соответствующие гомеостатические контрмеры возникают на границах этого диапазона [7].Для простоты мы предположили, что голод является жесткой границей, но тот же принцип применим и к мягким границам. Что еще более важно, наши результаты расширяют понятие, проиллюстрированное аналогией с термостатом. В соответствии с недавними теоретическими взглядами на гомеостаз у здоровых и психиатрических популяций [8,10], мы предположили, что люди, принимающие решения, могут оценить вероятность будущих нарушений гомеостаза. Таким образом, в отличие от термостата, который может реагировать на угрозы гомеостазу только после того, как они возникли, люди, принимающие решения, и, возможно, многие животные, могут проактивно избегать угроз гомеостазу.
Одна и та же модель лучше всего показала себя как в фуражке, так и в раме казино. Мы подчеркиваем это сходство между двумя рамками, потому что оно показывает, что гомеостатический принцип минимизации вероятности нулевого результата действует даже тогда, когда участники не ориентированы на это описанием задачи. Более того, та же модель объясняет поведение в последовательности азартных игр и в одноэтапных играх. В кадре казино вероятность голода напрямую отображалась размером сектора на круговой диаграмме, который указывал на вероятность нулевого исхода.Поразительно, что в рамке для сбора пищи участники интегрировали вероятности получения энергии за указанное количество дней, чтобы вычислить вероятность голода. Участники не могли узнать распределение результатов на собственном опыте, потому что мы не предоставили им обратную связь. Таким образом, решения в рамках кормодобывающей среды не зависели от участников, непосредственно испытавших последовательности в виртуальных кормовых средах. Принятие рискованных решений зависит от того, описано ли распределение результатов или получено из опыта [22,28].Например, редкие события, как правило, оказывают меньшее влияние на решения, основанные на опыте. Наши результаты предполагают, что такого недооценки вероятности голода не могло быть [28].
В рамках модели-победителя более детальный анализ выявил различия между двумя рамками в наиболее подходящих оценках параметров. Вероятность голодной смерти в фрейме поиска пищи имела большее влияние на решения участников, чем соответствующая вероятность ничего не получить в фрейме казино — эффект, не связанный с последовательным или пошаговым представлением азартных игр (см. Раздел 3 текста S2).Это имело место, несмотря на то, что участникам приходилось вычислять вероятность голода в кормодобывающем хозяйстве, комбинируя информацию о внутреннем состоянии, вариантах кормодобывания и временном горизонте. Приближение вероятностей голода может стать более трудным и, следовательно, неточным по мере увеличения количества шагов. В текущем исследовании участники смогли приблизительно оценить вероятность голода с достаточной точностью, по крайней мере, для трех шагов, поскольку их решения были основаны на этой метрике.
Структура наших задач соответствует требованиям экономических парадигм, таких как полное знание и совместимость по стимулам [12].Таким образом, характеристики конкретных задач вряд ли могут объяснить, почему наша гомеостатическая модель превзошла стандартные экономические модели, основанные на статистических моментах [15,17,18] или нелинейном вероятностном взвешивании [23,24]. Вместо этого мы полагаем, что биологические ограничения, актуальные в экологических контекстах, таких как охота или сельское хозяйство, оказывают преобладающее влияние на решения человека в лаборатории, даже если они явно не имеют отношения к поставленной задаче. Подобное обоснование недавно было выдвинуто при обсуждении вопроса о том, отклоняется ли принятие решений животными [29,30] и людьми [31–34] от нормативных моделей.Согласно вероятностным объяснениям функции мозга, мозг использует априорные вероятности для выполнения вероятностных выводов [8,14,35,36]. Эти априорные вероятности настраиваются — в результате эволюции и / или опыта — на естественную статистику окружающей среды реального мира [30,31]. Следовательно, люди, принимающие решения, и люди, не являющиеся людьми, могут вести себя рационально в соответствии со своими убеждениями, но они кажутся иррациональными, потому что эти убеждения не оправданы в намеренно упрощенных лабораторных задачах или в некоторых других контекстах [30,37].В целом, этот недавний подход требует дополнения соображений об экономической рациональности (т. Е. Максимизации финансовой выгоды с учетом имеющейся в настоящее время информации) соображениями экологической рациональности (то есть максимизации пригодности с учетом априорных значений экологической статистики). Его обещание заключается в объединении и объяснении разнообразного набора, казалось бы, иррационального поведения, в то время как его задача заключается в выявлении и проверке экологических принципов, на которых основываются такие объяснения [30].
Настоящее исследование демонстрирует, что базовый биологический принцип, касающийся внутренней среды, дает уточненное и экономное объяснение человеческих решений в условиях риска.Наша виртуальная задача по поиску пищи была специально разработана для проверки влияния принципов гомеостаза на рискованный выбор. Таким образом, он относится к предыдущим тестам теории кормодобывания с учетом риска на животных и расширяет их [38–41]. Чувствительная к риску теория кормодобывания дает представление о том, как животное должно выбирать между рискованными вариантами кормодобывания, чтобы максимизировать свою приспособленность [40]. Важнейшее понимание теории кормодобывания, чувствительного к риску, состоит в том, что кормящим животным следует выбирать варианты с более высокой дисперсией, если варианты с меньшей дисперсией не могут обеспечить достаточное количество энергии для достижения критических уровней до определенного момента времени.Например, голодные птицы зимой должны стать более подверженными риску с приближением темноты. Таким образом, чувствительная к риску теория кормодобывания обеспечивает экологически рациональный ориентир [38–40], хотя эмпирические данные в его пользу неоднозначны [40]. Подобно теории кормодобывания, чувствительной к риску, наша модель содержит жесткую границу, которая актуальна для лиц, принимающих решения, в пределах заданного временного горизонта. Важно отметить, что мы представляем новое и простое математическое описание для получения последовательных азартных игр, которые отражают условия поиска пищи.Тестирование модели в виртуальной среде на людях позволяет обойти проблемы, связанные с исследованиями на животных, не относящихся к человеку, такие как необходимость навязывать участникам реальные угрозы или необходимость сообщать распределение результатов посредством обширного обучения.
Теория кормодобывания, чувствительная к риску, была связана с неприятием потерь [40], что отсылает к эмпирическому наблюдению, что люди, кажется, больше заботятся о потерях, чем о прибылях эквивалентной величины [40,42]. Можно предположить, что неприятие потерь может быть связано с нашим выводом о том, что участники минимизировали вероятность голода.Однако неприятие проигрышей может возникнуть только в смешанных играх (то есть, когда опционы влекут за собой прибыли и убытки) [23,42], и наши игры не предполагали потерь. Таким образом, неприятие потерь не может объяснить наши выводы.
Наш подход согласуется с некоторыми недавними исследованиями, в которых использовались задачи, подобные виртуальному поиску пищи, для исследования психологических и нейронных механизмов принятия сложных решений у животных [41,43] и людей [44–46]. Одно известное исследование показало, что люди динамически корректируют свое рискованное поведение в зависимости от последовательности азартных игр [44].Другое исследование предоставляет доказательства того, что люди постоянно переоценивают последовательность азартных игр, доступных им в будущем, хотя экономически оптимальные стратегии предписывают, чтобы решения не зависели от порядка следования [47]. Наши результаты дополняют эти выводы, предполагая, что такое поведение можно легко объяснить, если люди примут во внимание вероятность «голодной смерти» во время последовательности выбора. В целом, текущее исследование дает подробные прогнозы очевидной иррациональности в задачах динамического поиска пищи, которые согласуются с более ранними отчетами.
Наша модель принятия гомеостатических решений допускает возможные расширения. Во-первых, лица, принимающие решения, обычно должны поддерживать несколько переменных в гомеостатическом диапазоне. Наша модель может быть легко расширена на такие ситуации с предсказанием, что лица, принимающие решения, минимизируют общую вероятность голода, что может означать отказ от большого количества одной переменной, чтобы избежать обнуления другой. Когда границы мягкие, а не жесткие, это можно рассматривать как минимизацию ограниченного функционала, который описывает траекторию через гомеостатическое пространство.Во-вторых, предпочтения к риску часто считаются довольно стабильными чертами личности [48], но наша модель предполагает, что они должны варьироваться в зависимости от угроз гомеостазу [38]. В-третьих, страхование на случай редких событий со значительными последствиями стало в последнее время в центре внимания экономики [49]. Концепция голода в нашей модели может помочь в исследовании влияния таких событий на человеческие решения.
Наши результаты показывают, что стремление к этой фундаментальной биологической цели трансформируется в простые, но конкретные прогнозы для принятия решений, которые поддаются эмпирическим проверкам.Стандартные экономические модели обеспечивают незаменимый ориентир, по которому можно проверить включение дополнительных соображений, касающихся биологических соображений [12,15,23]. Обычно модели принятия рискованных решений должны обеспечивать баланс между элегантностью аксиоматических экономических основ, которые расходятся с эмпирическими наблюдениями, и громоздкими произвольными предположениями об иррациональных предубеждениях. Наши результаты показывают пример того, что модели, основанные на фундаментальных биологических принципах, таких как гомеостаз, могут согласовать экономию с объяснением очевидных иррациональностей.
Методы
Заявление об этике
Исследование было проведено в соответствии с Хельсинкской декларацией и одобрено правительственным комитетом по этике исследований (Kantonale Ethikkommission Zürich, KEK-ZH-Nr. 2013–0328). Все участники дали письменное информированное согласие, используя форму, утвержденную комитетом по этике.
Участники
Двадцать два участника (15 женщин; возраст: средний = 25 лет, SD = 5,0) были набраны из числа студентов через списки рассылки местных университетов.Участникам был выплачен сбор за участие в 15 швейцарских франков плюс переменная сумма (см. Ниже).
Задача
Участники выполнили 960 испытаний в двух вариантах (фреймах) задачи бинарного выбора: фреймы поиска и фреймы казино (рис. 1A и 1B). Один и тот же список из 480 комбинаций азартных игр использовался для обоих фреймов (то есть распределения результатов были численно равны; см. Таблицу 1 для обзора переменных; см. Ниже и S1 Text для подробностей о том, как были получены азартные игры). Для обоих кадров участники получили подробные письменные инструкции и выполнили восемь тренировочных испытаний, за которыми следовали два блока фактического задания.Задача была представлена с использованием набора инструментов MATLAB Cogent 2000 (www.vislab.ucl.ac.uk). В инструкции для фуражировки участникам предлагалось представить себя в контексте охотника-собирателя. Поскольку мы хотели исключить то, что установка участников на фуражированное мышление влияла на выбор в фрейме казино, все участники завершили фрейм казино раньше фрейма фуражировки. 480 игровых комбинаций для каждого кадра были разделены на два блока, которые были уравновешены по порядку. Каждый список содержал 80 уникальных игровых комбинаций; остальные 400 игровых комбинаций были включены в оба списка.Во время игры участники не видели результатов своего выбора. То есть участникам были даны примеры возможных результатов в письменной инструкции, но они не испытали их на собственном опыте. В конце эксперимента случайным образом выбирали по одной пробе из каждого из четырех блоков. Результаты этих испытаний были определены на основе выбора участников, и была выплачена соответствующая сумма (1 балл стоил 0,75 швейцарского франка). Таким образом, оба фрейма были стимулированы одинаково. См. Рис. 1 и текст S2 для получения дополнительных сведений.
Гэмблс
Мы использовали математику случайных блужданий, чтобы получить распределения результатов для 480 комбинаций азартных игр (таблица 1). Кратко представим основную логику (рис. 1С ) . Подробнее см. S1 Text. Во время случайного блуждания воображаемый агент начинает с заданной позиции в строке положительных целых чисел. Исходное положение соответствует начальному количеству очков энергии. Агент выполняет в этой строке количество шагов, которые соответствуют количеству дней.На каждом шаге агент перемещается «вправо» с определенной вероятностью p и «влево» с вероятностью q = 1-p. Движение влево соответствует неудачному поиску пищи, а размер шага соответствует фиксированной стоимости одного очка энергии. Движение вправо соответствует успешному поиску корма, а размер шага соответствует разным очкам, которые нужно набрать (за вычетом стоимости одного очка). Ноль представляет собой поглощающую границу (т.е. если агент достигает нуля, случайное блуждание прекращается). Возможные позиции на числовой прямой после определенного количества шагов соответствуют диапазону исходов.Чтобы получить вероятность результата, необходимо суммировать все вероятности всех «ветвей на дереве», ведущие к этому исходу. (Количество «ветвей» рассчитывается с помощью биномиального коэффициента.) Вдоль данной ветви необходимо умножить вероятности (то есть p или q). Мы создали разные азартные игры, варьируя комбинации стартовых позиций, вероятностей движения вправо и размеров шага вправо. В текущее исследование мы включили азартные игры с четырьмя различными комбинациями начальных позиций и количества шагов (a) начальная позиция 1 и шаг 1, (b) начальная позиция 1 и шаги 2, (c) начальная позиция 2 и шаг 2, и (г) начальные позиции 2 и 3 шага (каждая комбинация встречается в 120 комбинациях игр).Мы использовали результаты и их соответствующие вероятности для расчета статистических моментов выбранных игр. Вероятности достижения нуля обозначены p голодать . Обратите внимание, что в играх, включенных в текущее исследование, p голодать никогда не было нулевым (см. Таблицу 1).
Модели
Обзор.
В среднем участники пропустили 2,0 испытания из 960 в общей сложности (SD = 3,3). Мы определили наиболее подходящие параметры модели, используя оценку максимального правдоподобия (MLE).Оптимизация использовала нелинейный алгоритм симплексного поиска Нелдера-Мида (реализованный в функции MATALB fminsearch) для минимизации отрицательной логарифмической вероятности, суммированной по всем испытаниям для каждого участника. Сначала мы запустили оптимизацию с положительными начальными параметрами. В случае несовпадения мы засевали модели с отрицательными стартовыми параметрами. Логарифмическое правдоподобие было получено из наиболее подходящих параметров. Для каждой модели мы аппроксимировали свидетельство модели путем вычисления байесовского информационного критерия (BIC), который снижает сложность модели (см. Раздел 7 в тексте S2 для результатов, основанных на информационном критерии Акаике, AIC).
Во всех моделях использовалась функция логистики / softmax со свободным параметром β для генерации пробных вероятностей для выбора одного из двух вариантов, который учитывает шум при выборе действия и концептуально аналогичен логистической регрессии.
(1.1)V 1 и V 2 — значения двух опций, указанные в моделях. Следующее описание следует порядку моделей, указанному в таблице 2. Первоначально мы разработали девять моделей, которые были сгруппированы в три семейства моделей.
Вкратце, первые два семейства моделей включали вариации двух типов экономических моделей, в то время как третье семейство моделей включало новые модели, основанные на гомеостатических соображениях. В четырех моделях первого семейства переменная решения была основана на различных линейных комбинациях статистических моментов распределения результатов (ожидаемое значение, EV; дисперсия, Var; и асимметрия, Skw). Во второе семейство вошли два варианта рангово-зависимых полезных моделей. В трех моделях третьего семейства переменная решения была основана на вероятности голодания (p голодать ) в дополнение к различным линейным комбинациям статистических моментов (см. Таблицу 2).
Семейство моделей 1: Моменты без р.
голодают .В первом семействе модели (модели 1–4) основывались на статистических моментах распределения результатов. В простейшей модели (Модель 1) только параметр β был включен в качестве свободного параметра для сравнения EV между двумя вариантами, и, таким образом, V просто равно EV варианта.
(1.2)Три другие модели (модели 2–4) первого семейства дополнительно включали линейные комбинации Var и Skw, каждая из которых была взвешена соответствующими свободными параметрами.То есть участникам разрешено выражать различные предпочтения в отношении этих переменных, и каждая переменная взвешивается свободным параметром (ρ или λ).
(1,3) (1,4) (1,5)Семейство моделей 2: полезность, зависящая от ранга.
Второе семейство (модели 5–6) состояло из двух вариантов моделей полезности, зависящих от ранга [23], в которых вероятности ранжированных результатов взвешивались нелинейно (и в которых результаты преобразовывались в полезности с использованием экспоненциальной функции). с одним свободным параметром).В частности, варианты поиска пищи содержали J результатов x 1 , x 2 ,…, x J с соответствующими вероятностями p 1 , p 2 ,…, стр. J . В моделях полезности, зависящей от ранга, варианты оценивались в соответствии с (1.6) u ( x j ) — возрастающая функция полезности по денежным результатам с u (0) = 0.В соответствии со многими спецификациями в литературе мы выбираем степенную функцию со свободным параметром μ.
(1,7)Веса решений π j были определены в зависимости от w ( p j ), которая представляет собой функцию взвешивания с возрастающей вероятностью с w (0) = 0, w (1) = 1 и.
(1.8)Одна из двух зависимых от ранга полезных моделей (Модель 5) использовала весовую функцию, определенную Prelec-I, которая включает один свободный параметр α [23].
(1.9)Другая из двух зависимых от ранга полезных моделей (Модель 6) использовала весовую функцию, определенную Prelec-II, которая включает два свободных параметра и, таким образом, допускает более гибкие формы, чем Prelec-I [23]. Дополнительный свободный параметр называется β для согласованности (не путать с β логистической функции).
(1.10)Семейство моделей 3: Моменты и р
голодают .В третьем семействе модели (модели 7–9) включали свободный параметр для p starve (ξ) в дополнение к комбинациям статистических моментов.Таким образом, модели этого третьего семейства включают параметр свободного веса, выведенный из гомеостатических принципов.
(1.11) (1.12) (1.13)Сравнение моделей.
Мы аппроксимировали данные модели, используя байесовский информационный критерий (BIC). Кроме того, мы сообщаем информационный критерий Акаике (AIC). Они были рассчитаны для каждого участника и модели в соответствии со следующими уравнениями. (1.14) (1.15) где L — функция правдоподобия, k — количество параметров в модели, а n — количество точек данных соответствующего участника.Во-первых, мы провели анализ случайных эффектов в предположении, что разные участники могут использовать разные модели или семейства моделей. Мы использовали процедуру выбора байесовской модели (BMS), реализованную в VBA-toolbox (http://mbb-team.github.io/VBA-toolbox) для расчета вероятностей превышения, которые измеряют, насколько вероятно, что любая данная модель чаще, чем все другие модели в популяции [50–52]. Мы проверили семейство моделей, наиболее подходящих по размеру, а затем — модель, наиболее подходящую для семейства-победителей.Во-вторых, мы провели анализ фиксированных эффектов, исходя из предположения, что все участники используют одну и ту же модель. Мы вычислили логарифмические байесовские факторы, вычитая AIC / BIC для эталонной модели из AIC / BIC каждой протестированной модели, и просуммировали их по группе, чтобы получить лог-групповые байесовские факторы (см. Основной текст для анализа на основе BIC; см. Раздел 7 текста S2 и таблиц S1, S2, S4 и S5 для анализа на основе AIC). Согласно принятому здесь соглашению, меньшие байесовские факторы логарифмической группы указывают на большее количество доказательств для соответствующей модели по сравнению с базовой моделью.Для сравнения оценок параметров мы использовали точный метод знакового теста, реализованный в MATLAB.
Дополнительные модели.
Поскольку модель, основанная на электромобиле и p starve (модель 7) была общей выигрышной моделью среди начальных девяти моделей, мы адаптировали ее и протестировали десятую модель (модель 10), которая включала два отдельных свободных параметра для p starve : добыча пищи — p голодать и казино — p голодать .
(1.16)Где f = 1 для всех вариантов в фрейме поиска пищи и f = 0 для всех вариантов в фрейме казино.
Мы дополнительно протестировали две дополнительные модели. Результаты, относящиеся к этим моделям, описаны в разделах 1 и 2 текста S2. Модель 11 — это вариант Модели 7 и Модели 1, в которой стоимость опции просто задается p голодать , а не ее EV.
(1.17)Модель 12 — это вариант Модели 7, в которой используются три отдельных свободных параметра для трех различных количеств дней сбора пищи в играх (соответствующих количеству вариантов).
(1.18)Где d 1 = 1 для всех игр с одним днем и d 2 = 0 и d 3 = 0 в противном случае. Аналогично, d 2 = 1 для игр с двумя днями и d 3 = 1 для игр с тремя днями.
Связь между р
голод и перекос.Мы отмечаем, что модель, основанная на EV и p голодать (Модель 7), превзошла модели, включая параметры, которые взвешивали разницу в асимметрии (Модель 4, Модель 9), даже несмотря на то, что различия в асимметрии и вероятности голода были коррелированы (Пирсон г =.901). Хотя эта корреляция могла привести к завышенным ошибкам оценок параметров в модели, которая включала линейные весовые коэффициенты для p голод и Skw (Модель 9), она не ставила под угрозу оценку модели и, следовательно, сравнение моделей для моделей, которые отдельно включены весовые коэффициенты для p starve (модель 7, модель 8) или Skw (модель 3, модель 4). Кроме того, в рамках Модели 9 коэффициенты инфляции дисперсии для всех индивидов лежат ниже 8.
Анализ РЦ
логарифмически преобразованных RT были проанализированы с использованием линейной модели смешанных эффектов, реализованной в пакете R lmer [53] (http: // cran.r-project.org/web/packages/lme4/index.html). Логарифмически преобразованные RT были приблизительно нормально распределены. Независимыми переменными в модели смешанных эффектов были переменные, которые при сравнении моделей выбора были определены как релевантные (т. Е. EV и p голодают ). В частности, фиксированные эффекты модели включали разницу между двумя вариантами в EV и p голодать , а также их взаимодействие. Случайные эффекты для участников включали случайный перехват и случайные наклоны для EV, p голод и их взаимодействие.Модель задается следующим уравнением:
(1.19)Уровни значимости фиксированных эффектов были определены путем выполнения тестов логарифмического правдоподобия, в которых полная модель сравнивалась с моделями без соответствующего фактора.
Вспомогательная информация
S1 Рис. Выбор и данные RT, объединенные в соответствии с различиями в p
, истощают и EV.(A) Мы сгруппировали данные в соответствии с разницей в p голодать между двумя вариантами выбора и средними значениями графика для участников.Планки погрешностей указывают стандартные ошибки среднего. Верхний ряд: поскольку количество испытаний в разных ячейках было нечетным, мы предоставляем гистограммы. Средний ряд: чем выше была разница в p голодать между двумя вариантами выбора, тем более вероятно, что участники выбрали вариант с меньшим значением p голодать . Средний уклон был более отрицательным при добыче корма по сравнению с рамой казино. Нижний ряд: время реакции модулировалось с помощью p starve .
(B) Мы сгруппировали данные в соответствии с разницей в EV между двумя вариантами выбора.Условные обозначения как в A. Чем выше разница в EV между двумя вариантами выбора, тем больше вероятность, что участники выберут вариант с более высоким EV.
(C) Мы объединили данные в соответствии с различиями в p голодающих и в EV. Чтобы получить достоверное количество испытаний в каждой ячейке, мы включаем только «средние» ячейки для различий в p голодать . Для различий в EV мы выполнили медианное разделение.
https://doi.org/10.1371/journal.pcbi.1004301.s003
(TIF)
Благодарности
Мы благодарим Маттиаса Стаиба, Джузеппе Кастегнетти, Атину Цовару, Юлию Оганян и Питера Мора за полезные обсуждения и комментарии к более раннему варианту этой рукописи.
Вклад авторов
Задумал и спроектировал эксперименты: CWK DRB. Проведены эксперименты: CWK. Проанализированы данные: CWK DRB. Написал статью: CWK DRB.
Ссылки
- 1. Пушка WB. Организация физиологического гомеостаза. Physiol Rev.1929; 9 (3): 399–431.
- 2. Дэвис GW. Гомеостатическая передача сигналов и стабилизация нервной функции. Нейрон. 2013; 80 (3): 718–28. pmid: 24183022
- 3. Фиглевич Д.П., Сиполс А.Дж.Сигналы регулирования энергетики и пищевое вознаграждение. Pharmacol Biochem Behav. 2010. 97 (1): 15–24. pmid: 20230849
- 4. McEwen BS, Wingfield JC. Понятие аллостаза в биологии и биомедицине. Horm Behav. 2003. 43 (1): 2–15. pmid: 12614627
- 5. Рангель А. Регулирование выбора диеты схемой принятия решений. Nat Neurosci. 2013. 16 (12): 1717–24. pmid: 24270272
- 6. Бернсон Г.Г., Качиоппо Дж. Т.. Интегративная физиология: гомеостаз, аллостаз и оркестровка системной физиологии.В: Cacioppo JT, Tassinary LG, Berntson GG, редакторы. Справочник по психофизиологии. 3-е изд. Кембридж: Cambridge University Pres; 2007. с. 433–52.
- 7. Котас М.Е., Меджитов Р. Гомеостаз, воспаление и восприимчивость к болезням. Клетка. 2015; 160 (5): 816–27. pmid: 25723161
- 8. Гу Х, Фитцджеральд THB. Интероцептивный вывод: гомеостаз и принятие решений. Trends Cogn Sci. 2014; 18 (6): 269–70. pmid: 24582825
- 9. Паулюс М.П. Нарушения в принятии решений в психиатрии — изменение гомеостатической обработки? Наука.2007. 318 (5850): 602–6. pmid: 17962553
- 10. Керамати М., Гуткин Б. Сбор награды для защиты гомеостаза: теория обучения гомеостатическому подкреплению. bioRxiv. 2014; 140.
- 11. Фон Нейман Дж, Моргенштерн О. Теория игр и экономического поведения. Принстон, Нью-Джерси: Издательство Принстонского университета; 1953.
- 12. Кагель Дж. Х., Рот А. Э., редакторы. Справочник экспериментальной экономики. Принстон, Нью-Джерси: Издательство Принстонского университета; 1995 г.
- 13. Босартс П. Чему учит нас нейробиология принятия финансовых решений. Annu Rev Financ Econ. 2009. 1 (1): 383–404.
- 14. Бах Д.Р., Долан Р.Дж. Понимание того, сколько вы не знаете: нейронная организация оценок неопределенности. Nat Rev Neurosci. 2012. 13 (8): 572–86. pmid: 22781958
- 15. Тоблер П.Н., Вебер ЕС. Оценка рискованного и неопределенного выбора. В: Glimcher PW, Fehr E, редакторы. Принятие решений нейроэкономики и мозг.2-е изд. Лондон: Academic Press; 2014. с. 149–72.
- 16. Джонсон Дж. Г., Уилке А., Вебер ЕС. За пределами типичного взгляда на принятие риска: шкала для конкретной предметной области, измеряющая восприятие риска, ожидаемые выгоды и отношение к предполагаемому риску среди немецкоговорящего населения. Польский Psychol Bull. 2004. 35 (3): 153–63.
- 17. Симмондс М, Райт Н.Д., Бах Д.Р., Долан Р.Дж. Деконструкция риска: раздельное кодирование дисперсии и асимметрии в мозге. Нейроизображение. 2011. 58 (4): 1139–49.pmid: 21763444
- 18. Симмондс М, Райт Н.Д., Фаган Э., Долан Р.Дж. Анализ влияния леводопы на оценку риска у здоровых людей. PLoS One. 2013; 8 (7).
- 19. Mohr PNC, Biele G, Krugel LK, Li S-C, Heekeren HR. Нейронные основы компромисса между риском и доходностью при принятии инвестиционных решений. Нейроизображение. 2010. 49 (3): 2556–63. pmid: 19879367
- 20. Карако Т., Часин М. Предпочтения в собирательстве: реакция на перекос в вознаграждении. Anim Behav. 1984. 32: 76–85.
- 21. Пролив CE, Hayden BY. Паттерны предпочтений для перекошенных азартных игр у макак-резусов. Biol Lett. 2013; 9: 20130902. pmid: 24335272
- 22. Шенберг Т., Фокс С.Р., Полдрак Р.А. Помните о пробеле: преодоление экономического и натуралистического риска с помощью когнитивной нейробиологии. Trends Cogn Sci. 2011; 15 (1): 11–9. pmid: 21130018
- 23. Фер-Дуда Х., Эппер Т. Вероятность и риск: основы и экономические последствия вероятностно-зависимых предпочтений риска.Annu Rev Econom. 2012; 4 (1): 567–93.
- 24. Сюй М., Крайбич И., Чжао С., Камерер С.Ф. Нейронная реакция на ожидание вознаграждения в условиях риска нелинейна по вероятностям. J Neurosci. 2009. 29 (7): 2231–7. pmid: 19228976
- 25. Канеман Д. Взгляд на суждение и выбор: отображение ограниченной рациональности. Am Psychol. 2003. 58 (9): 697–720. pmid: 14584987
- 26. Camerer CF. Цели, методы и успехи нейроэкономики. Annu Rev Econom. 2013. 5 (1): 425–55.
- 27. Вебер ЕС, Джонсон Э. Внимательное суждение и принятие решений. Annu Rev Psychol. 2009. 60: 53–85. pmid: 18798706
- 28. Хертвиг Р., Эрев И. Пробел между описанием и опытом в рискованном выборе. Trends Cogn Sci. 2009; 13: 517–23. pmid: 19836292
- 29. Кацельник А. Смыслы рациональности. В: Nudds M, Hurley S, редакторы. Рациональные животные? Оксфорд: издательство Оксфордского университета; 2006. с. 87–106.
- 30. Фосетт Т.В., Фалленштейн Б., Хиггинсон А.Д., Хьюстон А.И., Mallpress DEW, Trimmer PC и др.Эволюция правил принятия решений в сложных средах. Trends Cogn Sci. 2014. 18 (3): 153–61. pmid: 24467913
- 31. Гигеренцер Г., Гайсмайер В. Эвристическое принятие решений. Annu Rev Psychol. 2011; 62: 451–82. pmid: 21126183
- 32. Haselton MG, Bryant GA, Wilke A, Frederick DA, Galperin A, Frankenhuis WE, et al. Адаптивная рациональность: эволюционный взгляд на когнитивную предвзятость. Soc Cogn. 2009. 27 (5): 733–63.
- 33. Рэнд Д.Г., Новак М. Сотрудничество между людьми.Trends Cogn Sci. 2013. 17 (8): 413–25. pmid: 23856025
- 34. Даян П. Рациональные иррациональности выбора. Top Cogn Sci. 2014; 6: 204–28. pmid: 24648392
- 35. Фристон К. Принцип свободной энергии: единая теория мозга? Nat Rev Neurosci. 2010. 11 (2): 127–38. pmid: 20068583
- 36. Пуже А., Бек Дж. М., Ма В. Дж., Латам ЧП. Вероятностный мозг: известные и неизвестные. Nat Neurosci. 2013. 16 (9): 1170–8. pmid: 23955561
- 37. Макнамара Дж. М., Фосетт Т. В., Хьюстон А. И..Адаптивный ответ на неопределенность порождает положительные и отрицательные эффекты контраста. Наука. 2013. 340 (6136): 1084–6. pmid: 23723234
- 38. Бейтсон М. Последние достижения в понимании чувствительных к риску кормовых предпочтений. Proc Nutr Soc. 2007. 61 (04): 509–16.
- 39. Касельник А., Бейтсон М. Чувствительность к риску: перекресток теорий принятия решений. Trends Cogn Sci. 1997; 1 (8): 304–9. pmid: 21223933
- 40. Хьюстон А.И., Фосетт Т.В., Mallpress DEW, Макнамара Дж. М..Уточнение взаимосвязи между теорией перспектив и теорией кормодобывания, чувствительной к риску. Evol Hum Behav. 2014; 35 (6): 502–7.
- 41.
Стивенс DW. Экология принятия решений: добыча пищи и экология принятия решений животными. Cogn влияет на поведение Neurosci. 2008. 8 (4): 475–84. pmid: 1
- 42
- 42. Fox CR, Poldrack RA. Теория перспективы и мозг. В: Глимчер П., Фер Э, редакторы. Принятие решений нейроэкономики и мозг. 2-е изд. Лондон: Academic Press; 2014. с. 533–67.
- 43. Хайден Б., Пирсон Дж. М., Платт М. Л.. Нейронная основа последовательных решений о поиске пищи в неоднородной среде. Nat Neurosci. 2011; 14 (7): 933–9. pmid: 21642973
- 44. Коллинг Н., Виттманн М., Рашворт MFS. Множественные нейронные механизмы принятия решений и их конкуренция под изменяющимся давлением риска. Нейрон. 2014. 81 (5): 1190–202. pmid: 24607236
- 45. Коллинг Н, Беренс TEJ, Марс РБ, Рашворт МФС. Нейронные механизмы кормодобывания.Наука. 2012. 336 (6077): 95–8. pmid: 22491854
- 46. Бах Д.Р., Гитарт-Масип М., Паккард П.А., Миро Дж., Фалип М., Фуэнтемилла Л. и др. Человеческий гиппокамп разрешает конфликт приближения и избегания. Curr Biol. 2014; 24 (5): 541–7. pmid: 24560572
- 47. Симмондс М, Босартс П., Долан Р.Дж. Поведенческая и нейронная оценка предполагаемого принятия решений в условиях риска. J Neurosci. 2010. 30 (43): 14380–9. pmid: 20980595
- 48. Бец Н.Е., Вебер ЕС. Шкала отношения к риску для конкретной предметной области: измерение восприятия риска и рискованного поведения.J Behav Decis Mak. 2002; 15: 263–90.
- 49. Барберис Н. Психология хвостовых событий: прогресс и вызовы. Am Econ Rev.2013; 103 (3): 611–6. pmid: 25067844
- 50. Стефан К.Е., Пенни В.Д., Даунизо Дж., Моран Р.Дж., Фристон К.Дж. Выбор байесовской модели для групповых исследований. Нейроизображение. 2009. 46 (4): 1004–17. pmid: 19306932
- 51. Rigoux L, Stephan KE, Friston KJ, Daunizeau J. Повторный выбор байесовской модели для групповых исследований. Нейроизображение. 2014; 84: 971–85.pmid: 24018303
- 52. Daunizeau J, Adam V, Rigoux L. VBA: вероятностная обработка нелинейных моделей нейробиологических и поведенческих данных. PLoS Comput Biol. 2014; 10 (1): e1003441. pmid: 24465198
- 53. Баайен Р.Х., Дэвидсон Диджей, Бейтс ДМ. Моделирование смешанных эффектов с перекрещенными случайными эффектами для предметов и предметов. J Mem Lang. 2008. 59 (4): 390–412.
Что такое гомеостаз? — Scientific American
Гомеостаз, от греческих слов «одинаковый» и «устойчивый», относится к любому процессу, который живые существа используют для активного поддержания достаточно стабильных условий, необходимых для выживания.Термин был изобретен в 1930 году врачом Уолтером Кэнноном. Его книга « Мудрость тела » описывает, как человеческое тело поддерживает постоянный уровень температуры и других жизненно важных условий, таких как вода, соль, сахар, белок, жир, кальций и кислород в крови. Подобные процессы динамически поддерживают установившиеся условия в окружающей среде Земли.
Гомеостаз нашел полезное применение в социальных науках. Это относится к тому, как человек в условиях конфликтных стрессов и мотиваций может поддерживать стабильное психологическое состояние.Общество гомеостатически поддерживает свою стабильность, несмотря на конкурирующие политические, экономические и культурные факторы. Хорошим примером является закон спроса и предложения, согласно которому взаимодействие спроса и предложения сохраняет рыночные цены на достаточно стабильном уровне.
Гомеостатические идеи разделяет наука кибернетика (от греческого «рулевой»), определенная в 1948 году математиком Норбертом Винером как «вся область теории управления и коммуникации, будь то в машине или в животном».«Кибернетические системы могут« запоминать »нарушения и, таким образом, используются в информатике для хранения и передачи информации. Отрицательная обратная связь является центральной гомеостатической и кибернетической концепцией, относящейся к тому, как организм или система автоматически противодействуют любому изменению, наложенному на него.
Например, человеческое тело использует ряд процессов для контроля своей температуры, поддерживая ее близкое к среднему значению или норме 98,6 градусов по Фаренгейту. Одна из наиболее очевидных физических реакций на перегрев — потоотделение, которое охлаждает тело, делая кожу доступной для испарения.С другой стороны, тело снижает потери тепла в холодной окружающей среде за счет меньшего потоотделения и уменьшения кровообращения в коже. Таким образом, любое изменение, повышающее или понижающее нормальную температуру, автоматически вызывает противодействующую обратную связь, обратную или отрицательную. Здесь отрицательное означает просто противоположное, а не плохое; на самом деле, в этом примере он действует на наше благополучие. Положительная обратная связь — это реакция на изменение нормального состояния, которая еще больше увеличивает отклонение.
Например, если температура человека повышается примерно до 107 градусов по Фаренгейту, системы отрицательной обратной связи перестают работать.У человека с высокой температурой будет горячая и сухая кожа, если он потеет, чтобы охладить ее. В этом случае не только отключаются системы отрицательной обратной связи; повышенная температура ускоряет химический состав тела, что вызывает еще большее повышение температуры, что, в свою очередь, еще больше ускоряет химический состав тела и т. д. Этот порочный круг положительной обратной связи, процесс «побега» может закончиться только смертью, если его не остановить.
Важно подчеркнуть, что гомеостатические реакции неизбежны и автоматические, если система функционирует должным образом, и что устойчивое состояние или гомеостаз могут поддерживаться многими системами, работающими вместе.Например, приливы крови — это еще одна автоматическая реакция организма на нагревание: кожа краснеет, потому что ее мелкие кровеносные сосуды автоматически расширяются, приближая нагретую кровь к поверхности, где она может охладиться. Дрожь — это еще одна реакция на озноб: непроизвольные движения сжигают ткани тела, выделяя больше тепла.
Отрицательная обратная связь возникает из-за баланса между силами и факторами, которые взаимно влияют друг на друга. Чтобы проиллюстрировать некоторые из его важных характеристик, мы можем рассматривать автомобиль и его водителя как единую, сложную, гомеостатическую или «целенаправленную» систему — киборга или «кибернетический организм» в том смысле, что они стремятся поддерживать движение автомобиля. на ходу.Водитель не управляет, удерживая колесо в фиксированном положении, а продолжает слегка поворачивать колесо влево и вправо, ища положения колес, которые вернут естественно извилистую машину на ход. Нарушение или отклонение от равновесия не менее важно, чем отрицательная обратная связь: системы не могут исправить себя, если они не отклоняются.
Колебания — обычное и необходимое поведение многих систем. Если автомобиль заносит, водитель автоматически реагирует, быстро поворачивая в противоположном направлении.Однако такая резкая отрицательная обратная связь обычно приводит к чрезмерной коррекции, заставляя автомобиль двигаться к другой стороне дороги.