Эмоции: понятие и функции
Эмоции являются процессом импульсивной регуляции поведения, который основан на отражении значимости внешних воздействий и их влияния на жизнедеятельность человека.
Эмоции появляются как реакция на явления окружающей среды, которые удовлетворяют или не удовлетворяют потребности человека. В случае, когда явление не соответствует потребностям, появляются отрицательные эмоции, и, соответственно, при соответствии потребности – положительные. Эмоции успеха (положительные) увеличивают стремление человека выполнить заданную цель, эмоции неуспеха (отрицательные) препятствуют, а иногда и дезорганизуют деятельность.
Эмоции энергетически мобилизуют организм. Например, радость приводит к усилению иннервации в мышцах, при этом мелкие артерии расширяются, увеличивается приток крови к коже, кожа становится теплее, ускоренное кровообращение делает легче питание тканей и улучшает физиологические процессы. Радость омолаживает, поскольку происходит оптимизация условий питания всех тканей тела.
Люди не могут жить без эмоций. Если жизнь человека чересчур однообразна, если он не получает сильные эмоции, даже если они отрицательные, человек начинает испытывать эмоциональный голод. В результате отсутствия сильных эмоций снижается результативность деятельности. К примеру, спортсмен, относящийся к тренировкам и выступлениям без эмоций, не может испытывать радость побед и горечь поражений. В итоге у него не будет никакого успеха в деятельности.
Для преодоления эмоционального голода, люди подсознательно стремятся к совершению рискованных действий. Все опасные поступки вызывают сильные эмоции. Для того, чтобы случился эмоциональный подъем и был получен адреналин, кто-то едет покорять горные вершины, кто-то несётся на огромной скорости по автостраде, отправляется в далёкие и опасные путешествия.
Итак, ежедневные сильные эмоции вредны для человека, но нельзя исключать сильные переживания. Эмоции занимают важное место в жизни человека.
- сигнальная;
- управляющая;
- защитная;
- мобилизующая;
- компенсаторная;
- дезорганизующая.
Сигнальная функция эмоций
Эмоциям свойственна субъективная окраска происходящего вокруг человека и в нём самом. Эмоции показывают окружающим внутреннее состояние: «не подходи, я злюсь», «обрати на меня внимание, у меня радость, которой хочу поделиться». На одинаковые события люди реагируют абсолютно по-разному, а эмоции могут помочь определить реакцию собеседника на ситуацию. Например, болельщики испытывают разочарование и огорчение от проигрыша их любимой команды, а болельщики команды соперника – радость. Эмоции помогают верно оценить эмоциональное состояние другого человека, прогнозировать развитие взаимоотношений с ним.
Управляющая функция эмоций
Эмоции нужны также, чтобы управлять поведением человека, они являются одним из психофизиологических механизмов этого управления. Это важно, поскольку появление определённого отношения к объекту оказывает влияние на мотивацию, на процесс принятия решения о действии или поступке, а физиологические изменения, которые сопровождают эмоции, влияют на качество деятельности, работоспособность человека. Эмоции оказывают мобилизующее воздействие на человека, при помощи эмоций возможно без труда переключаться на другой вид деятельности, сильные эмоции способны поддержать высокую работоспособность в течение длительного времени.
Защитная функция эмоций
Она связывается с возникновением страха. Страх является предупреждением о реальной или о мнимой угрозе, чем способствует лучшему продумыванию возникшей ситуации, более тщательному определению вероятности достижения успеха или неудачи. Таким образом, страх защищает нас от неприятных последствий и даже от гибели.
Академик П.К. Анохин считал, что эмоции нужны для закрепления, стабилизации рационального поведения животных и человека. Положительные эмоции, которые возникают при достижении цели, запоминаются и в нужной ситуации извлекаются из памяти с целью получения такого же полезного результата. Отрицательные эмоции, которые извлекаются из памяти, наоборот, предостерегают от повторного совершения ошибок. С точки зрения П.К. Анохина, эмоциональные переживания были закреплены в эволюции как механизмы, удерживающие жизненные процессы в оптимальных границах и предупреждающие разрушительный характер недостатка или избытка жизненно важных факторов.
Мобилизующая функция эмоций
Она состоит в том, что сильные эмоции позволяют организму мобилизовать все силы, даже те, которые глубоко зарезервированы. Страх способствует мобилизации резервов человека за счет выброса в кровь дополнительного количества адреналина, например, при спасении бегством. Способствуют мобилизации сил организма также и воодушевление, радость.
Компенсаторная функция эмоций
Она заключается в возмещении информации, которой недостаёт для принятия решения или вынесения суждения о чем-либо. Эмоция, которая возникает при столкновении с незнакомым объектом, придает объекту определённую окраску (плохой встретился человек или хороший), основываясь на его схожести с ранее встречавшимися объектами. Хотя при помощи эмоций человек выносит обобщенную и не всегда обоснованную оценку объекта и ситуации, они помогают ему выйти из тупика, когда он не знает, что ему делать в данной ситуации.
Существование отражательно-оценочной и компенсаторной функций позволяет быстро решить вопрос – идти на контакт с человеком или нет.
Дезорганизующая функция эмоций
Сильные отрицательные эмоции (гнев, страх, злость и т.п.) иногда нарушают поведение человека, препятствуют достижению целей. Например, злость может провоцировать человека повторять одни и те же действия, которые не приводят к успеху. Сильное волнение мешает сосредоточиться на задании, человек может забыть, что ему надо делать.
Решение задач
от 1 дня / от 150 р.
Курсовая работа
от 5 дней / от 1800 р.
Реферат
от 1 дня / от 700 р.
Эмоции: понятие и функции
Эмоции являются процессом импульсивной регуляции поведения, который основан на отражении значимости внешних воздействий и их влияния на жизнедеятельность человека.
Эмоции появляются как реакция на явления окружающей среды, которые удовлетворяют или не удовлетворяют потребности человека. В случае, когда явление не соответствует потребностям, появляются отрицательные эмоции, и, соответственно, при соответствии потребности – положительные. Эмоции успеха (положительные) увеличивают стремление человека выполнить заданную цель, эмоции неуспеха (отрицательные) препятствуют, а иногда и дезорганизуют деятельность.
Эмоции энергетически мобилизуют организм. Например, радость приводит к усилению иннервации в мышцах, при этом мелкие артерии расширяются, увеличивается приток крови к коже, кожа становится теплее, ускоренное кровообращение делает легче питание тканей и улучшает физиологические процессы. Радость омолаживает, поскольку происходит оптимизация условий питания всех тканей тела. С другой стороны, физиологическое проявление печали характеризуется парализующим влиянием на мышцы. Движения становятся медленными и слабыми, сосуды сужаются, ткани обескровливаются, появляется озноб, недостаток воздуха и тяжесть в груди. Огорчения делают человека старее, они изменяют кожу, волосы, ногти, зубы.
Радость омолаживает, поскольку происходит оптимизация условий питания всех тканей тела. С другой стороны, физиологическое проявление печали характеризуется парализующим влиянием на мышцы. Движения становятся медленными и слабыми, сосуды сужаются, ткани обескровливаются, появляется озноб, недостаток воздуха и тяжесть в груди. Огорчения делают человека старее, они изменяют кожу, волосы, ногти, зубы.
Люди не могут жить без эмоций. Если жизнь человека чересчур однообразна, если он не получает сильные эмоции, даже если они отрицательные, человек начинает испытывать эмоциональный голод. В результате отсутствия сильных эмоций снижается результативность деятельности. К примеру, спортсмен, относящийся к тренировкам и выступлениям без эмоций, не может испытывать радость побед и горечь поражений. В итоге у него не будет никакого успеха в деятельности.
Для преодоления эмоционального голода, люди подсознательно стремятся к совершению рискованных действий. Все опасные поступки вызывают сильные эмоции. Для того, чтобы случился эмоциональный подъем и был получен адреналин, кто-то едет покорять горные вершины, кто-то несётся на огромной скорости по автостраде, отправляется в далёкие и опасные путешествия. Когда человек стремится к удовлетворению потребности в сильных эмоциях, он создает для себя проблемные ситуации, которые потом у него не получается решить.
Для того, чтобы случился эмоциональный подъем и был получен адреналин, кто-то едет покорять горные вершины, кто-то несётся на огромной скорости по автостраде, отправляется в далёкие и опасные путешествия. Когда человек стремится к удовлетворению потребности в сильных эмоциях, он создает для себя проблемные ситуации, которые потом у него не получается решить.
Итак, ежедневные сильные эмоции вредны для человека, но нельзя исключать сильные переживания. Эмоции занимают важное место в жизни человека.
- сигнальная;
- управляющая;
- защитная;
- мобилизующая;
- компенсаторная;
- дезорганизующая.
Сигнальная функция эмоций
Эмоциям свойственна субъективная окраска происходящего вокруг человека и в нём самом. Эмоции показывают окружающим внутреннее состояние: «не подходи, я злюсь», «обрати на меня внимание, у меня радость, которой хочу поделиться». На одинаковые события люди реагируют абсолютно по-разному, а эмоции могут помочь определить реакцию собеседника на ситуацию. Например, болельщики испытывают разочарование и огорчение от проигрыша их любимой команды, а болельщики команды соперника – радость. Эмоции помогают верно оценить эмоциональное состояние другого человека, прогнозировать развитие взаимоотношений с ним.
Например, болельщики испытывают разочарование и огорчение от проигрыша их любимой команды, а болельщики команды соперника – радость. Эмоции помогают верно оценить эмоциональное состояние другого человека, прогнозировать развитие взаимоотношений с ним.
Управляющая функция эмоций
Эмоции нужны также, чтобы управлять поведением человека, они являются одним из психофизиологических механизмов этого управления. Это важно, поскольку появление определённого отношения к объекту оказывает влияние на мотивацию, на процесс принятия решения о действии или поступке, а физиологические изменения, которые сопровождают эмоции, влияют на качество деятельности, работоспособность человека. Эмоции оказывают мобилизующее воздействие на человека, при помощи эмоций возможно без труда переключаться на другой вид деятельности, сильные эмоции способны поддержать высокую работоспособность в течение длительного времени.
Защитная функция эмоций
Она связывается с возникновением страха. Страх является предупреждением о реальной или о мнимой угрозе, чем способствует лучшему продумыванию возникшей ситуации, более тщательному определению вероятности достижения успеха или неудачи. Таким образом, страх защищает нас от неприятных последствий и даже от гибели.
Таким образом, страх защищает нас от неприятных последствий и даже от гибели.
Академик П.К. Анохин считал, что эмоции нужны для закрепления, стабилизации рационального поведения животных и человека. Положительные эмоции, которые возникают при достижении цели, запоминаются и в нужной ситуации извлекаются из памяти с целью получения такого же полезного результата. Отрицательные эмоции, которые извлекаются из памяти, наоборот, предостерегают от повторного совершения ошибок. С точки зрения П.К. Анохина, эмоциональные переживания были закреплены в эволюции как механизмы, удерживающие жизненные процессы в оптимальных границах и предупреждающие разрушительный характер недостатка или избытка жизненно важных факторов.
Мобилизующая функция эмоций
Она состоит в том, что сильные эмоции позволяют организму мобилизовать все силы, даже те, которые глубоко зарезервированы. Страх способствует мобилизации резервов человека за счет выброса в кровь дополнительного количества адреналина, например, при спасении бегством. Способствуют мобилизации сил организма также и воодушевление, радость.
Способствуют мобилизации сил организма также и воодушевление, радость.
Компенсаторная функция эмоций
Она заключается в возмещении информации, которой недостаёт для принятия решения или вынесения суждения о чем-либо. Эмоция, которая возникает при столкновении с незнакомым объектом, придает объекту определённую окраску (плохой встретился человек или хороший), основываясь на его схожести с ранее встречавшимися объектами. Хотя при помощи эмоций человек выносит обобщенную и не всегда обоснованную оценку объекта и ситуации, они помогают ему выйти из тупика, когда он не знает, что ему делать в данной ситуации.
Существование отражательно-оценочной и компенсаторной функций позволяет быстро решить вопрос – идти на контакт с человеком или нет.
Дезорганизующая функция эмоций
Сильные отрицательные эмоции (гнев, страх, злость и т.п.) иногда нарушают поведение человека, препятствуют достижению целей. Например, злость может провоцировать человека повторять одни и те же действия, которые не приводят к успеху. Сильное волнение мешает сосредоточиться на задании, человек может забыть, что ему надо делать.
Сильное волнение мешает сосредоточиться на задании, человек может забыть, что ему надо делать.
Решение задач
от 1 дня / от 150 р.
Курсовая работа
от 5 дней / от 1800 р.
Реферат
от 1 дня / от 700 р.
Эмоции: понятие и функции
Эмоции являются процессом импульсивной регуляции поведения, который основан на отражении значимости внешних воздействий и их влияния на жизнедеятельность человека.
Эмоции появляются как реакция на явления окружающей среды, которые удовлетворяют или не удовлетворяют потребности человека. В случае, когда явление не соответствует потребностям, появляются отрицательные эмоции, и, соответственно, при соответствии потребности – положительные. Эмоции успеха (положительные) увеличивают стремление человека выполнить заданную цель, эмоции неуспеха (отрицательные) препятствуют, а иногда и дезорганизуют деятельность.
Эмоции энергетически мобилизуют организм. Например, радость приводит к усилению иннервации в мышцах, при этом мелкие артерии расширяются, увеличивается приток крови к коже, кожа становится теплее, ускоренное кровообращение делает легче питание тканей и улучшает физиологические процессы.
Люди не могут жить без эмоций. Если жизнь человека чересчур однообразна, если он не получает сильные эмоции, даже если они отрицательные, человек начинает испытывать эмоциональный голод. В результате отсутствия сильных эмоций снижается результативность деятельности. К примеру, спортсмен, относящийся к тренировкам и выступлениям без эмоций, не может испытывать радость побед и горечь поражений. В итоге у него не будет никакого успеха в деятельности.
Для преодоления эмоционального голода, люди подсознательно стремятся к совершению рискованных действий. Все опасные поступки вызывают сильные эмоции. Для того, чтобы случился эмоциональный подъем и был получен адреналин, кто-то едет покорять горные вершины, кто-то несётся на огромной скорости по автостраде, отправляется в далёкие и опасные путешествия. Когда человек стремится к удовлетворению потребности в сильных эмоциях, он создает для себя проблемные ситуации, которые потом у него не получается решить.
Для того, чтобы случился эмоциональный подъем и был получен адреналин, кто-то едет покорять горные вершины, кто-то несётся на огромной скорости по автостраде, отправляется в далёкие и опасные путешествия. Когда человек стремится к удовлетворению потребности в сильных эмоциях, он создает для себя проблемные ситуации, которые потом у него не получается решить.
Итак, ежедневные сильные эмоции вредны для человека, но нельзя исключать сильные переживания. Эмоции занимают важное место в жизни человека.
- сигнальная;
- управляющая;
- защитная;
- мобилизующая;
- компенсаторная;
- дезорганизующая.
Сигнальная функция эмоций
Эмоциям свойственна субъективная окраска происходящего вокруг человека и в нём самом. Эмоции показывают окружающим внутреннее состояние: «не подходи, я злюсь», «обрати на меня внимание, у меня радость, которой хочу поделиться». На одинаковые события люди реагируют абсолютно по-разному, а эмоции могут помочь определить реакцию собеседника на ситуацию. Например, болельщики испытывают разочарование и огорчение от проигрыша их любимой команды, а болельщики команды соперника – радость. Эмоции помогают верно оценить эмоциональное состояние другого человека, прогнозировать развитие взаимоотношений с ним.
Например, болельщики испытывают разочарование и огорчение от проигрыша их любимой команды, а болельщики команды соперника – радость. Эмоции помогают верно оценить эмоциональное состояние другого человека, прогнозировать развитие взаимоотношений с ним.
Управляющая функция эмоций
Эмоции нужны также, чтобы управлять поведением человека, они являются одним из психофизиологических механизмов этого управления. Это важно, поскольку появление определённого отношения к объекту оказывает влияние на мотивацию, на процесс принятия решения о действии или поступке, а физиологические изменения, которые сопровождают эмоции, влияют на качество деятельности, работоспособность человека. Эмоции оказывают мобилизующее воздействие на человека, при помощи эмоций возможно без труда переключаться на другой вид деятельности, сильные эмоции способны поддержать высокую работоспособность в течение длительного времени.
Защитная функция эмоций
Она связывается с возникновением страха. Страх является предупреждением о реальной или о мнимой угрозе, чем способствует лучшему продумыванию возникшей ситуации, более тщательному определению вероятности достижения успеха или неудачи. Таким образом, страх защищает нас от неприятных последствий и даже от гибели.
Таким образом, страх защищает нас от неприятных последствий и даже от гибели.
Академик П.К. Анохин считал, что эмоции нужны для закрепления, стабилизации рационального поведения животных и человека. Положительные эмоции, которые возникают при достижении цели, запоминаются и в нужной ситуации извлекаются из памяти с целью получения такого же полезного результата. Отрицательные эмоции, которые извлекаются из памяти, наоборот, предостерегают от повторного совершения ошибок. С точки зрения П.К. Анохина, эмоциональные переживания были закреплены в эволюции как механизмы, удерживающие жизненные процессы в оптимальных границах и предупреждающие разрушительный характер недостатка или избытка жизненно важных факторов.
Мобилизующая функция эмоций
Она состоит в том, что сильные эмоции позволяют организму мобилизовать все силы, даже те, которые глубоко зарезервированы. Страх способствует мобилизации резервов человека за счет выброса в кровь дополнительного количества адреналина, например, при спасении бегством. Способствуют мобилизации сил организма также и воодушевление, радость.
Способствуют мобилизации сил организма также и воодушевление, радость.
Компенсаторная функция эмоций
Она заключается в возмещении информации, которой недостаёт для принятия решения или вынесения суждения о чем-либо. Эмоция, которая возникает при столкновении с незнакомым объектом, придает объекту определённую окраску (плохой встретился человек или хороший), основываясь на его схожести с ранее встречавшимися объектами. Хотя при помощи эмоций человек выносит обобщенную и не всегда обоснованную оценку объекта и ситуации, они помогают ему выйти из тупика, когда он не знает, что ему делать в данной ситуации.
Существование отражательно-оценочной и компенсаторной функций позволяет быстро решить вопрос – идти на контакт с человеком или нет.
Дезорганизующая функция эмоций
Сильные отрицательные эмоции (гнев, страх, злость и т.п.) иногда нарушают поведение человека, препятствуют достижению целей. Например, злость может провоцировать человека повторять одни и те же действия, которые не приводят к успеху. Сильное волнение мешает сосредоточиться на задании, человек может забыть, что ему надо делать.
Сильное волнение мешает сосредоточиться на задании, человек может забыть, что ему надо делать.
Решение задач
от 1 дня / от 150 р.
Курсовая работа
от 5 дней / от 1800 р.
Реферат
от 1 дня / от 700 р.
Эмоции: понятие и функции
Эмоции являются процессом импульсивной регуляции поведения, который основан на отражении значимости внешних воздействий и их влияния на жизнедеятельность человека.
Эмоции появляются как реакция на явления окружающей среды, которые удовлетворяют или не удовлетворяют потребности человека. В случае, когда явление не соответствует потребностям, появляются отрицательные эмоции, и, соответственно, при соответствии потребности – положительные. Эмоции успеха (положительные) увеличивают стремление человека выполнить заданную цель, эмоции неуспеха (отрицательные) препятствуют, а иногда и дезорганизуют деятельность.
Эмоции энергетически мобилизуют организм. Например, радость приводит к усилению иннервации в мышцах, при этом мелкие артерии расширяются, увеличивается приток крови к коже, кожа становится теплее, ускоренное кровообращение делает легче питание тканей и улучшает физиологические процессы.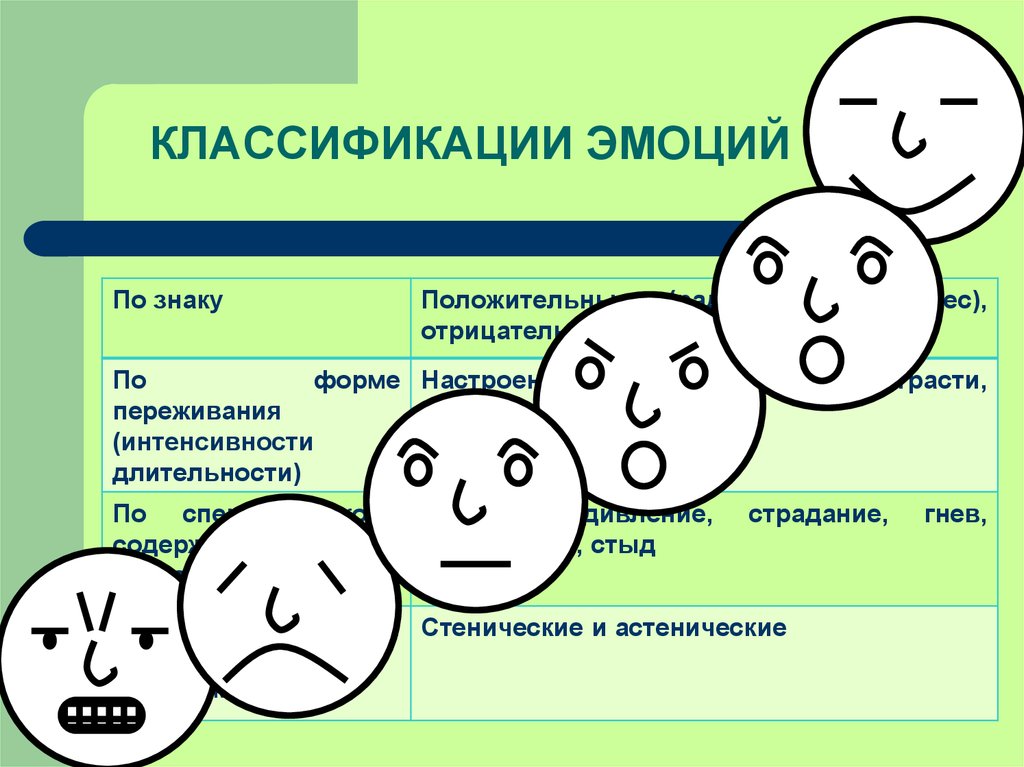 Радость омолаживает, поскольку происходит оптимизация условий питания всех тканей тела. С другой стороны, физиологическое проявление печали характеризуется парализующим влиянием на мышцы. Движения становятся медленными и слабыми, сосуды сужаются, ткани обескровливаются, появляется озноб, недостаток воздуха и тяжесть в груди. Огорчения делают человека старее, они изменяют кожу, волосы, ногти, зубы.
Радость омолаживает, поскольку происходит оптимизация условий питания всех тканей тела. С другой стороны, физиологическое проявление печали характеризуется парализующим влиянием на мышцы. Движения становятся медленными и слабыми, сосуды сужаются, ткани обескровливаются, появляется озноб, недостаток воздуха и тяжесть в груди. Огорчения делают человека старее, они изменяют кожу, волосы, ногти, зубы.
Люди не могут жить без эмоций. Если жизнь человека чересчур однообразна, если он не получает сильные эмоции, даже если они отрицательные, человек начинает испытывать эмоциональный голод. В результате отсутствия сильных эмоций снижается результативность деятельности. К примеру, спортсмен, относящийся к тренировкам и выступлениям без эмоций, не может испытывать радость побед и горечь поражений. В итоге у него не будет никакого успеха в деятельности.
Для преодоления эмоционального голода, люди подсознательно стремятся к совершению рискованных действий. Все опасные поступки вызывают сильные эмоции. Для того, чтобы случился эмоциональный подъем и был получен адреналин, кто-то едет покорять горные вершины, кто-то несётся на огромной скорости по автостраде, отправляется в далёкие и опасные путешествия. Когда человек стремится к удовлетворению потребности в сильных эмоциях, он создает для себя проблемные ситуации, которые потом у него не получается решить.
Для того, чтобы случился эмоциональный подъем и был получен адреналин, кто-то едет покорять горные вершины, кто-то несётся на огромной скорости по автостраде, отправляется в далёкие и опасные путешествия. Когда человек стремится к удовлетворению потребности в сильных эмоциях, он создает для себя проблемные ситуации, которые потом у него не получается решить.
Итак, ежедневные сильные эмоции вредны для человека, но нельзя исключать сильные переживания. Эмоции занимают важное место в жизни человека.
- сигнальная;
- управляющая;
- защитная;
- мобилизующая;
- компенсаторная;
- дезорганизующая.
Сигнальная функция эмоций
Эмоциям свойственна субъективная окраска происходящего вокруг человека и в нём самом. Эмоции показывают окружающим внутреннее состояние: «не подходи, я злюсь», «обрати на меня внимание, у меня радость, которой хочу поделиться». На одинаковые события люди реагируют абсолютно по-разному, а эмоции могут помочь определить реакцию собеседника на ситуацию. Например, болельщики испытывают разочарование и огорчение от проигрыша их любимой команды, а болельщики команды соперника – радость. Эмоции помогают верно оценить эмоциональное состояние другого человека, прогнозировать развитие взаимоотношений с ним.
Например, болельщики испытывают разочарование и огорчение от проигрыша их любимой команды, а болельщики команды соперника – радость. Эмоции помогают верно оценить эмоциональное состояние другого человека, прогнозировать развитие взаимоотношений с ним.
Управляющая функция эмоций
Эмоции нужны также, чтобы управлять поведением человека, они являются одним из психофизиологических механизмов этого управления. Это важно, поскольку появление определённого отношения к объекту оказывает влияние на мотивацию, на процесс принятия решения о действии или поступке, а физиологические изменения, которые сопровождают эмоции, влияют на качество деятельности, работоспособность человека. Эмоции оказывают мобилизующее воздействие на человека, при помощи эмоций возможно без труда переключаться на другой вид деятельности, сильные эмоции способны поддержать высокую работоспособность в течение длительного времени.
Защитная функция эмоций
Она связывается с возникновением страха. Страх является предупреждением о реальной или о мнимой угрозе, чем способствует лучшему продумыванию возникшей ситуации, более тщательному определению вероятности достижения успеха или неудачи. Таким образом, страх защищает нас от неприятных последствий и даже от гибели.
Таким образом, страх защищает нас от неприятных последствий и даже от гибели.
Академик П.К. Анохин считал, что эмоции нужны для закрепления, стабилизации рационального поведения животных и человека. Положительные эмоции, которые возникают при достижении цели, запоминаются и в нужной ситуации извлекаются из памяти с целью получения такого же полезного результата. Отрицательные эмоции, которые извлекаются из памяти, наоборот, предостерегают от повторного совершения ошибок. С точки зрения П.К. Анохина, эмоциональные переживания были закреплены в эволюции как механизмы, удерживающие жизненные процессы в оптимальных границах и предупреждающие разрушительный характер недостатка или избытка жизненно важных факторов.
Мобилизующая функция эмоций
Она состоит в том, что сильные эмоции позволяют организму мобилизовать все силы, даже те, которые глубоко зарезервированы. Страх способствует мобилизации резервов человека за счет выброса в кровь дополнительного количества адреналина, например, при спасении бегством. Способствуют мобилизации сил организма также и воодушевление, радость.
Способствуют мобилизации сил организма также и воодушевление, радость.
Компенсаторная функция эмоций
Она заключается в возмещении информации, которой недостаёт для принятия решения или вынесения суждения о чем-либо. Эмоция, которая возникает при столкновении с незнакомым объектом, придает объекту определённую окраску (плохой встретился человек или хороший), основываясь на его схожести с ранее встречавшимися объектами. Хотя при помощи эмоций человек выносит обобщенную и не всегда обоснованную оценку объекта и ситуации, они помогают ему выйти из тупика, когда он не знает, что ему делать в данной ситуации.
Существование отражательно-оценочной и компенсаторной функций позволяет быстро решить вопрос – идти на контакт с человеком или нет.
Дезорганизующая функция эмоций
Сильные отрицательные эмоции (гнев, страх, злость и т.п.) иногда нарушают поведение человека, препятствуют достижению целей. Например, злость может провоцировать человека повторять одни и те же действия, которые не приводят к успеху.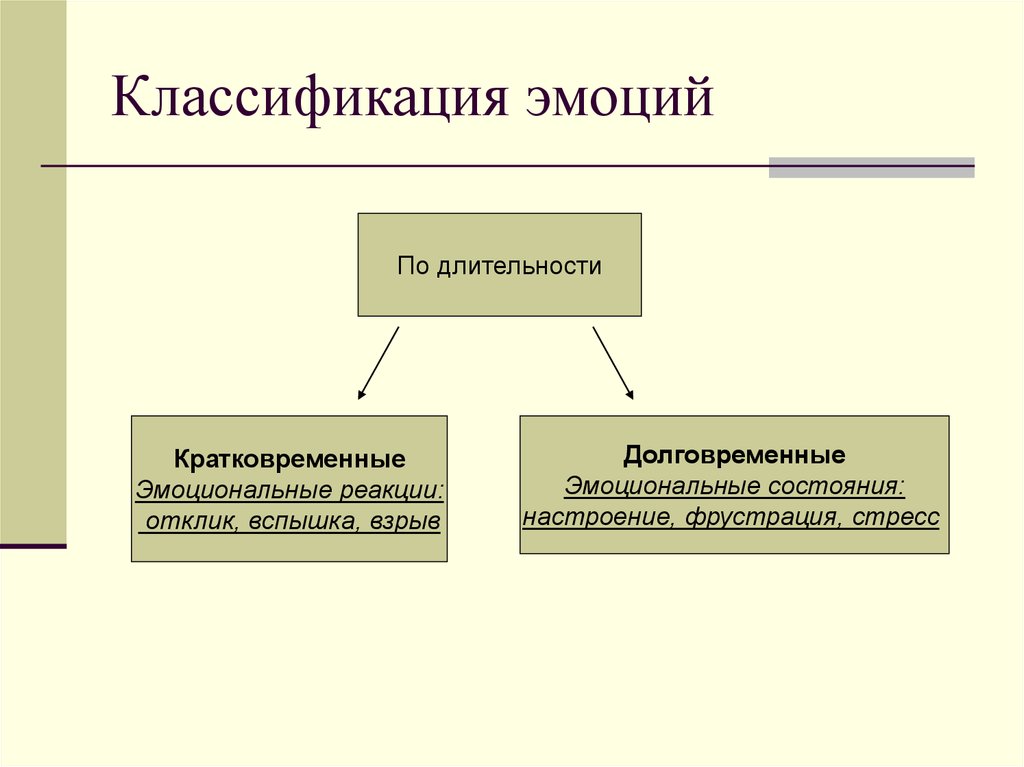 Сильное волнение мешает сосредоточиться на задании, человек может забыть, что ему надо делать.
Сильное волнение мешает сосредоточиться на задании, человек может забыть, что ему надо делать.
Решение задач
от 1 дня / от 150 р.
Курсовая работа
от 5 дней / от 1800 р.
Реферат
от 1 дня / от 700 р.
Функции и виды эмоций
За эмоциями была признана важная положительная роль в жизни людей, и с ними стали связывать следующие положительные функции: Мотивационно-регулирующую, коммуникативную, сигнальную и защитную.
Мотивационно-регулирующая функциязаключается в том, что эмоции участвуют в мотивации поведения человека, могут побуждать, направлять и регулировать. Иногда эмоции могут заменять собой мышление в регуляции поведения.
Коммуникативная функция заключается в том, что эмоции, точнее, способы их внешнего выражения, несут в себе информацию о психическом и физическом состоянии человека. Благодаря эмоциям мы лучше понимаем друг друга. Наблюдая за изменениями эмоциональных состояний, появляется возможность судить о том, что происходит в психике. Комментарий: люди, принадлежащие к разным культурам, способны безошибочно воспринимать и оценивать многие выражения человеческого лица, определять по нему такие эмоции, как радость, гнев, печаль, страх, отвращение, удивление. Это относится и к тем народам, которые вообще никогда не находились в прямых контактах друг с другом.
Это относится и к тем народам, которые вообще никогда не находились в прямых контактах друг с другом.
Сигнальная функция. Жизнь без эмоций так же невозможна, как и без ощущений. Эмоции, утверждал Ч. Дарвин, возникли в процессе эволюции как средство, при помощи которого живые существа устанавливают значимость тех или иных условий для удовлетворения актуальных для них потребностей. Эмоционально-выразительные движения (мимика, жесты, пантомимика) выполняют функцию сигналов о том, в каком состоянии находится система потребностей человека.
Защитная функция выражается в том, что, возникая как моментальная, быстрая реакция организма, может защитить человека от опасностей.
Установлено, что чем сложнее организовано живое существо, чем более высокую ступень на эволюционной лестнице оно занимает, тем богаче и разнообразнее гамма эмоций, которую оно способно переживать.
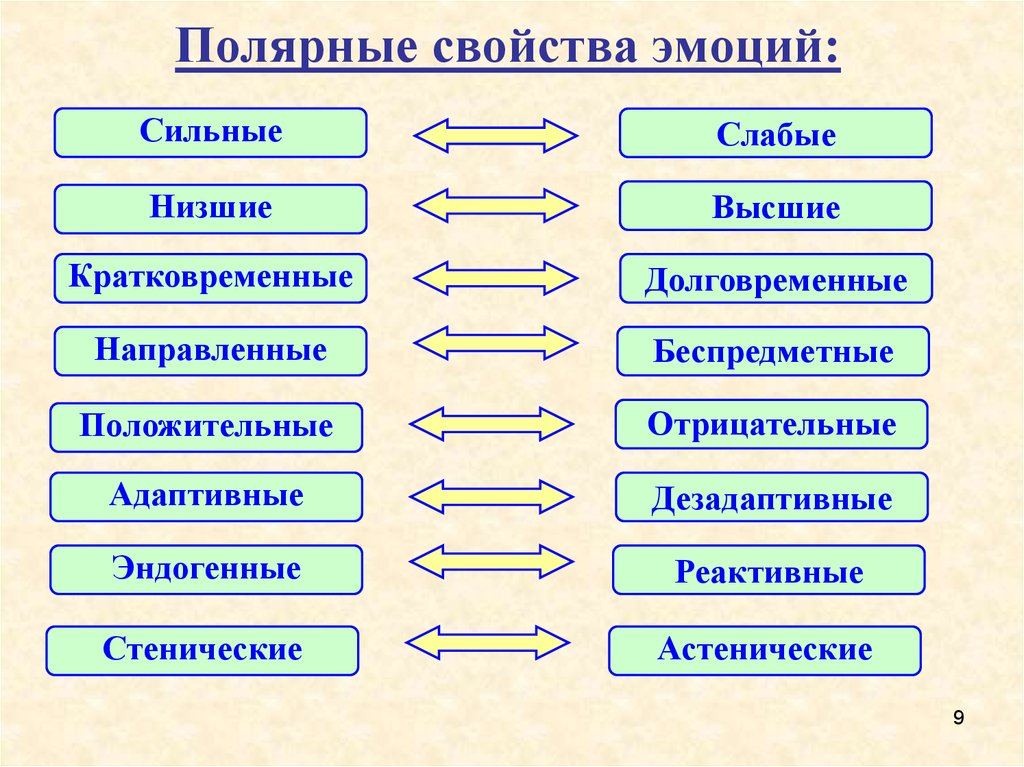
Характер переживания (удовольствие или неудовольствие) определяет знак эмоций — положительные и отрицательные. С точки зрения влияния на деятельность человека эмоции делятся на стенические и астенические. Стенические эмоции стимулируют деятельность, увеличивают энергию и напряжение сил человека, побуждают его к поступкам, высказываниям. Крылатое выражение: “готов горы свернуть”. И, наоборот, иногда переживания характеризуются своеобразной скованностью, пассивностью, тогда говорят об астенических эмоциях. Поэтому в зависимости от ситуации и индивидуальных особенностей эмоции могут по-разному влиять на поведение. Так, горе может вызвать апатию, бездеятельность у слабого человека, в то время как сильный человек удваивает свою энергию, находя утешение в работе и творчестве.
С точки зрения влияния на деятельность человека эмоции делятся на стенические и астенические. Стенические эмоции стимулируют деятельность, увеличивают энергию и напряжение сил человека, побуждают его к поступкам, высказываниям. Крылатое выражение: “готов горы свернуть”. И, наоборот, иногда переживания характеризуются своеобразной скованностью, пассивностью, тогда говорят об астенических эмоциях. Поэтому в зависимости от ситуации и индивидуальных особенностей эмоции могут по-разному влиять на поведение. Так, горе может вызвать апатию, бездеятельность у слабого человека, в то время как сильный человек удваивает свою энергию, находя утешение в работе и творчестве.
Модальность — основная качественная характеристика эмоций, определяющая их вид по специфике и особой окрашенности переживаний. По модальности выделяются три базовые эмоции: страх, гнев и радость. При всем многообразии практически любая эмоция является своеобразным выражением одной из этих эмоций. Тревожность, беспокойство, боязнь, ужас представляют собой различные проявления страха; злоба, раздражительность, ярость — гнева; веселье, ликование, торжество — радости.
К. Изард выделил следующие основные эмоции
Интерес (как эмоция) — положительное эмоциональное состояние, способствующее развитию навыков и умений, приобретению знаний.
Радость — положительное эмоциональное состояние, связанное с возможностью достаточно полно удовлетворить актуальную потребность, вероятность чего до этого момента была невелика или, во всяком случае, неопределенна.
Удивление — не имеющая четко выраженного положительного или отрицательного знака эмоциональная реакция на внезапно возникшие обстоятельства. Удивление тормозит все предыдущие эмоции, направляя внимание на объект, его вызвавший, и может переходить в интерес.
Страдание — отрицательное эмоциональное состояние, связанное с полученной достоверной или кажущейся таковой информацией о невозможности удовлетворения важнейших жизненных потребностей, которое до этого момента представлялось более или менее вероятным, чаще всего протекает в форме эмоционального стресса.
Гнев — эмоциональное состояние, отрицательное по знаку, как правило, протекающее в форме аффекта и вызываемое внезапным возникновением серьезного препятствия на пути удовлетворения исключительно важной для субъекта потребности.
Отвращение — отрицательное эмоциональное состояние, вызываемое объектами (предметами, людьми, обстоятельствами), соприкосновение с которыми (физическое взаимодействие, коммуникация в общении и пр.) вступает в резкое противоречие с идеологическими, нравственными или эстетическими принципами и установками субъекта. Отвращение, если оно сочетается с гневом, может в межличностных отношениях мотивировать агрессивное поведение, где нападение мотивируется гневом, а отвращение — желанием избавиться от кого-либо или чего-либо.
Внимание!
Если вам нужна помощь в написании работы, то рекомендуем обратиться к
профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные
корректировки и доработки. Узнайте стоимость своей работы.
Узнайте стоимость своей работы.
Расчет стоимостиГарантииОтзывы
Презрение — отрицательное эмоциональное состояние, возникающее в межличностных взаимоотношениях и порождаемое рассогласованием жизненных позиций, взглядов и поведения субъекта с жизненными позициями, взглядами и поведением объекта чувства. Последние представляются субъекту как низменные, не соответствующие принятым нравственным нормам и эстетическим критериям.
Страх — отрицательное эмоциональное состояние, появляющееся при получении субъектом информации о возможной угрозе его жизненному благополучию, о реальной или воображаемой опасности. В отличие от эмоции страдания, вызываемой прямым блокированием важнейших потребностей, человек, переживая эмоцию страха, располагает лишь вероятностным прогнозом возможного неблагополучия и действует на основе этого (часто недостаточно достоверного или преувеличенного прогноза).
Стыд — отрицательное состояние, выражающееся в осознании несоответствия собственных помыслом, поступков и внешности не только ожиданиям окружающих, но и собственным представлениям о подобающем поведении и внешнем облике.
Эмоции характеризуются также силой, продолжительностью и осознанием. Диапазон различий по силе внутреннего переживания и внешних проявлений очень велик для эмоции любой модальности. Радость может проявляться как слабая по силе эмоция, например, когда человек испытывает чувство удовлетворения. Восторг — эмоция большей силы. Гнев проявляется в диапазоне от раздражительности и негодования до ненависти и ярости, страх — от легкого беспокойства до ужаса. По продолжительности эмоции длятся от нескольких секунд до многих лет. Степень осознанности эмоций также может быть различной. Порой человеку трудно понять, какую эмоцию он испытывает и почему она возникает.
Эмоциональные переживания носят неоднозначный характер. Один и тот же объект может вызвать несогласованные, противоречивые эмоции. Это явление получило название амбивалентность (двойственность) чувств. Например, можно уважать кого-то за работоспособность и одновременно осуждать за вспыльчивость.
Качества, характеризующие каждую конкретную эмоциональную реакцию, могут сочетаться различным образом, что создает многоликие формы их выражения. Основные формы проявления эмоций — чувственный тон, ситуативная эмоция, аффект, страсть, стресс, настроение и чувство.
Основные формы проявления эмоций — чувственный тон, ситуативная эмоция, аффект, страсть, стресс, настроение и чувство.
Чувственный тон выражается в том, что многие ощущения человека имеют свою эмоциональную окраску. То есть люди, не просто ощущают какой-либо запах или вкус, а воспринимают его как приятный или неприятный. Образы восприятия, памяти, мышления, воображения также эмоционально окрашены. А. Н. Леонтьев считал одним из существенных качеств человеческого познания феномен, который называл “пристрастностью” отражения мира.
Ситуативные эмоции возникают в процессе жизнедеятельности человека чаще всех других эмоциональных реакций. Их главными характеристиками считают относительно малую силу, кратковременность, быструю смену эмоций, малую внешнюю наглядность.
Нравственные чувства отражают отношение человека к требованиям морали. Система нравственных чувств российского человека состоит из чувства справедливости, чести, долга, ответственности, патриотизма, солидарности. Нравственные чувства тесно связаны с мировоззрением человека, его убеждениями, помыслами, принципами поведения.
Нравственные чувства тесно связаны с мировоззрением человека, его убеждениями, помыслами, принципами поведения.
В общении следователя и судьи с участниками уголовного процесса эмоции и чувства играют особую роль. В частности, без них было бы невозможно достижение контакта с допрашиваемыми, в особенности в конфликтных ситуациях. Однако, являясь необходимыми и неизбежными, эмоции и чувства в расследовании и рассмотрении уголовного дела играют не только положительную роль. Отрицательные эмоциональные состояния следователя и судьи могут нарушать, а иногда и разрушать их деятельность. Эмоции и чувства способны вызывать представления, не соответствующие действительности. Под влиянием чрезмерных чувств следователь или судья могут иногда прийти к результатам, лишь желаемым, но далеким от истинного положения дел.
Поможем написать любую работу на аналогичную тему
Реферат
Функции и виды эмоций
От 250 руб
Контрольная работа
Функции и виды эмоций
От 250 руб
Курсовая работа
Функции и виды эмоций
От 700 руб
Получить выполненную работу или консультацию специалиста по вашему учебному проекту
Узнать стоимость
Функции эмоций — Предмет «Общая психология».
 Лекции
ЛекцииОценочная (отражательная, сигнальная) функция эмоций выражается в обобщенной оценке событий. Она заключается в оценке значимости для субъекта той или иной ситуации или объектов в их отношении к его мотивационной сфере. В форме эмоции субъекту предстает степень настоятельности потребности, в том числе и степень ее удовлетворения к настоящему моменту, в наличной ситуации, а также вероятность ее удовлетворения в ожидаемой ситуации.
Примером может служить поведение человека, получившего травму конечности. Ориентируясь на боль, человек немедленно находит такое ее положение, которое уменьшает болевые ощущения.
Подкрепляюще-побуждающая
Подкрепляющая функция заключается в том, что после достижения ситуации, получения предмета или выполнения действия, способных удовлетворить потребность, возникает состояние, в ряде случаев очень сильное, как награда за достижение цели, которая в свою очередь в дальнейшем становится силой, побуждающей к достижению подобных ситуаций.
Эта схема верна как в отношении положительных, так и отрицательных эмоций, которые «наказывают» человека за осуществление или неосуществление определенных действий и в дальнейшем побуждают его избегать ситуаций, наносящих вред организму или личности, т.е. не способствуют, а препятствуют удовлетворению потребностей.
Т.о. «+» эмоции определяют к чему нужно стремится,
«−»эмоции определяют чего нужно избегать.
Подкрепляющая функция эмоций наиболее успешно была исследована на экспериментальной модели «эмоционального резонанса», предложенной П.В. Симоновым. Было обнаружено, что эмоциональные реакции одних животных могут возникать под влиянием отрицательных эмоциональных состояний других особей, подвергнутых воздействию электрокожного раздражения. Эта модель воспроизводит типичную для социальных взаимоотношений ситуацию возникновения отрицательных эмоциональных состояний в сообществе и позволяет изучать функции эмоций в наиболее чистом виде без непосредственного действия болевых раздражителей. В опытах Л.А.Преображенской, в которых собака-«жертва» подвергалась наказанию электрически током на глазах у собаки-«наблюдателя», у последней возрастала частота сердцебиения и увеличивалась синхронизация гиппокампального тета-ритма. Это указывает на появление у нее негативного эмоционального напряжения. В таких условиях собака-«наблюдатель» способна выработать избегательный инструментальный рефлекс (в виде подъема лапы), прекращающий подачу тока собаке-«жертве». Выработка такого инструментального рефлекса у собаки-«наблюдателя» сопровождается снижением у нее ЧСС и уменьшением гиппокампального тета-ритма, т.е. исчезновением отрицательного эмоционального состояния. Следовательно, предотвращение отрицательного эмоционального напряжения и служит ей той наградой, на которой и вырабатывается данный условный инструментальный рефлекс.
В опытах Л.А.Преображенской, в которых собака-«жертва» подвергалась наказанию электрически током на глазах у собаки-«наблюдателя», у последней возрастала частота сердцебиения и увеличивалась синхронизация гиппокампального тета-ритма. Это указывает на появление у нее негативного эмоционального напряжения. В таких условиях собака-«наблюдатель» способна выработать избегательный инструментальный рефлекс (в виде подъема лапы), прекращающий подачу тока собаке-«жертве». Выработка такого инструментального рефлекса у собаки-«наблюдателя» сопровождается снижением у нее ЧСС и уменьшением гиппокампального тета-ритма, т.е. исчезновением отрицательного эмоционального состояния. Следовательно, предотвращение отрицательного эмоционального напряжения и служит ей той наградой, на которой и вырабатывается данный условный инструментальный рефлекс.
Три правила наказания
1) постоянство и понятность требований
2) постоянство в применении наказания (или вообще не наказывать)
3) знание о возможности избежать наказания в случае правильного поведения.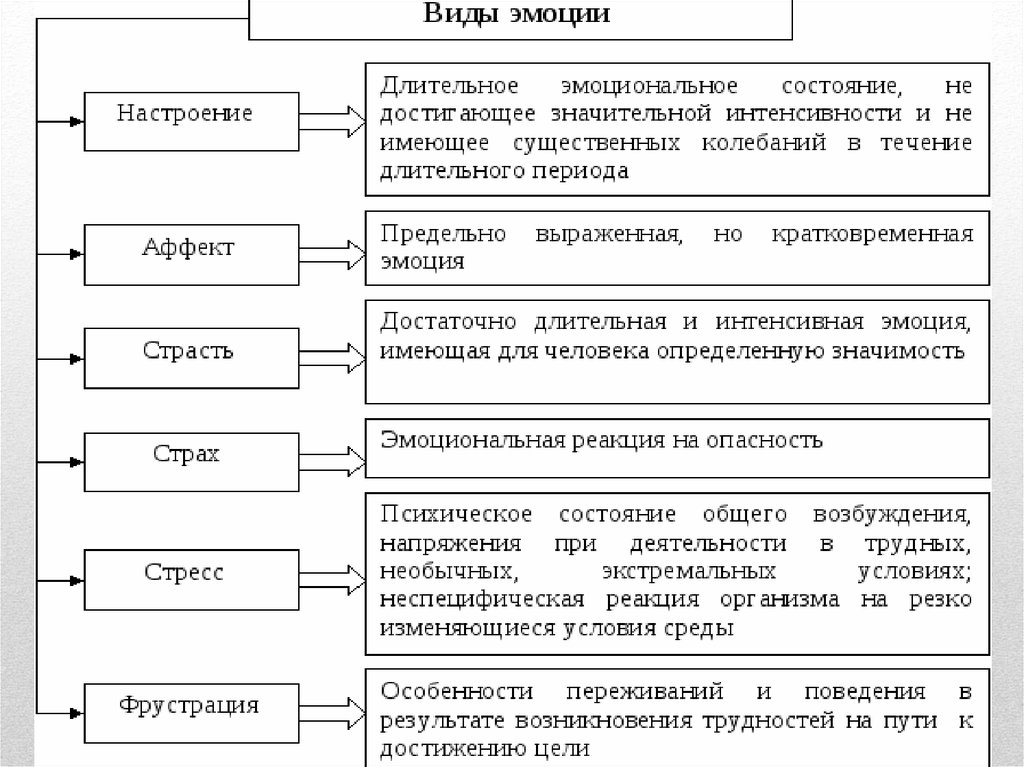
Функция побуждения заключается в том, что эмоции не только сигнализируют о потребности или желании, но и с достижением ими определенной интенсивности они настоятельно побуждают субъекта их удовлетворить.
Нехватка или несовместимость чего-то с внутренними условиями существования организма или личности вызывает эмоциональные переживания, иногда очень сильные, побуждающие человека действовать. Это и испуг, и гнев, и зависть, и ненависть, и чувство вины, и любовь, и множество других оттенков эмоциональных переживаний.
Коммуникативная — для каждой эмоции характерен свой комплекс невербальных проявлений, по этим невербальным проявлениям мы узнаем о том, что чувствует человек, получаем представление о его мыслях и намерениях.
Пример улыбка бывает радостная, печальная, презрительная, злорадная и т.д.
Внимание!
Если вам нужна помощь в написании работы, то рекомендуем обратиться к
профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные
корректировки и доработки. Узнайте стоимость своей работы.
Бесплатные
корректировки и доработки. Узнайте стоимость своей работы.
Расчет стоимостиГарантииОтзывы
Вопрос имеет ли традиция приветствовать друг друга при встрече какое-либо рациональное объяснение?
Мобилизирующая — в критических ситуациях эмоции запускают аварийный энергетический режим. (убегая от собаки человек способен перепрыгнуть ч/з 2-х метровый забор
Поможем написать любую работу на аналогичную тему
Реферат
Функции эмоций
От 250 руб
Контрольная работа
Функции эмоций
От 250 руб
Курсовая работа
Функции эмоций
От 700 руб
Получить выполненную работу или консультацию специалиста по вашему учебному проекту
Узнать стоимость
Значение социального сигнала эмоций
. 2012;26(3):385-9.
2012;26(3):385-9.
дои: 10.1080/02699931.2012.665029.
Шломо Харели 1 , Урсула Хесс
принадлежность
- 1 Высшая школа менеджмента Хайфского университета, Хайфа, Израиль. [email protected]
- PMID: 22471847
- DOI: 10.1080/02699931.2012.665029
Шломо Харели и др. Познание эмоций. 2012.
. 2012;26(3):385-9.
дои: 10. 1080/02699931.2012.665029.
1080/02699931.2012.665029.
Авторы
Шломо Харели 1 , Урсула Хесс
принадлежность
- 1 Высшая школа менеджмента Хайфского университета, Хайфа, Израиль. [email protected]
- PMID: 22471847
- DOI: 10.1080/02699931.2012.665029
Абстрактный
Человеческие взаимодействия изобилуют эмоциональными обменами. В этих обменах информация об эмоциональном состоянии партнеров по взаимодействию является только одним из видов передаваемой информации. Кроме того, дисплеи эмоций предоставляют информацию о расположении партнеров по взаимодействию и ситуации как таковой. То есть эмоции служат социальными сигналами. Признавая эту роль эмоций, в этом специальном разделе собраны исследования, которые иллюстрируют, как восприятие человека и понимание ситуации могут быть получены из эмоциональных проявлений и модуляции этого процесса через контекст. Три вклада сосредоточены на информации о выразителях и их намерениях. В дополнительной статье основное внимание уделяется информативной ценности эмоциональных выражений для интерпретации социальных ситуаций наблюдателем, а в другой статье приводится пример того, как контекст определяет социальное воздействие эмоций. Наконец, в последней статье представлена динамическая природа взаимного влияния эмоций. В попытке объединить эти вклады и предложить линзы для будущих исследований, эта редакционная статья предлагает контекстуализированную модель социального восприятия, которая пытается систематизировать не только типы информации, которую могут передавать выражения эмоций, но и разработать понятие контекста.
То есть эмоции служат социальными сигналами. Признавая эту роль эмоций, в этом специальном разделе собраны исследования, которые иллюстрируют, как восприятие человека и понимание ситуации могут быть получены из эмоциональных проявлений и модуляции этого процесса через контекст. Три вклада сосредоточены на информации о выразителях и их намерениях. В дополнительной статье основное внимание уделяется информативной ценности эмоциональных выражений для интерпретации социальных ситуаций наблюдателем, а в другой статье приводится пример того, как контекст определяет социальное воздействие эмоций. Наконец, в последней статье представлена динамическая природа взаимного влияния эмоций. В попытке объединить эти вклады и предложить линзы для будущих исследований, эта редакционная статья предлагает контекстуализированную модель социального восприятия, которая пытается систематизировать не только типы информации, которую могут передавать выражения эмоций, но и разработать понятие контекста.
Похожие статьи
Социальное восприятие у детей с ограниченными интеллектуальными возможностями: интерпретация доброжелательных и враждебных намерений.

Лефферт Дж.С., Сиперштейн Г.Н., Видаман К.Ф. Лефферт Дж. С. и соавт. J Intellect Disabil Res. 2010 февраль; 54 (2): 168-80. doi: 10.1111/j.1365-2788.2009.01240.x. Epub 2010 12 января. J Intellect Disabil Res. 2010. PMID: 20070474
Эмоции и толкование социальных ситуаций: выводы о сотрудничестве и конкуренции на основе выражений гнева, счастья и разочарования.
Ван Дорн Э.А., Хердинк М.В., Ван Клиф Г.А. Ван Дорн Э.А. и др. Познание эмоций. 2012;26(3):442-61. дои: 10.1080/02699931.2011.648174. Познание эмоций. 2012. PMID: 22471851
Роль эмоций в развитии и организации личности.
Малатеста Чехия. Малатеста Чехия. Небр Симп Мотив. 1988; 36:1-56. Небр Симп Мотив. 1988 год.
PMID: 3078934
Обзор.Эмоции как сигналы нормативного поведения.
Харели С., Моран-Амир О., Дэвид С., Хесс У. Харели С. и др. Познание эмоций. 2013;27(8):1395-404. дои: 10.1080/02699931.2013.7. Epub 2013 7 мая. Познание эмоций. 2013. PMID: 23650979
Социальная функциональность человеческих эмоций.
Ниденталь П.М., Брауэр М. Ниденталь П.М. и др. Анну Рев Психол. 2012;63:259-85. doi: 10.1146/annurev.psych.121208.131605. Epub 2011 10 октября. Анну Рев Психол. 2012. PMID: 22017377 Обзор.

 PMID: 3078934
Обзор.
PMID: 3078934
Обзор.Посмотреть все похожие статьи
Цитируется
Модуляция боли вашим партнером: экспериментальное исследование с социально-аффективной точки зрения.
Хиллмер К., Каппессер Дж., Герман К. Хиллмер К. и др. ПЛОС Один. 2021 22 июля; 16 (7): e0254069. doi: 10.1371/journal.pone.0254069. Электронная коллекция 2021. ПЛОС Один. 2021. PMID: 342
Бесплатная статья ЧВК.
Клиническое испытание.Динамические эмоциональные выражения не модулируют реакции на жесты.
Фармер Х., Махмуд Р., Грегори СЭА, Тишина П., Гамильтон АФК. Фармер Х. и др. Acta Psychol (Амст). 2021 янв;212:103226. doi: 10.1016/j.actpsy.2020.103226. Epub 2020 10 декабря. Acta Psychol (Амст). 2021. PMID: 33310344 Бесплатная статья ЧВК.
Правое височно-теменное соединение модулирует внутригрупповую предвзятость в эмоциональной мимикрии лица: исследование tDCS.
Пэн С.
, Куанг Б., Ху П.
Пэн С. и др.
Фронт Behav Neurosci. 2020 28 авг; 14:143. дои: 10.3389/fnbeh.2020.00143. Электронная коллекция 2020.
Фронт Behav Neurosci. 2020.
PMID: 33061895
Бесплатная статья ЧВК.Взаимосвязь между регулированием эмоций и знанием эмоций у дошкольников: лонгитюдное исследование.
Лукас-Молина Б., Кинтанилья Л., Сарменто-Энрике Р., Мартин Бабарро Х., Хименес-Даси М. Лукас-Молина Б. и соавт. Общественное здравоохранение Int J Environ Res. 2020 7 августа; 17 (16): 5726. дои: 10.3390/ijerph27165726. Общественное здравоохранение Int J Environ Res. 2020. PMID: 32784765 Бесплатная статья ЧВК.
Врачи испытывают сильные эмоции, видя своих пациентов: что происходит?
Сильва СП, Карвалью И.
Сильва СП и др.
Пермь Ж. Лето 2016;20(3):15-229. doi: 10.7812/TPP/15-229. Epub 2016 29 июля.
Пермь Ж. 2016.
PMID: 27479947
Бесплатная статья ЧВК.
 , Куанг Б., Ху П.
Пэн С. и др.
Фронт Behav Neurosci. 2020 28 авг; 14:143. дои: 10.3389/fnbeh.2020.00143. Электронная коллекция 2020.
Фронт Behav Neurosci. 2020.
PMID: 33061895
Бесплатная статья ЧВК.
, Куанг Б., Ху П.
Пэн С. и др.
Фронт Behav Neurosci. 2020 28 авг; 14:143. дои: 10.3389/fnbeh.2020.00143. Электронная коллекция 2020.
Фронт Behav Neurosci. 2020.
PMID: 33061895
Бесплатная статья ЧВК. Сильва СП и др.
Пермь Ж. Лето 2016;20(3):15-229. doi: 10.7812/TPP/15-229. Epub 2016 29 июля.
Пермь Ж. 2016.
PMID: 27479947
Бесплатная статья ЧВК.
Сильва СП и др.
Пермь Ж. Лето 2016;20(3):15-229. doi: 10.7812/TPP/15-229. Epub 2016 29 июля.
Пермь Ж. 2016.
PMID: 27479947
Бесплатная статья ЧВК.Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
Эмоциональные сигналы от лиц, тел и сцен влияют на выражение лиц, фиксации и размер зрачков наблюдателей
Введение
Представьте, что к вам приближается человек, сжимая кулаки, его мышцы напряжены. Такой эмоциональный сигнал иначе воспринимается в контексте спортивного события, чем на узкой улице посреди ночи. Однако в описанной выше ситуации, скорее всего, сразу реагировать , а не активно навешивать ярлыки на эмоциональное выражение мужчины. Распознаванию выражений лица уделяется много внимания в литературе по эмоциям (Haxby et al., 2000; Adolphs, 2002). Более поздние исследования показали, что на наше распознавание выражения лица влияет выражение тела (Meeren et al., 2005; Van den Stock et al., 2007; Kret and de Gelder, 2013; Kret et al., 2013) и окружающей сценой, то есть контекстом (Righart and de Gelder, 2006, 2008a,b; Kret and de Gelder, 2012a). Целью настоящего исследования является изучение того, как воспринимается наличие нескольких эмоциональных сигналов, состоящих из одновременно представленных выражений лица и тела или выражения тела, расположенного в эмоциональной сцене, путем исследования физиологических коррелятов в естественной ситуации пассивного просмотра. .
Более поздние исследования показали, что на наше распознавание выражения лица влияет выражение тела (Meeren et al., 2005; Van den Stock et al., 2007; Kret and de Gelder, 2013; Kret et al., 2013) и окружающей сценой, то есть контекстом (Righart and de Gelder, 2006, 2008a,b; Kret and de Gelder, 2012a). Целью настоящего исследования является изучение того, как воспринимается наличие нескольких эмоциональных сигналов, состоящих из одновременно представленных выражений лица и тела или выражения тела, расположенного в эмоциональной сцене, путем исследования физиологических коррелятов в естественной ситуации пассивного просмотра. .
Когда мы наблюдаем эмоциональное состояние другого человека, инициируются различные процессы. Во-первых, наше внимание привлекает лицо (Грин и др., 2003; Лундквист и Оман, 2005; Фокс и Дамьянович, 2006) и тело (Бандеттини и др., 1992), поскольку они содержат наиболее важную информацию и обычно дополняют друг друга. друг друга. Затем мы тоже возбуждаемся: сердцебиение меняется, мы начинаем потеть, а зрачки расширяются (Bradley et al. , 2008). Более того, вполне вероятно, что наблюдаемая эмоция отражается на нашем собственном лице (Димберг, 19 лет).82; Гесс и Фишер, 2013). Таким образом, до сих пор эти физиологические исследования в основном рассматривали восприятие изолированных выражений эмоций на лице, а вовсе не влияние контекста, такого как поза тела. Изучение восприятия смешанных сообщений с этих разных точек зрения будет способствовать модификации существующих моделей, которые пытаются предсказать восприятие неконгруэнтных эмоциональных контекстных сигналов, но пока безуспешно (Mondloch et al., 2013). Настоящее исследование направлено на изучение двух вопросов:
, 2008). Более того, вполне вероятно, что наблюдаемая эмоция отражается на нашем собственном лице (Димберг, 19 лет).82; Гесс и Фишер, 2013). Таким образом, до сих пор эти физиологические исследования в основном рассматривали восприятие изолированных выражений эмоций на лице, а вовсе не влияние контекста, такого как поза тела. Изучение восприятия смешанных сообщений с этих разных точек зрения будет способствовать модификации существующих моделей, которые пытаются предсказать восприятие неконгруэнтных эмоциональных контекстных сигналов, но пока безуспешно (Mondloch et al., 2013). Настоящее исследование направлено на изучение двух вопросов:
- Как обрабатываются выражения лица и тела при одновременном представлении? На лицо смотрят по-разному в зависимости от выражения тела и наоборот? Изменится ли выражение лица и уровень возбуждения участника в зависимости от различных эмоциональных сигналов, которые он наблюдает на лице и теле?
- Как обрабатываются выражения тела, когда они представлены в социально-эмоциональном контексте? Будет ли центральная фигура восприниматься по-разному в зависимости от эмоций социальной сцены и наоборот? Будет ли отличаться выражение лица и уровень возбуждения участника в зависимости от эмоциональных сигналов от тела и сцены?
В экспериментах 1 и 2 мы исследовали влияние контекста на физиологические реакции на сигналы лица и тела. В эксперименте 1 использовались реалистичные соединения лица и тела, выражающие эмоционально конгруэнтные или неконгруэнтные сигналы гнева, страха и счастья. Мы выбрали эти выражения по следующим причинам. Во-первых, эти три эмоции могут быть одинаково хорошо выражены через тело и лицо, в отличие от удивления и отвращения, которые не очень хорошо распознаются только по мимике тела. Во-вторых, все три эти эмоции являются возбуждающими и содержат явный компонент действия в телесном выражении (в отличие от грустного телесного выражения). В-третьих, гнев, страх и счастливые выражения — это эмоции, которые изучались чаще всего, а также те, которые мы использовали в наших предыдущих исследованиях, в которых мы использовали аналогичные экспериментальные парадигмы (но с другими зависимыми переменными) (Kret and de Gelder, 2010). , 2012a, 2013; Kret et al., 2011a,b,c, 2013). Гневное выражение лица можно интерпретировать как признак доминирования. Напротив, страх может сигнализировать о покорности.
В эксперименте 1 использовались реалистичные соединения лица и тела, выражающие эмоционально конгруэнтные или неконгруэнтные сигналы гнева, страха и счастья. Мы выбрали эти выражения по следующим причинам. Во-первых, эти три эмоции могут быть одинаково хорошо выражены через тело и лицо, в отличие от удивления и отвращения, которые не очень хорошо распознаются только по мимике тела. Во-вторых, все три эти эмоции являются возбуждающими и содержат явный компонент действия в телесном выражении (в отличие от грустного телесного выражения). В-третьих, гнев, страх и счастливые выражения — это эмоции, которые изучались чаще всего, а также те, которые мы использовали в наших предыдущих исследованиях, в которых мы использовали аналогичные экспериментальные парадигмы (но с другими зависимыми переменными) (Kret and de Gelder, 2010). , 2012a, 2013; Kret et al., 2011a,b,c, 2013). Гневное выражение лица можно интерпретировать как признак доминирования. Напротив, страх может сигнализировать о покорности. Улыбка может означать и то, и другое. В контексте агрессивной позы улыбка скорее интерпретируется как доминанта, смех в лицо. Но когда тело выражает страх, улыбка может быть воспринята как аффилиативный сигнал.
Улыбка может означать и то, и другое. В контексте агрессивной позы улыбка скорее интерпретируется как доминанта, смех в лицо. Но когда тело выражает страх, улыбка может быть воспринята как аффилиативный сигнал.
В эксперименте 2 использовались соединения сцен тела, т. е. похожие сердитые и счастливые выражения тела, но в сочетании с натуралистическими социальными сценами, показывающими эмоционально конгруэнтные или неконгруэнтные сердитые, счастливые или нейтральные сцены. В эксперименте 3 распознавание выражений тела участниками тестировалось с теми же стимулами, что и в эксперименте 2, чтобы выяснить, лучше ли распознаются позы тела в эмоционально конгруэнтной или неконгруэнтной контекстной сцене (Kret and de Gelder, 2010).
Что касается нашего первого исследовательского вопроса, мы предсказали, что гневные и испуганные выражения лица или тела будут привлекать наибольшее внимание, что согласуется с предыдущими исследованиями, которые показали, что гневные сигналы привлекают внимание больше, чем счастливые сигналы (Öhman и др. , 2001 г., Грин и др., 2003 г., Баннерман и др., 2009 г.). Таким образом, мы ожидали наибольшей продолжительности фиксации на рассерженных телах, особенно когда на одновременно предъявленном лице было счастливое выражение. Кроме того, мы предсказали, что разгневанные лица в сочетании с разгневанными телами вызовут наибольшие значения расширения зрачка, поскольку присутствие обоих сигналов может увеличить общую воспринимаемую интенсивность стимула. Мы ожидаем, что это отразится и на лице участника, то есть наибольшая активность сморщивания в ответ на сердитые лица в сочетании с сердитыми телами, наибольшая скуловая активность, когда счастливые лица сочетались со счастливыми выражениями тела. Во-вторых, мы предположили, что взгляд будет привлекать гнев в теле и сцене, и что внимание будет преимущественно приковано к разгневанному телу, представленному в нейтральном контексте, поскольку нейтральный контекст меньше всего отвлечет внимание от тела. Кроме того, мы ожидали наибольшего расширения зрачка в ответ на стимулы, содержащие наиболее возбуждающие сигналы, т.
, 2001 г., Грин и др., 2003 г., Баннерман и др., 2009 г.). Таким образом, мы ожидали наибольшей продолжительности фиксации на рассерженных телах, особенно когда на одновременно предъявленном лице было счастливое выражение. Кроме того, мы предсказали, что разгневанные лица в сочетании с разгневанными телами вызовут наибольшие значения расширения зрачка, поскольку присутствие обоих сигналов может увеличить общую воспринимаемую интенсивность стимула. Мы ожидаем, что это отразится и на лице участника, то есть наибольшая активность сморщивания в ответ на сердитые лица в сочетании с сердитыми телами, наибольшая скуловая активность, когда счастливые лица сочетались со счастливыми выражениями тела. Во-вторых, мы предположили, что взгляд будет привлекать гнев в теле и сцене, и что внимание будет преимущественно приковано к разгневанному телу, представленному в нейтральном контексте, поскольку нейтральный контекст меньше всего отвлечет внимание от тела. Кроме того, мы ожидали наибольшего расширения зрачка в ответ на стимулы, содержащие наиболее возбуждающие сигналы, т. е. сердитое выражение тела, показанное в агрессивном контексте, и что лицо участника будет отражать валентность всей сцены, включая фигуру на переднем плане. . В целом, мы прогнозируем, что на реакции участников большее влияние оказывают эмоциональные сигналы и что суммируются несколько сигналов одной и той же эмоции, чем несоответствие между несколькими сигналами.
е. сердитое выражение тела, показанное в агрессивном контексте, и что лицо участника будет отражать валентность всей сцены, включая фигуру на переднем плане. . В целом, мы прогнозируем, что на реакции участников большее влияние оказывают эмоциональные сигналы и что суммируются несколько сигналов одной и той же эмоции, чем несоответствие между несколькими сигналами.
Результаты
Эксперимент 1. Составные изображения лица и тела
Участники свободно просматривали сердитое, счастливое и испуганное выражение лица в сочетании с выражением тела во всех комбинациях (сердитое лицо с сердитым, счастливым и испуганным телом, счастливое лицо с сердитым, счастливое и испуганное тело, испуганное лицо с сердитым, счастливым и испуганным телом). См. Рисунок 1A для двух примеров стимула. Этот эксперимент был организован, чтобы выяснить, как спонтанно обрабатываются эмоциональные сигналы от тела (интересующая область тела, ROI) и лица (ROI лица) и в какой степени выражения лица и тела привлекают внимание, вызывают возбуждение и выражение лица у наблюдателя. Все средние значения и стандартные ошибки для всех показателей можно найти в дополнительной таблице 1.
Все средние значения и стандартные ошибки для всех показателей можно найти в дополнительной таблице 1.
Рисунок 1. Эксперимент 1 . Пассивный просмотр пар лица и тела (A) Примеры стимулов (B) Продолжительность фиксации на телах (ROI тела) в основном зависела от выражения тела. Сердитые выражения вызывали самые длительные фиксации. (C) Продолжительность фиксации на телах (ROI тела) под счастливыми лицами была больше, когда тела выражали страх или гнев, чем когда они выражали счастье. (D) Продолжительность фиксации на лицах (область интереса лица) с конгруэнтными выражениями тела показала, что на сердитые лица смотрели дольше, чем на счастливые лица. (E) Корругатор реагировал на сердитые и испуганные лица, независимо от положения тела (F) Размер зрачка был наибольшим при одновременном наблюдении гнева на лице и теле. Столбики погрешностей представляют среднюю стандартную ошибку. × р < 0,1; * р < 0,05; ** р < 0,01; *** р < 0,005.
× р < 0,1; * р < 0,05; ** р < 0,01; *** р < 0,005.
Фиксации на теле
A 3 × 3 (выражение лица × выражение тела) Повторные измерения ANOVA показал, что в интересующей области тела (ROI) мы наблюдали основной эффект выражения тела: испуганные и злые тела рассматривались дольше, чем счастливые тела F (2, 72) = 12,026, стр. = 0,25, SE = 0,03) p < 0,001; страх ( M = 0,27, SE = 0,03) против счастья p = 0,06]. Других основных эффектов или эффектов взаимодействия не наблюдалось (см. рис. 1В).
Чтобы проверить эффекты конгруэнтности, мы провели дисперсионный анализ повторных измерений 2 × 3 с конгруэнтностью (конгруэнтностью или неконгруэнтностью) сигнала тела × выражением лица (гнев, страх, радость) и выражением лица, что дало значительное взаимодействие F (2, 72) = 5,189, p < 0,01, η p 2 = 0,126. Последующий t -тест показал, что на тела дольше смотрели, когда они были эмоционально неконгруэнтны по сравнению с конгруэнтными счастливому выражению лица (т. = 3,799, р = 0,001. При включении только конгруэнтных стимулов мы не обнаружили статистически значимого влияния эмоций, хотя наблюдалась тенденция с несколько большей фиксацией, приписываемой ROI тела в случае гнева по сравнению со страхом или счастливыми позами F (2, 72) = 2,330, p = 0,10, η p 2 = 0,061 (см. рис. 1C) 1 .
= 3,799, р = 0,001. При включении только конгруэнтных стимулов мы не обнаружили статистически значимого влияния эмоций, хотя наблюдалась тенденция с несколько большей фиксацией, приписываемой ROI тела в случае гнева по сравнению со страхом или счастливыми позами F (2, 72) = 2,330, p = 0,10, η p 2 = 0,061 (см. рис. 1C) 1 .
Фиксации на лице
A 3 × 3 (выражение лица × выражение тела) Повторные измерения Дисперсионный анализ неожиданно показал, что фиксации на лице не модулируются выражениями лица, а демонстрируют только статистические тенденции F (2, 72) = 2,779, p = 0,069, η p 2 = 0,072. Взаимодействие между выражением лица и тела также показало статистическую тенденцию к значимости 9.0141 F (4, 144) = 2,212, p = 0,071, η p 2 = 0,058. Дальнейшие испытания не выявили существенных различий. Не было никакого основного эффекта выражения тела.
Чтобы проверить эффекты конгруэнтности, мы провели дисперсионный анализ повторных измерений 2 × 3 с конгруэнтностью сигнала тела и выражения лица, что дало значительное взаимодействие F (2, 72) = 4,272, p < 0,05, η р 2 = 0,106. Продолжение t -тест показал, что сердитые лица несколько дольше рассматривались в паре с сердитыми, чем со счастливыми или испуганными телами t (36) = 1,951, p = 0,059. При включении в дисперсионный анализ повторных измерений только конгруэнтных стимулов мы наблюдали влияние выражения лица F (2, 72) = 5,664, p = 0,005, η p 2 = 0,133. В конгруэнтном состоянии на сердитые лица смотрели дольше, чем на счастливые лица (9).0141 p <0,05) (см. рис. 1D). EMG-гофроагрегат . A 3 × 3 (лицо × выражение тела) Повторные измерения ANOVA показали основной эффект выражения лица F (2, 56) = 11,394, p < 0,001, η p 2 = 0,289, corrugator активность показала избирательное увеличение после злых и испуганных лиц ( M = 100,61, SE = 1,06 и M = 100,14, SE = 1,06) по сравнению со счастливыми лицами ( M = 97,54, SE = 1,05) ( p — значения < 0,005). Таблица 1. Электромиография . ЭМГ скуловой . Анализ повторных измерений 3 × 3 (выражение лица × тела) показал, что скуловая мышца в равной степени реагировала на все стимулы, т. е. не было значительных эффектов выражения лица или тела. Других основных или интерактивных эффектов не было. Мы не обнаружили признаков конгруэнтности (см. Таблицу 1). Чтобы проверить, было ли отсутствие основного влияния тел на ответы ЭМГ связано с короткими фиксациями на телах, мы рассчитали корреляции между продолжительностью фиксации и активностью скуловой мышцы и мышц, сморщивающих мышцы. Пупиллометрия . Мы проанализировали размер зрачка с помощью дисперсионного анализа с повторными измерениями 3 × 3. Результаты не показали основных эффектов или эффектов взаимодействия. Чтобы проверить эффекты конгруэнтности, мы провели дисперсионный анализ повторных измерений 2 × 3 с конгруэнтностью сигнала тела и выражения лица, что дало значительное взаимодействие F (2, 72) = 3,653, p < 0,05, η р 2 = 0,092. Злые лица вызывали большее расширение зрачков в сочетании с сердитыми, чем с напуганными или счастливыми телами т (36) = 2,610, р < 0,05. В повторных измерениях ANOVA только с эмоционально конгруэнтными стимулами наблюдалось сильное влияние эмоций F (2, 72) = 5,701, p < 0,005. Чтобы проверить, не связано ли отсутствие основного влияния тел на реакцию зрачков с короткой фиксацией на телах, мы рассчитали корреляцию между временем взгляда и размером зрачка. Мы не нашли доказательств такой связи. Мы также исследовали корреляцию между фиксацией на голове и размером зрачка и обнаружили одну значимую отрицательную корреляцию между продолжительностью фиксации на ROI головы счастливых лиц над испуганными телами и размером зрачка (9).0141 r = -0,452, p = 0,005, без поправки, p = 0,045, с поправкой Бонферрони). Этот вывод согласуется с нашим выводом о том, что размеры зрачков были наименьшими после сигналов счастья по сравнению с сигналами гнева или страха, поэтому чем дольше участники зацикливались на сигналах счастья, тем меньше должны быть размеры их зрачков. В эксперименте 1 участники наблюдали составные изображения лица и тела, и мы показали, что взгляды участников были привлечены угрожающими сигналами тела, что зрачки участников расширялись в основном в ответ на конгруэнтные гневные сигналы и что корругатор реагировал на злые и испуганные лица, но не на тела. В эксперименте 2 те же участники рассматривали новый набор натуралистических стимулов, состоящий из сердитых и счастливых выражений тела, расположенных в сердитых, счастливых и нейтральных социальных сценах. Мы часто сталкиваемся с кем-то в контексте, который включает в себя других людей. Особенно, когда вы видите кого-то эмоциональным, контекст и, в частности, социальный контекст могут способствовать пониманию эмоций наблюдаемого. Цель эксперимента 2 состояла в том, чтобы исследовать, как обрабатываются выражения тела, когда они представлены в социально-эмоциональном контексте. A 2 × 3 (выражение тела × эмоциональная сцена) Повторные измерения ANOVA выявил взаимодействие между эмоциями тела и сцены на продолжительности фиксации в теле ROI F (2, 72) = 3,991, р < 0,05, η р 2 = 0,100. В нейтральной сцене на злых тел смотрели дольше, чем на счастливых [ t (36) = 3,120, p < 0,05]. Напротив, в эмоциональных сценах эти различия исчезали. Подсчитывали длительность фиксаций, приходящихся на область кисти ( М = 0,06, СЭ = 0,004). Поскольку большинство участников хотя бы раз зацикливались на руках, мы решили дополнительно проанализировать этот паттерн. Не было никакого основного воздействия телесного выражения и взаимодействия между телесным выражением и эмоциями сцены на фиксации в области руки. Существенного влияния эмоциональности сцены не было, хотя тенденция наблюдалась F (2, 72) = 2,360, p = 0,10, η p 2 = 0,062 по длительности фиксации на руках, но последующие сравнения не показали достоверных различий ( p ≥ 0,08). Никаких эффектов не было обнаружено, когда мы тестировали конгруэнтность только с эмоциональными состояниями (исключая нейтральные сцены) (см. Таблица 2. Длительность фиксации . Рис. 2. Эксперимент 2 . Пассивный просмотр тел в социальных сценах. (А) Две тепловые карты показывают, что участники зациклились на людях в этой сцене. (B) Зрачки участников расширились в основном в ответ на гневные сигналы, исходящие как от тела, так и от сцены (C) . (D) Напротив, гофроагрегат реагировал исключительно на сцены гнева, а не на тела гнева. (E) Точно так же zygomaticus реагировал на счастливые сцены и не реагировал на мимику тела. * р < 0,05; *** р < 0,005. Расширение зрачка . A 2 × 3 (эмоции тела × сцены) Повторные измерения ANOVA выявили основные эффекты эмоций тела и сцены ; F (2, 72) = 8,049, р < 0,005, η р 2 = 0,183. Размер зрачка был больше после разгневанных и счастливых тел ( M = 174,08, SE = 21,11 против M = 133. ЭМГ-гофроагрегат . ANOVA повторных измерений 2 × 3 (эмоции тела × сцены) показал, что корругатор более чувствителен к сценам гнева по сравнению со сценами счастья ( M = 106,634, SE = 2,060 против 100,966, SE = 0,699) ( p < 0,05) F (2, 72) = 5,584, p < 0,01, η p 2 = 0,134. Не было никакого основного эффекта выражения тела и никакого взаимодействия. Никаких эффектов не было обнаружено, когда мы проверяли соответствие только эмоциональным состояниям (см. ЭМГ-скуловой . ANOVA с повторными измерениями 2 × 3 (тело × эмоция сцены) показал, что скуловые кости показали противоположный характер реакции [больше для счастья ( M = 110,004, SE = 2,653) по сравнению со сценами гнева ( M = 101,257, SE = 2,093) ( p < 0,005) и незначительно значимо для счастья ( М = 107,413, СЭ = 2,653) р = 0,069]. Не было никаких основных эффектов или взаимодействий с выражением тела. Когда мы проверили конгруэнтность, включив только эмоциональные состояния, мы обнаружили взаимодействие между конгруэнтностью тела и сцены F (1, 36) = 11,968, p = 0,001, η p 2 = 0,250. Реакция скуловой кости была сильнее после счастливых тел в сценах счастья, чем в сценах гнева 90–141 t 90–142 90–195 (36) 90–196 = 2,378, p <0,05 (см. рис. 2Е). Мы исследовали возможную взаимосвязь между длительностью фиксации и ЭМГ и реакцией зрачка, но не нашли доказательств какой-либо взаимосвязи. После завершения экспериментов 1 и 2 мы еще раз показали тем же участникам стимулы эксперимента 2 и попросили их классифицировать выражение тела, игнорируя сцену. эмоции, что было легко, поскольку стимулы предъявлялись только в течение 100 мс (Kret and de Gelder, 2010). Мы предсказывали, что обнаружим эффект конгруэнтности в том, что участники смогут лучше распознавать выражения тела, когда они представлены в эмоционально конгруэнтной контекстной сцене по сравнению с эмоционально неконгруэнтной контекстной сценой. Точность . ANOVA повторных измерений 2 × 3 (тело × эмоция сцены) показал, что существует взаимодействие между эмоциями тела и сцены = 0,127. Разгневанные тела лучше распознавались в контексте гнева и счастья t (35) = 2,477, p = 0,018, а счастливые тела несколько лучше распознавались в контексте счастья и гнева, хотя этот эффект не достиг статистической значимости т (35) = 1,755, р = 0,088. Тридцать семь участников (26 женщин, средний возраст 22,7 года, диапазон 19–29 лет; 11 мужчин, средний возраст: 23,8 года, диапазон 19–32 лет) заполнили информированную анкету. согласие и приняли участие во всех трех экспериментах и в дополнительных задачах распознавания эмоций, которые опубликованы в другом месте (Kret et al., 2013). Участники не имели неврологического или психиатрического анамнеза, были правшами и имели нормальное или скорректированное до нормального зрение. Исследование проведено в соответствии с Хельсинкской декларацией и одобрено местным комитетом по медицинской этике. Испуганные, счастливые и сердитые выражения лиц шести лиц мужского пола, которые были правильно распознаны выше 80%, были выбраны из набора NimStim (Tottenham et al., 2009). Соответствующие выражения тела были взяты из нашей собственной базы данных стимулов, содержащей 254 цифровых изображения. После наложения электродов на лица участников устройство слежения за глазами было помещено на голову участника. Далее выполнялась 9-точечная калибровка, которая повторялась перед каждым блоком. Стимулы предъявлялись с помощью программы E-prime на экране ПК с разрешением 1024 на 768 и частотой обновления 100 Гц. Каждое испытание начиналось с фиксированного креста, показываемого в течение как минимум 3000 мс, пока участник не зафиксировался, и руководитель эксперимента не выполнил ручную коррекцию дрейфа, после чего показывали изображение в течение 4000 мс и серый экран (3000 мс). Стимулы были разделены на два блока по 36 проб, в каждом по 18 совпадающих и 18 неконгруэнтных стимулов. Чтобы участники оставались наивными в отношении цели электромиографии (ЭМГ), им сказали, что электроды регистрируют пот. Участников попросили посмотреть на картинки, не дав ответа. После эксперимента их попросили описать то, что они видели. Все упомянули, что видели эмоциональные выражения, но иногда выражения лица и тела не были одинаковыми. Фиксации, расширение зрачка и ответы ЭМГ анализировали в отдельных 3 × 3 (выражение лица × выражение тела) повторных измерениях ANOVA. Фиксации анализировались по ROI (тело, руки и лицо ROI), которые определялись пикселями на всем теле (включая шею) и пикселями головы. Значимые основные эффекты были исследованы с помощью попарных сравнений с поправкой Бонферрони, а взаимодействия с помощью двусторонних t -тестов с поправкой Бонферрони. Стимулирующие материалы состояли из конгруэнтных и неконгруэнтных пар тело-сцена (примеры см. на рис. 2А). Фотографии тел (восьми актеров-мужчин, с размытыми чертами лица) были взяты из той же серии, что и в эксперименте 1, и выражали гнев и счастье. Сцены (восемь уникальных сцен для каждого эмоционального состояния) были выбраны из Интернета и показывали злые, счастливые или нейтральные сцены. Количество людей в разных сценах было одинаковым в зависимости от эмоционального состояния. Эти сцены были проверены ранее в задаче на распознавание эмоций и были распознаны очень точно, даже несмотря на то, что они демонстрировались только в течение 100 мс (гнев 88%, счастье 9%). Мы провели дополнительное проверочное исследование среди 36 студентов в соответствии со стандартными проверочными процедурами Брэдли и Ланга (1999). Нейтральные сцены были оценены как значительно более спокойные, чем счастливые сцены 90–141 t 90–142 90–195 (35) 90–196 = 4,098, 90–141 p 90–142 < 0,001 и несколько спокойнее, чем сцены гнева т (35) = 1,836, р = 0,075. Сцены гнева и радости были одинаково эмоционально интенсивными t (35) = 0,462, p = 0,647 и обе были более интенсивными, как нейтральные сцены t (35) = 4,208, < 90; т (35) = 7,109, р < 0,001. Продолжительность предъявления стимула и межпробный интервал в эксперименте 2 были такими же, как и в эксперименте 1. Половина участников начала с Эксперимента 1, а другая половина – с Эксперимента 2. Процедура Эксперимента 2 была такой же, как и для Эксперимента 1, за исключением того, что было 48 испытаний, которые были случайным образом представлены в рамках одного блокировать. Данные были проанализированы в отдельных 2 (эмоции тела) × 3 (эмоции сцены) повторных измерений ANOVA. После завершения Экспериментов 1 и 2 мы еще раз показывали участникам стимулы Эксперимента 2, на этот раз с короткой продолжительностью предъявления (100 мс) и с заданием классифицировать выражение тела, игнорируя эмоция сцены. Долю правильных ответов анализировали с помощью дисперсионного анализа повторных измерений 2 (эмоции тела) × 3 (эмоции сцены). Параметры для регистрации и анализа ЭМГ лица были выбраны в соответствии со стандартными рекомендациями (Van Boxtel, 2010). Использовались активные электроды BioSemi плоского типа, и ЭМГ лица измерялась биполярно над большой скуловой мышцей и мышцей, сморщивающей бровь, на правой стороне лица с частотой дискретизации 1024 Гц. Скуловая мышца в основном участвует в выражении счастья. Корругаторную мышцу можно использовать для измерения выражения негативных эмоций, включая гнев и страх. Но для того, чтобы различить эти две отрицательные эмоции, необходимо измерить дополнительные лицевые мышцы, такие как лобная мышца (Ekman and Friesen, 1978). Однако в текущем эксперименте это было невозможно из-за айтрекера, закрепленного на голове. Активность корругатора в определенном контексте, например, при предъявлении четких эмоциональных стимулов, можно интерпретировать как выражение наблюдаемой эмоции (Overbeek et al., 2012). Движения глаз регистрировали с частотой дискретизации 250 Гц с помощью наголовной системы EyeLink Eye Tracking System (SensoMotoric Instruments GmbH, Германия). Коррекция дрейфа выполнялась в каждом испытании, чтобы гарантировать, что данные были скорректированы с учетом движения. Мы исследовали восприятие эмоциональных выражений, используя натуралистические стимулы, состоящие из выражений всего тела и сцен. Из исследований вытекают два основных вывода: (1) реакция наблюдателей на выражение лица и тела зависит от выражения всего тела и окружающей социальной среды. Таким образом, на восприятие выражений лица и тела влияют естественные условия просмотра лица и тела. (2) Когда люди сталкиваются с угрозой, исходящей от лица, тела или сцены, зрачки участников расширяются, их мышцы, сморщивающие губы, становятся более активными, и они направляют свой взгляд на угрозу. Эти выводы основаны на результатах двух основных экспериментов. В эксперименте 1 были показаны эмоционально конгруэнтные и неконгруэнтные пары лицо-тело. Эксперимент 2 показал эмоционально конгруэнтные и неконгруэнтные пары тело-сцена. В эксперименте 1, где участники наблюдали конгруэнтные и неконгруэнтные пары «лицо-тело», мы показали, что участники не только смотрели на выражение лица, но и выбирали сигналы всего тела. Участники всегда сканировали лицо и тело. Это может отражать стратегию, используемую наблюдателями, желающими проверить эмоции, наблюдаемые на лице. В ходе развития люди узнают, что в социальных ситуациях, и в частности в стрессовых ситуациях, люди пытаются контролировать выражение своего лица и улыбаются, когда не чувствуют себя счастливыми или не в своей тарелке (de Gelder et al. Также наш второй основной вывод согласуется с предыдущими исследованиями. Участники обращали внимание в основном на угрожающие реплики. Например, аналогичные результаты были получены Green et al. (2003), которые обнаружили более длительную фиксацию на выражениях, связанных с угрозой, включая гнев, по сравнению с выражениями, не относящимися к угрозе (такими как счастье). Кроме того, исследования визуального поиска показали, что сердитые лица обычно распознаются быстрее и точнее, чем счастливые (Fox et al., 1987; Öhman et al., 2001; Lundqvist and Ohman, 2005). Таким образом, распределение внимания во время социальных взаимодействий может отражать необходимость подготовки адаптивного ответа на социальную угрозу. Только счастливое выражение лица сигнализировало бы о безопасности и, следовательно, было бы наименее релевантным, на что указывает более короткая фиксация. Предыдущие исследования ЭМГ постоянно демонстрировали, что люди склонны реагировать конгруэнтной активностью лицевых мышц, когда смотрят на эмоциональные лица (Hess and Fischer, 2013). Действительно, в эксперименте 1 корругатор участников был более активен при наблюдении за сердитыми и испуганными лицами, а не за счастливыми, но независимо от выражения тела, с которым они сочетались. Более того, zygomaticus в этом эксперименте не различал выражения лица. Участники всегда в той или иной степени улыбались в ответ на все раздражители. Но здесь выражения лица и тела были совмещены, и, возможно, для ответа ЭМГ (и для пупиллометрии) наличие выражения лица превалировало над реакцией на выражение тела. Был поставлен вопрос, реагируют ли скуловые и корругаторные мышцы исключительно на выражение лица или реагируют более широко. Предыдущие исследования предполагают последнее. Например, два более ранних исследования показали выражение эмоций на лице, которое было похоже на эмоции, выраженные либо телом, либо голосом (Magnee et al. Эксперимент 2 демонстрирует, что выражение лица участника отражает эмоции социальных сцен, в которых все выражения лиц были размыты, но были видны выражения тела людей на заднем плане. Таким образом, корругатор и скуловой аппарат реагируют не только на лица, но и на другие сигналы. В эксперименте 1 корругатор реагировал на выражение лица, но не на выражение тела. Кажется, что для ответа ЭМГ наличие выражения лица, даже если оно меньше по размеру, чем положение всего тела, перевешивает эффект выражения тела. То же самое можно сказать и о сценах: присутствие толпы, испытывающей определенную эмоцию, отменяет эмоциональную синхронизацию с одной эмоциональной позой тела впереди. Эмоциональное возбуждение является ключевым элементом модуляции реакции зрачка (Gilzenrat et al., 2010). В экспериментах 1 и 2 мы показали, что размер зрачка участников был самым большим в ответ на разгневанные лица, тела и сцены. Хотя интенсивность эмоций, отображаемых в сценах счастья и гнева, оценивалась одинаково, сцены гнева вызывали большее возбуждение. Счастливые сцены были четко распознаны как счастливые сцены, а гневные сцены — как гневные. Эти данные опровергают более ранние гипотезы о том, что диаметр зрачка увеличивается, когда люди обрабатывают эмоционально привлекательные стимулы, независимо от гедонистической валентности (Bradley et al., 2008). Расширение зрачка при постоянном освещении вызывается норадреналином, вырабатываемым в голубом пятне. Различные физиологические манипуляции (например, тревога, вредная/болевая стимуляция) повышают активность в этой области и приводят к повышенному возбуждению и изменениям вегетативной функции, согласующимся с симпатической активацией (Gilzenrat et al. Здравый смысл подсказывает, что мы читаем выражение лица так же, как читаем отдельные слова на странице, напрямую и недвусмысленно обращаясь к значению слова вне контекста предложения. Но это не так, поскольку выражение лица воспринимается по-разному, в зависимости от выражения тела. Выражения тела также не свободны от контекстуальных влияний, и участники сканируют тело по-разному, в зависимости от выражения лица и социальной сцены. В целом, мы обнаружили, что участники больше всего обращали внимание на сигналы гнева и страха, и размеры их зрачков значительно увеличились. Выражения лиц участников соответствовали валентности стимулов. Однако, когда выражение лица сочеталось с выражением тела, наблюдаемые лица сильнее влияли на ЭМГ-ответы, чем выражения тела. Mariska E. Kret и Jeroen J. Stekelenburg участвовали в сборе и фильтрации данных, Mariska E. Kret проанализировала данные и подготовила рисунки. Маришка Э. Крет, Йерун Дж. Стекеленбург, Карин Рулофс и Беатрис де Гелдер внесли свой вклад в написание основного текста рукописи. Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов. Мы благодарим A. van Boxtel за его советы относительно измерений ЭМГ, J. Shen за помощь с системой Eyelink, Jorik Caljauw из службы поддержки Matlab за помощь в написании скриптов. Исследование было поддержано грантами NWO (Nederlandse Organisatie voor Wetenschappelijk Onderzoek; 400.04081) и Европейской комиссии (COBOL FP6-NEST-043403 и FP7 TANGO) для Беатрис де Гелдер, грантом Vidi (№ 452-07008) от NWO для Карин Рулофс и по гранту Королевской академии наук Нидерландов (KNAW Dr. JL Dobberke Stichting) Мариске Э. Крет. За разработкой набора стимуляторов для лица MacBrain наблюдал Ним Тоттенхэм и поддерживала Исследовательская сеть Фонда Д. Джона и Кэтрин Т. Макартур по раннему опыту и развитию мозга. Пожалуйста, свяжитесь с Ним Тоттенхэм по адресу [email protected] для получения дополнительной информации о наборе стимулов. 9Мы также извлекли фиксации из ROI на глазах, рту и руках, но из-за небольшого размера наблюдали эффект пола ( M = 0,03, SE = 0,004), поэтому их дальнейший анализ не проводился. Адольф Р. (2002). Распознавание эмоций по выражению лица: психологические и неврологические механизмы. Поведение. Познан. Неврологи. Версия . 1, 21–61. doi: 10.1177/1534582302001001003 Опубликовано Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка Авиезер Х., Троп Ю. и Тодоров А. (2012). Сигналы тела, а не выражение лица, позволяют различать сильные положительные и отрицательные эмоции. Наука 338, 1225–1229. doi: 10.1126/science.1224313 Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Баннерман Р.Л., Милдерс М., де Гелдер Б. и Сахрайе А. (2009). Ориентация на угрозу: более быстрая локализация испуганных выражений лица и поз тела, выявляемых саккадическими движениями глаз. Проц. биол. Наука . 276, 1635–1641. doi: 10.1098/rspb.2008.1744 Pubmed Abstract | Опубликован полный текст | CrossRef Full Text Bandettini, P.A., Wong, E.C., Hinks, R. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Брэдли, М.М., и Ланг, П.Дж. (1999). Эмоциональные нормы для английских слов (ANEW): Инструкция по эксплуатации и эмоциональные рейтинги. Технический отчет C-1 . Гейнсвилл, Флорида: Центр исследований психофизиологии Университета Флориды. Брэдли М.М., Микколи Л., Эскриг М.А. и Ланг П.Дж. (2008). Зрачок как мера эмоционального возбуждения и вегетативной активации. [Научная поддержка, Национальный институт здравоохранения, заочная]. Психофизиология 45, 602–607. doi: 10.1111/j.1469-8986.2008.00654.x Опубликовано Аннотация | Опубликован полный текст | Полнотекстовая перекрестная ссылка Брэдли, М. М., Сабатинелли, Д., Ланг, П. Дж., Фитцсиммонс, Дж. Р., Кинг, В., и Десаи, П. (2003). Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef де Гелдер, Б., и Ван ден Сток, Дж. (2011). Тест телесно-выразительных действий (BEAST). Построение и проверка основы стимула для измерения восприятия выражения эмоций всего тела. Фронт. Психол . 2:181. doi: 10.3389/fpsyg.2011.00181 Опубликовано Аннотация | Опубликован полный текст | CrossRef Full Text de Gelder, B., van den Stock, J., Meeren, HKM, Sinke, CBA, Kret, M.E., and Tamietto, M. (2010). Встаем за тело. Недавний прогресс в раскрытии сетей, участвующих в обработке тел и телесных выражений. Неврологи. Биоповедение. Версия . 34, 513–527. doi: 10.1016/j.neubiorev.2009.10.008 Pubmed Abstract | Опубликован полный текст | Полнотекстовая перекрестная ссылка Димберг, У. (1982). Реакции лица на мимику. Психофизиология 19, 643–647. Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef Экман П. и Фризен В. В. (1978). Руководство по системе кодирования движений лица . Пало-Альто, Калифорния: Consulting Psychologists Press. Фокс, Э., и Дамьянович, Л. (2006). Глаз достаточно, чтобы произвести эффект превосходства угрозы. Эмоция 6, 534–539. doi: 10.1037/1528-3542.6.3.534 Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Фокс, П. Т., Бертон, Х., и Райхл, М. Е. (1987). Картирование соматосенсорной коры человека с помощью позитронно-эмиссионной томографии. Дж. Нейрохирург . 67, 34–43. doi: 10.3171/jns.1987.67.1.0034 Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Gilzenrat, M.S., Nieuwenhuis, S., Jepma, M., and Cohen, J.D. (2010). Диаметр зрачка отслеживает изменения в состоянии контроля, предсказанные теорией адаптивного усиления функции голубого пятна. Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef Грин, М.Дж., Уильямс, Л.М., и Дэвидсон, Д.Дж. (2003). Перед лицом опасности: конкретные стратегии просмотра выражения лица угрозы? Познан. Эмот . 17, 779–786. doi: 10.1080/02699 Взаимодействие между выражением лица и телом было незначительным, но показало тенденцию F (4, 112) = 2,087, p = 0,087, η p 2 = 0,069. Дальнейшие испытания не выявили существенных различий. Не было никакого основного эффекта выражения тела. Мы не обнаружили никаких указаний на эффекты конгруэнтности, поскольку были проверены с помощью дисперсионного анализа повторных измерений 2 × 3 (конгруэнтность × выражение лица) с конгруэнтностью сигнала тела и выражением лица в качестве факторов (см. Таблицу 1). См. рисунок 1Е.
Взаимодействие между выражением лица и телом было незначительным, но показало тенденцию F (4, 112) = 2,087, p = 0,087, η p 2 = 0,069. Дальнейшие испытания не выявили существенных различий. Не было никакого основного эффекта выражения тела. Мы не обнаружили никаких указаний на эффекты конгруэнтности, поскольку были проверены с помощью дисперсионного анализа повторных измерений 2 × 3 (конгруэнтность × выражение лица) с конгруэнтностью сигнала тела и выражением лица в качестве факторов (см. Таблицу 1). См. рисунок 1Е. Мы не нашли доказательств такой связи. Другие исследования показали четкие ответы ЭМГ на невидимые стимулы, предполагая, что модели фиксации не должны влиять на ответы ЭМГ (Tamietto et al., 2009).).
Мы не нашли доказательств такой связи. Другие исследования показали четкие ответы ЭМГ на невидимые стимулы, предполагая, что модели фиксации не должны влиять на ответы ЭМГ (Tamietto et al., 2009).). Наблюдение за рассерженными людьми ( M = 157,86, SE = 20,99) вызывало большее расширение зрачков, чем наблюдение за напуганными ( M = 118,48, SE = 23,77) ( p (20 192 < 0,09) 14 человек) или счастливыми = 96,44, SE = 23,77) ( p < 0,005). Не было никакой разницы между страхом и счастьем (см. рис. 1F).
Наблюдение за рассерженными людьми ( M = 157,86, SE = 20,99) вызывало большее расширение зрачков, чем наблюдение за напуганными ( M = 118,48, SE = 23,77) ( p (20 192 < 0,09) 14 человек) или счастливыми = 96,44, SE = 23,77) ( p < 0,005). Не было никакой разницы между страхом и счастьем (см. рис. 1F). Эти исследовательские анализы можно найти в дополнительной таблице 2.
Эти исследовательские анализы можно найти в дополнительной таблице 2. Эксперимент 2. Составные изображения тела и сцены
 Фигура со счастливым или сердитым выражением лица, обращенная к участнику, помещалась в центр толпы, состоящей из других эмоциональных или нейтральных фигур. Центральную фигуру было легко отличить от толпы, так как она всегда появлялась в центре сцены, лицом к наблюдателю. Ключевые вопросы заключались в том, будет ли по-разному смотреться центральная фигура в зависимости от эмоций социальной сцены и будут ли различаться выражение лица и уровень возбуждения участника в зависимости от эмоциональных сигналов от представленного тела и сцены.
Фигура со счастливым или сердитым выражением лица, обращенная к участнику, помещалась в центр толпы, состоящей из других эмоциональных или нейтральных фигур. Центральную фигуру было легко отличить от толпы, так как она всегда появлялась в центре сцены, лицом к наблюдателю. Ключевые вопросы заключались в том, будет ли по-разному смотреться центральная фигура в зависимости от эмоций социальной сцены и будут ли различаться выражение лица и уровень возбуждения участника в зависимости от эмоциональных сигналов от представленного тела и сцены. Фиксации на теле
 Основных эффектов не было. Никаких эффектов не было обнаружено, когда мы тестировали конгруэнтность только с эмоциональными состояниями (т. е. с помощью дисперсионного анализа с повторными измерениями 2 × 2 без нейтрального условия).
Основных эффектов не было. Никаких эффектов не было обнаружено, когда мы тестировали конгруэнтность только с эмоциональными состояниями (т. е. с помощью дисперсионного анализа с повторными измерениями 2 × 2 без нейтрального условия). Таблицу 2). См. рисунок 2А.
Таблицу 2). См. рисунок 2А. 56, SE = 21.63) ( p < 0.005) and angry vs. neutral ( M = 185.73, SE = 28.02 vs. 109.66, SE = 20.38) ( p < 0.05 ) и счастливых против нейтральных ( M = 166,07, SE = 20,46 против M = 109) ( p < 0,001) сцен. Взаимодействий не было. Никаких эффектов не было обнаружено, когда мы проверяли соответствие только эмоциональным состояниям (см. рисунки 2B, C).
56, SE = 21.63) ( p < 0.005) and angry vs. neutral ( M = 185.73, SE = 28.02 vs. 109.66, SE = 20.38) ( p < 0.05 ) и счастливых против нейтральных ( M = 166,07, SE = 20,46 против M = 109) ( p < 0,001) сцен. Взаимодействий не было. Никаких эффектов не было обнаружено, когда мы проверяли соответствие только эмоциональным состояниям (см. рисунки 2B, C). Рисунок 2D).
Рисунок 2D).
Эксперимент 3. Быстрое распознавание выражений тела на составных изображениях тела и сцены

Методы
Участники Эксперименты 1–3
Материалы Эксперимент 1
 Снимки сделаны в профессиональной фотостудии при постоянном освещении. Непрофессиональные актеры были индивидуально проинструктированы по стандартной процедуре отображать четыре выражения (гнев, страх, счастье и печаль) всем телом. В инструкциях было указано несколько конкретных и репрезентативных ежедневных событий, обычно связанных с каждой эмоцией (подробнее см. de Gelder and Van den Stock, 2011). Для текущего исследования мы выбрали лучших актеров с оценкой правильности распознавания выше 80%. Мы использовали только мужские лица и тела, потому что ранее мы обнаружили, что они вызывают более сильное возбуждение, когда выражаются гнев и страх (Kret et al., 2011a; Kret и de Gelder, 2012b). Изображения были представлены в оттенках серого на сером фоне. С помощью Adobe Photoshop яркость каждого стимула была отрегулирована до среднего значения. Окончательная проверка была сделана с помощью люксметра на экране тестового компьютера. Размер стимулов составлял 354 × 532 пикселя. См. рисунок 1A для двух примеров.
Снимки сделаны в профессиональной фотостудии при постоянном освещении. Непрофессиональные актеры были индивидуально проинструктированы по стандартной процедуре отображать четыре выражения (гнев, страх, счастье и печаль) всем телом. В инструкциях было указано несколько конкретных и репрезентативных ежедневных событий, обычно связанных с каждой эмоцией (подробнее см. de Gelder and Van den Stock, 2011). Для текущего исследования мы выбрали лучших актеров с оценкой правильности распознавания выше 80%. Мы использовали только мужские лица и тела, потому что ранее мы обнаружили, что они вызывают более сильное возбуждение, когда выражаются гнев и страх (Kret et al., 2011a; Kret и de Gelder, 2012b). Изображения были представлены в оттенках серого на сером фоне. С помощью Adobe Photoshop яркость каждого стимула была отрегулирована до среднего значения. Окончательная проверка была сделана с помощью люксметра на экране тестового компьютера. Размер стимулов составлял 354 × 532 пикселя. См. рисунок 1A для двух примеров.
Процедура Эксперимент 1

Материалы Эксперимент 2
 7% и нейтральные 92%) (Kret and de Gelder, 2010). Здесь мы исключили страшные тела и сцены и вместо этого включили нейтральные сцены. Включение гнева, страха, счастливых и нейтральных тел и сцен дало бы слишком много условий.
7% и нейтральные 92%) (Kret and de Gelder, 2010). Здесь мы исключили страшные тела и сцены и вместо этого включили нейтральные сцены. Включение гнева, страха, счастливых и нейтральных тел и сцен дало бы слишком много условий.
Процедура Эксперимента 2
Процедура Эксперимент 3
Измерения
ЭМГ лица
 Активный электрод общего режима (CMS) и пассивный электрод ведомой правой ноги (DRL) были прикреплены к левой щеке и использовались в качестве эталонного и заземляющего электродов соответственно (http://www.biosemi.com/faq.htm). Перед наложением кожу протирали спиртом, а электроды заполняли электродной пастой. Необработанные данные были сначала отфильтрованы в автономном режиме с полосой пропускания 20–500 Гц в Brain Vision Analyzer версии 1.05 (Brain Products GmbH) и выпрямлены по полной волне. Данные были визуально проверены на наличие чрезмерных движений во время исходного уровня двумя независимыми оценщиками, которые не знали условий испытания. Испытания, которые считались проблематичными, были исключены, что привело к исключению 6,07% ( SD 7.50) испытаний из последующего анализа. Из-за технических проблем данные ЭМГ четырех участников эксперимента 1 и трех участников эксперимента 2 не записывались. Затем рассчитывали среднее исправленное значение ЭМГ в течение 4000 мс постстимульного периода и 1000 мс исходного периода перед стимулом.
Активный электрод общего режима (CMS) и пассивный электрод ведомой правой ноги (DRL) были прикреплены к левой щеке и использовались в качестве эталонного и заземляющего электродов соответственно (http://www.biosemi.com/faq.htm). Перед наложением кожу протирали спиртом, а электроды заполняли электродной пастой. Необработанные данные были сначала отфильтрованы в автономном режиме с полосой пропускания 20–500 Гц в Brain Vision Analyzer версии 1.05 (Brain Products GmbH) и выпрямлены по полной волне. Данные были визуально проверены на наличие чрезмерных движений во время исходного уровня двумя независимыми оценщиками, которые не знали условий испытания. Испытания, которые считались проблематичными, были исключены, что привело к исключению 6,07% ( SD 7.50) испытаний из последующего анализа. Из-за технических проблем данные ЭМГ четырех участников эксперимента 1 и трех участников эксперимента 2 не записывались. Затем рассчитывали среднее исправленное значение ЭМГ в течение 4000 мс постстимульного периода и 1000 мс исходного периода перед стимулом. Среднее исправленное значение ЭМГ выражали в процентах от средней исходной амплитуды ЭМГ до стимула. Процентные показатели амплитуды ЭМГ усреднялись по действительным испытаниям и по эмоциям.
Среднее исправленное значение ЭМГ выражали в процентах от средней исходной амплитуды ЭМГ до стимула. Процентные показатели амплитуды ЭМГ усреднялись по действительным испытаниям и по эмоциям. Eyetracking
 Мы использовали настройки Eyelink по умолчанию, которые определяли моргание как период активности детектора саккад, когда данные о зрачке отсутствовали для трех или более образцов в последовательности. Саккаду определяли как период времени, в течение которого детектор саккад был активен для 2 или более образцов последовательно и продолжался до начала периода бездействия детектора саккад в течение 20 мс. Настраиваемые пороги ускорения (8000°/с) и скорости (30°/с) были установлены для обнаружения саккад не менее 0,5° угла зрения. Фиксация определялась как любой период, который не был морганием или саккадой. Были проведены анализы доли времени, затрачиваемого на просмотр каждой области интереса, по отношению к времени, затраченному на просмотр экрана, при этом первые 200 мс отбрасывались из-за фиксированного положения фиксирующего креста. В соответствии с предыдущей литературой, базовый уровень 500 мс был вычтен из всех последующих точек данных. Отсутствующие данные из-за морганий интерполировались линейно.
Мы использовали настройки Eyelink по умолчанию, которые определяли моргание как период активности детектора саккад, когда данные о зрачке отсутствовали для трех или более образцов в последовательности. Саккаду определяли как период времени, в течение которого детектор саккад был активен для 2 или более образцов последовательно и продолжался до начала периода бездействия детектора саккад в течение 20 мс. Настраиваемые пороги ускорения (8000°/с) и скорости (30°/с) были установлены для обнаружения саккад не менее 0,5° угла зрения. Фиксация определялась как любой период, который не был морганием или саккадой. Были проведены анализы доли времени, затрачиваемого на просмотр каждой области интереса, по отношению к времени, затраченному на просмотр экрана, при этом первые 200 мс отбрасывались из-за фиксированного положения фиксирующего креста. В соответствии с предыдущей литературой, базовый уровень 500 мс был вычтен из всех последующих точек данных. Отсутствующие данные из-за морганий интерполировались линейно. Первые 2 секунды реакции зрачка не были включены в анализ, чтобы избежать влияния начального падения размера зрачка (Bradley et al., 2008; Kret et al., 2013).
Первые 2 секунды реакции зрачка не были включены в анализ, чтобы избежать влияния начального падения размера зрачка (Bradley et al., 2008; Kret et al., 2013). Обсуждение
 Критически и однозначно в обоих экспериментах мы объединили ЭМГ, реакции зрачков, а также фиксации на лицах, телах и сценах, а также проверили субъективные эмоциональные оценки (Эксперимент 3). Наши основные выводы подтверждают теорию мотивированного внимания (Lang and Cuthbert, 19).97; Брэдли и др., 2003). В соответствии с этой теорией зрительное внимание, на которое здесь указывают фиксации, находилось под влиянием эмоциональности стимула и было направлено на мотивационно значимые сигналы по сравнению с менее важными и не было специально направлено на эмоционально неконгруэнтные сигналы. Угрожающие сигналы, особенно злые сигналы от лиц, тел или сцен, рассматривались дольше, чем счастливые сигналы. Точно так же зрачки участников расширялись в ответ на различные категории социальных аффективных стимулов (лица, тела, сцены) и были значительно больше после гневных сигналов, чем счастливых или нейтральных сигналов. Таким образом, угрожающие сигналы привлекали внимание и вызывали возбуждение.
Критически и однозначно в обоих экспериментах мы объединили ЭМГ, реакции зрачков, а также фиксации на лицах, телах и сценах, а также проверили субъективные эмоциональные оценки (Эксперимент 3). Наши основные выводы подтверждают теорию мотивированного внимания (Lang and Cuthbert, 19).97; Брэдли и др., 2003). В соответствии с этой теорией зрительное внимание, на которое здесь указывают фиксации, находилось под влиянием эмоциональности стимула и было направлено на мотивационно значимые сигналы по сравнению с менее важными и не было специально направлено на эмоционально неконгруэнтные сигналы. Угрожающие сигналы, особенно злые сигналы от лиц, тел или сцен, рассматривались дольше, чем счастливые сигналы. Точно так же зрачки участников расширялись в ответ на различные категории социальных аффективных стимулов (лица, тела, сцены) и были значительно больше после гневных сигналов, чем счастливых или нейтральных сигналов. Таким образом, угрожающие сигналы привлекали внимание и вызывали возбуждение. Напротив, корругаторная мышца участников отражала валентность, как показано в выражении лица наблюдаемого, но не в парном выражении тела. Однако, когда участники смотрели сцену с позой тела на переднем плане, и мышца, сморщивающая морщины, и скуловая мышца реагировали исключительно на сцены, которые включали выражения тела нескольких людей, где выражения лиц были размыты. Теперь мы обсудим эти результаты более подробно, начиная с обсуждения фиксаций участников, а затем ответов ЭМГ и размера зрачка.
Напротив, корругаторная мышца участников отражала валентность, как показано в выражении лица наблюдаемого, но не в парном выражении тела. Однако, когда участники смотрели сцену с позой тела на переднем плане, и мышца, сморщивающая морщины, и скуловая мышца реагировали исключительно на сцены, которые включали выражения тела нескольких людей, где выражения лиц были размыты. Теперь мы обсудим эти результаты более подробно, начиная с обсуждения фиксаций участников, а затем ответов ЭМГ и размера зрачка. Длительность фиксации
 , 2010). Следовательно, их язык тела может быть более информативным. Это неявное знание могло привлечь внимание участников к телу. Эта гипотеза согласуется с нашим выводом о том, что на тела дольше смотрели, когда они были эмоционально неконгруэнтны, а не конгруэнтны счастливому выражению лица (т. е. когда тела были наиболее заметными). Однако результаты нашего предыдущего исследования ЭЭГ, демонстрирующие эффекты быстрой интеграции лица и тела (Meeren et al., 2005) и лица и контекста (Righart and de Gelder, 2008a), говорят против этого объяснения. Ранее предполагалось, что наблюдатели автоматически обращают внимание на тело, чтобы уловить действие наблюдаемого и подготовить свой собственный ответ (Kret et al., 2011a,b). Злой жест тела имеет самые прямые последствия для наблюдателя «бей / беги», и, возможно, поэтому он привлек наибольшее внимание. Следовательно, в Эксперименте 2 эти требования к действию наиболее заметны в гневных жестах тела, показанных в нейтральном контексте, где угрожающий передний план явно выделяется из незаметной фоновой сцены.
, 2010). Следовательно, их язык тела может быть более информативным. Это неявное знание могло привлечь внимание участников к телу. Эта гипотеза согласуется с нашим выводом о том, что на тела дольше смотрели, когда они были эмоционально неконгруэнтны, а не конгруэнтны счастливому выражению лица (т. е. когда тела были наиболее заметными). Однако результаты нашего предыдущего исследования ЭЭГ, демонстрирующие эффекты быстрой интеграции лица и тела (Meeren et al., 2005) и лица и контекста (Righart and de Gelder, 2008a), говорят против этого объяснения. Ранее предполагалось, что наблюдатели автоматически обращают внимание на тело, чтобы уловить действие наблюдаемого и подготовить свой собственный ответ (Kret et al., 2011a,b). Злой жест тела имеет самые прямые последствия для наблюдателя «бей / беги», и, возможно, поэтому он привлек наибольшее внимание. Следовательно, в Эксперименте 2 эти требования к действию наиболее заметны в гневных жестах тела, показанных в нейтральном контексте, где угрожающий передний план явно выделяется из незаметной фоновой сцены. Мы считаем, что фиксации на теле были автоматическими, а не стратегическими, и поэтому лучше объясняются теорией мотивированного внимания.
Мы считаем, что фиксации на теле были автоматическими, а не стратегическими, и поэтому лучше объясняются теорией мотивированного внимания.
ЭМГ-ответы лица
 , 2007; Tamietto et al., 2009).). В недавнем исследовании лица участников были сняты на видео, когда они наблюдали изображения неоднозначных выражений лиц в контексте победы или поражения в спорте. Когда новые участники оценивали выражения лиц более ранних участников по валентности, оказалось, что выигрышный или проигрышный контекст смещал оценку участников в положительную или отрицательную сторону (Aviezer et al., 2012).
, 2007; Tamietto et al., 2009).). В недавнем исследовании лица участников были сняты на видео, когда они наблюдали изображения неоднозначных выражений лиц в контексте победы или поражения в спорте. Когда новые участники оценивали выражения лиц более ранних участников по валентности, оказалось, что выигрышный или проигрышный контекст смещал оценку участников в положительную или отрицательную сторону (Aviezer et al., 2012).
Расширение зрачка
 , 2010). Наши результаты согласуются с этими последними выводами. Действительно, наши зрачки расширяются в ответ на все эмоциональные сигналы, но усиленный эффект наблюдался именно после гневных сигналов, вызывающих немедленное возбуждение.
, 2010). Наши результаты согласуются с этими последними выводами. Действительно, наши зрачки расширяются в ответ на все эмоциональные сигналы, но усиленный эффект наблюдался именно после гневных сигналов, вызывающих немедленное возбуждение. Наконец, мы заметили, что выражения тела распознаются по-разному в зависимости от социальной сцены, в которой они были представлены. В целом, наши результаты показывают, что на реакцию наблюдателей на выражения лица влияют выражения всего тела и что последние воспринимаются на фоне окружающей социальной сцены. Показатели, которые до сих пор считались специфичными для просмотра отдельных выражений лица, чувствительны к естественным условиям просмотра лица. Мы показываем, что при столкновении с угрозой, исходящей от лица, тела или сцены, зрачки участников расширялись, их корругационная мышца становилась более активной, и они направляли взгляд на угрозу.
Наконец, мы заметили, что выражения тела распознаются по-разному в зависимости от социальной сцены, в которой они были представлены. В целом, наши результаты показывают, что на реакцию наблюдателей на выражения лица влияют выражения всего тела и что последние воспринимаются на фоне окружающей социальной сцены. Показатели, которые до сих пор считались специфичными для просмотра отдельных выражений лица, чувствительны к естественным условиям просмотра лица. Мы показываем, что при столкновении с угрозой, исходящей от лица, тела или сцены, зрачки участников расширялись, их корругационная мышца становилась более активной, и они направляли взгляд на угрозу. Вклад автора
Заявление о конфликте интересов

Благодарности

Ссылки
 S., Tikofsky, R.S., and Hyde, J.S. (1992). Временная динамика EPI функции человеческого мозга во время активации задачи. Маг. Резон. Мед . 25, 390–397. doi: 10.1002/mrm.10220
S., Tikofsky, R.S., and Hyde, J.S. (1992). Временная динамика EPI функции человеческого мозга во время активации задачи. Маг. Резон. Мед . 25, 390–397. doi: 10.1002/mrm.10220 Активация зрительной коры при мотивированном внимании. Поведение. Нейроски . 117, 369–380. doi: 10.1037/0735-7044.117.2.369
Активация зрительной коры при мотивированном внимании. Поведение. Нейроски . 117, 369–380. doi: 10.1037/0735-7044.117.2.369 doi: 10.1111/j.1469-8986.1982.tb02516.x
doi: 10.1111/j.1469-8986.1982.tb02516.x Познан. Оказывать воздействие. Поведение Нейроски . 10, 252–269. doi: 10.3758/CABN.10.2.252
Познан. Оказывать воздействие. Поведение Нейроски . 10, 252–269. doi: 10.3758/CABN.10.2.252
CrossRef Полный текст
Хаксби, Дж. В., Хоффман, Э. А., и Гоббини, М. И. (2000). Распределенная нейронная система человека для восприятия лица. Тенденции Познан. Наука . 4, 223–233. doi: 10.1016/S1364-6613(00)01482-0
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Гесс, У., и Фишер, А. (2013). Эмоциональная мимикрия как социальная регуляция. чел. соц. Психол. Версия . 17, 142–157. doi: 10.1177/1088868312472607
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Крет, М.Э., и де Гелдер, Б. (2010). Социальный контекст влияет на распознавание телесных выражений. Экспл. Мозг Res . 203, 169–180. doi: 10.1007/s00221-010-2220-8
Социальный контекст влияет на распознавание телесных выражений. Экспл. Мозг Res . 203, 169–180. doi: 10.1007/s00221-010-2220-8
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Крет, М. Э., и де Гелдер, Б. (2012a). Исламский головной убор влияет на то, как эмоции распознаются по глазам. Перед. Психол . 3:110. doi: 10.3389/fpsyg.2012.00110
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Крет, М. Э., и де Гелдер, Б. (2012b). Обзор половых различий в обработке эмоциональных сигналов. Нейропсихология 50, 1211–1221. doi: 10.1016/j.neuropsychologia.2011.12.022
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Крет, М. Э., и де Гелдер, Б. (2013). Когда улыбка превращается в кулак: восприятие мимических и телесных выражений эмоций у насильственных преступников. Экспл. Мозг Res . 228, 399–410. doi: 10.1007/s00221-013-3557-6
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Крет, М. Е., Пишон, С., Грез, Дж., и де Гелдер, Б. (2011a). Мужчины больше всего боятся других мужчин: активация мозга в зависимости от пола при восприятии угрозы от динамичных лиц и тел — исследование фМРТ. Фронт. Психол . 2:3. doi: 10.3389/fpsyg.2011.00003
Е., Пишон, С., Грез, Дж., и де Гелдер, Б. (2011a). Мужчины больше всего боятся других мужчин: активация мозга в зависимости от пола при восприятии угрозы от динамичных лиц и тел — исследование фМРТ. Фронт. Психол . 2:3. doi: 10.3389/fpsyg.2011.00003
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Крет, М. Е., Пишон, С., Грез, Дж., и де Гелдер, Б. (2011b). Сходства и различия в восприятии угрозы от динамических лиц и тел. ФМРТ-исследование. Нейроизображение 54, 1755–1762 гг. doi: 10.1016/j.neuroimage.2010.08.012
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Крет, М. Э., Денолле, Дж., Грез, Дж., и де Гелдер, Б. (2011c). Роль негативной аффективности и социального торможения в восприятии социальной угрозы: исследование фМРТ. Нейропсихология 49, 1187–1193. doi: 10.1016/j.neuropsychologia.2011.02.007
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Крет, М. Е., Стекеленбург, Дж.Дж., Рулофс, К., и де Гелдер, Б. (2013). Восприятие выражений лица и тела с помощью ЭМГ и показателей взгляда. Перед. Психол . 4:28. doi: 10.3389/fpsyg.2013.00028
Е., Стекеленбург, Дж.Дж., Рулофс, К., и де Гелдер, Б. (2013). Восприятие выражений лица и тела с помощью ЭМГ и показателей взгляда. Перед. Психол . 4:28. doi: 10.3389/fpsyg.2013.00028
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Ланг, П.Дж., и Катберт, Б.Н. (1997). «Мотивированное внимание: аффект, активация и действие», в Attention and Orienting , eds PJ Lang, RF Simons, and M. Balabon (Mahwah, NJ: Erlbaum), 97–135.
Лундквист Д. и Оман А. (2005). Эмоции регулируют внимание: отношение между формами лица, эмоциями лица и зрительным вниманием. Виз. Код . 12, 51–84. doi: 10.1080/13506280444000085
CrossRef Полный текст
Magnee, M.J., Stekelenburg, J.J., Kemner, C., and de Gelder, B. (2007). Сходные электромиографические реакции лица на лица, голоса и мимику тела. Нейроотчет 18, 369–372. doi: 10.1097/WNR.0b013e32801776e6
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Мирен, Х. К., ван Хейнсберген, К.С., и де Гелдер, Б. (2005). Быстрая перцептивная интеграция выражения лица и эмоционального языка тела. Проц. Натл. акад. науч. США . 102, 16518–16523. doi: 10.1073/pnas.0507650102
К., ван Хейнсберген, К.С., и де Гелдер, Б. (2005). Быстрая перцептивная интеграция выражения лица и эмоционального языка тела. Проц. Натл. акад. науч. США . 102, 16518–16523. doi: 10.1073/pnas.0507650102
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Мондлох, С.Дж., Нельсон, Н.Л., и Хорнер, М. (2013). Асимметрии влияния: дифференциальное влияние поз тела на восприятие эмоциональных выражений лица. PLoS ONE 8:e73605. doi: 10.1371/journal.pone.0073605
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Оман А., Лундквист Д. и Эстевес Ф. (2001). Снова лицо в толпе: преимущество угрозы со схематическими стимулами. Дж. Перс. соц. Психол . 80, 381–396. doi: 10.1037/0022-3514.80.3.381
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Овербик, Т.Дж., ван Бокстель, А., и Вестеринк, Дж.Х. (2012). Реакция респираторной синусовой аритмии на индуцированные эмоциональные состояния: влияние индексов RSA, метода индукции эмоций, возраста и пола. биол. Психол . 91, 128–141. doi: 10.1016/j.biopsycho.2012.05.011
биол. Психол . 91, 128–141. doi: 10.1016/j.biopsycho.2012.05.011
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Ригхарт, Р., и де Гелдер, Б. (2006). Контекст влияет на ранний перцептивный анализ лиц — электрофизиологическое исследование. Церебр. Кора 16, 1249–1257. doi: 10.1093/cercor/bhj066
Pubmed Abstract | Опубликован полный текст | Полный текст CrossRef
Righart, R., and de Gelder, B. (2008a). Быстрое влияние эмоциональных сцен на кодирование выражений лица: исследование ERP. Соц. Познан. Оказывать воздействие. Нейроски . 3, 270–278. doi: 10.1093/scan/nsn021
Опубликовано Аннотация | Опубликован полный текст | Полный текст CrossRef
Ригхарт Р. и де Гелдер Б. (2008b). Эмоциональная суть сцены влияет на распознавание выражений лица. Познан. Оказывать воздействие. Поведение Нейроски . 8, 264–272. doi: 10.3758/CABN.8.3.264
Опубликовано Аннотация | Опубликован полный текст | CrossRef Full Text
Tamietto, M. , Castelli, L., Vighetti, S., Perozzo, P., Geminiani, G., Weiskrantz, L., et al. (2009 г.). Невидимые выражения лица и тела вызывают быстрые эмоциональные реакции. Проц. Натл. акад. науч. США . 106, 17661–17666. doi: 10.1073/pnas.0
, Castelli, L., Vighetti, S., Perozzo, P., Geminiani, G., Weiskrantz, L., et al. (2009 г.). Невидимые выражения лица и тела вызывают быстрые эмоциональные реакции. Проц. Натл. акад. науч. США . 106, 17661–17666. doi: 10.1073/pnas.0
Pubmed Abstract | Опубликован полный текст | CrossRef Полный текст
Тоттенхэм, Н., Танака, Дж. В., Леон, А. К., МакКэрри, Т., Нерс, М., Хэйр, Т. А., и др. (2009). Набор выражений лица NimStim: суждения неподготовленных участников исследования. Психиатрия Рез . 168, 242–249. doi: 10.1016/j.psychres.2008.05.006
Опубликовано Резюме | Опубликован полный текст | Полный текст CrossRef
Ван Бокстель, А. (2010). «ЭМГ лица как инструмент для определения аффективных состояний», в Proceedings of Measuring Behavior , eds F. Grieco, A.J. Spink, O. Krips., L. Loijens, L. Noldus и P. Zimmerman (Wageningen: Noldus Information Технология), 104–108.
Ван ден Сток, Дж., Ригхарт, Р., и де Гелдер, Б. (2007). Выражение тела влияет на распознавание эмоций по лицу и голосу. Эмоция 7, 487–494. doi: 10.1037/1528-3542.7.3.487
Выражение тела влияет на распознавание эмоций по лицу и голосу. Эмоция 7, 487–494. doi: 10.1037/1528-3542.7.3.487
Опубликовано Аннотация | Опубликован полный текст | CrossRef Полный текст
Увеличение передачи сигналов анандамидом спасает синаптическую функцию миндалины и сопутствующие эмоциональные изменения в модели эпилепсии
ПредыдущийСледующий G. Campbell Teskey
Journal of Neuroscience 29 июля 2020 г., 40 (31) 6068-6081; DOI: https://doi.org/10.1523/JNEUROSCI.0068-20.2020
- Статья
- Рисунки и данные
- Info & Metrics
- Электроны
Аннотация
. . Таким образом, стойкое изменение передачи сигналов eCB после повторных припадков может способствовать развитию эмоциональных расстройств, связанных с эпилепсией. Здесь мы сообщаем, что повторное возникновение припадков (зажигание) в миндалевидном теле вызывало долгосрочное усиление тревоги и нарушение сохранения памяти о страхе, что сопровождалось дисбалансом пресинаптической активности ГАМК/глутамата и изменением синаптической пластичности в базолатеральной миндалевидном теле (БЛА). ), у самцов крыс. Содержание анандамида (AEA) снижалось после повторных припадков, а фармакологическое усиление передачи сигналов AEA избавляло от судорожной тревоги, восстанавливая тонический контроль передачи сигналов eCB над глутаматергической передачей. Более того, усиление передачи сигналов AEA также спасало вызванные припадками изменения памяти о страхе, восстанавливая фазовый контроль передачи сигналов eCB над ГАМКергической активностью и пластичностью в BLA. Эти результаты показывают, что модуляция передачи сигналов AEA представляет собой потенциальную и многообещающую цель для лечения сопутствующей эмоциональной дисфункции, связанной с эпилепсией.
), у самцов крыс. Содержание анандамида (AEA) снижалось после повторных припадков, а фармакологическое усиление передачи сигналов AEA избавляло от судорожной тревоги, восстанавливая тонический контроль передачи сигналов eCB над глутаматергической передачей. Более того, усиление передачи сигналов AEA также спасало вызванные припадками изменения памяти о страхе, восстанавливая фазовый контроль передачи сигналов eCB над ГАМКергической активностью и пластичностью в BLA. Эти результаты показывают, что модуляция передачи сигналов AEA представляет собой потенциальную и многообещающую цель для лечения сопутствующей эмоциональной дисфункции, связанной с эпилепсией. ЗНАЧИМОСТЬ Эпилепсия представляет собой гетерогенное неврологическое расстройство, обычно связанное с сопутствующими эмоциональными изменениями. Однако лечение эпилепсии обычно ограничивается контролем приступов. Эндоканнабиноидная (eCB) система, особенно передача сигналов анандамида (AEA), контролирует возбудимость нейронов и экспрессию судорог, а также регулирует эмоциональное поведение.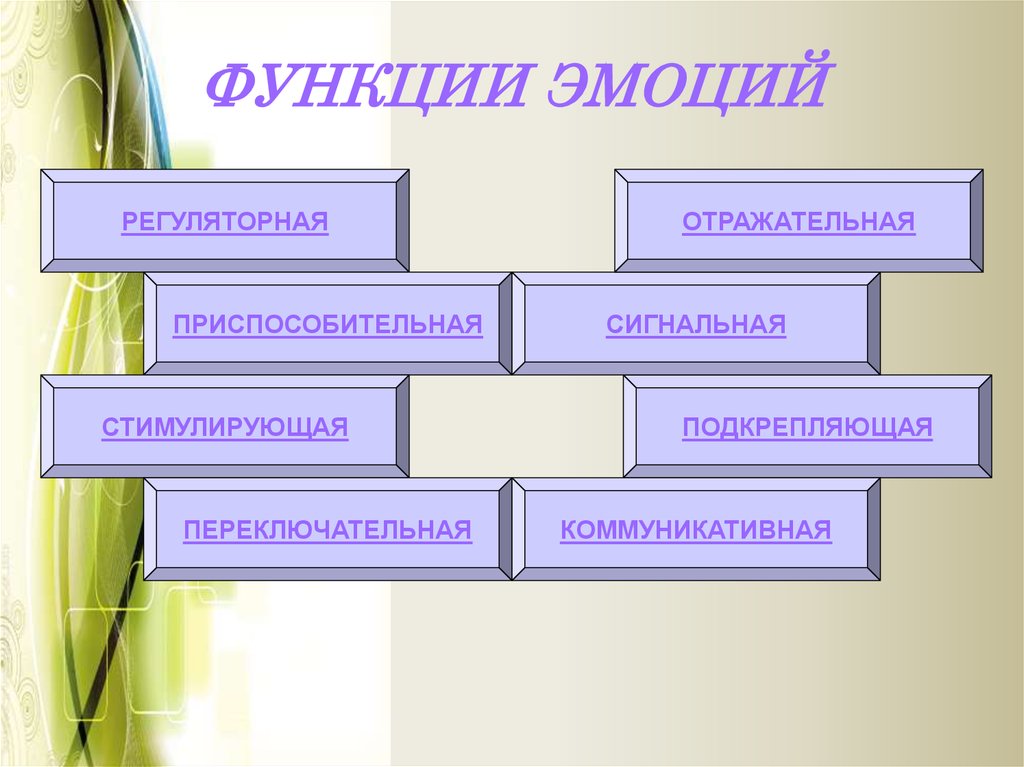 Мы обнаружили, что повторяющиеся припадки вызывают аллостатическую дезадаптацию передачи сигналов АЕА в миндалевидном теле, которая вызывает эмоциональные изменения. Усиление передачи сигналов AEA за счет ингибирования его деградирующего фермента, гидролазы амидов жирных кислот (FAAH), восстанавливало как синаптические, так и поведенческие изменения. Ингибиторы FAAH ослабляют судорожную активность в моделях на животных и используются в клинических исследованиях для лечения негативных последствий, связанных со стрессом. Таким образом, они доступны и могут быть клинически оценены для лечения как судорог, так и коморбидных состояний, связанных с эпилепсией.
Мы обнаружили, что повторяющиеся припадки вызывают аллостатическую дезадаптацию передачи сигналов АЕА в миндалевидном теле, которая вызывает эмоциональные изменения. Усиление передачи сигналов AEA за счет ингибирования его деградирующего фермента, гидролазы амидов жирных кислот (FAAH), восстанавливало как синаптические, так и поведенческие изменения. Ингибиторы FAAH ослабляют судорожную активность в моделях на животных и используются в клинических исследованиях для лечения негативных последствий, связанных со стрессом. Таким образом, они доступны и могут быть клинически оценены для лечения как судорог, так и коморбидных состояний, связанных с эпилепсией.
- AMYDALA
- ANANDAMIDE
- Эмоциональное поведение
- Эпилепсия
- FAAAH
- СИНАПТИЧЕСКИЙ ПЛАСТИЧЕСКИЙ ПЛАСТИЧЕСКИЙ
ВВЕДЕНИЕ
66666666 гг. Эти сопутствующие заболевания включают депрессию, тревожные расстройства и нарушения памяти, которые сильно влияют на качество жизни (Kanner et al. , 2010). Височная эпилепсия (ВЭЛ), наиболее распространенная форма фокальной эпилепсии у людей, характеризуется повторяющимися самопроизвольными разрядами, возникающими из височных областей, и часто связана с межприступной поведенческой дисфункцией (Kanner, 2016). Наиболее часто назначаемые противоэпилептические препараты снижают частоту или тяжесть приступов, но не оказывают никакого эффекта или даже могут усугубить эмоциональные нарушения и нарушения памяти (Eddy et al., 2011).
, 2010). Височная эпилепсия (ВЭЛ), наиболее распространенная форма фокальной эпилепсии у людей, характеризуется повторяющимися самопроизвольными разрядами, возникающими из височных областей, и часто связана с межприступной поведенческой дисфункцией (Kanner, 2016). Наиболее часто назначаемые противоэпилептические препараты снижают частоту или тяжесть приступов, но не оказывают никакого эффекта или даже могут усугубить эмоциональные нарушения и нарушения памяти (Eddy et al., 2011).
Миндалевидное тело является ключевой структурой мозга, участвующей в приписывании эмоциональной валентности внешним раздражителям (LeDoux, 2000), а также генерирует и распространяет судорожную активность (Aroniadou-Anderjaska et al., 2008). Доклинические исследования показали изменения эмоционального поведения после многократно вызванных приступов миндалины (Botterill et al., 2014; Chen et al., 2016). Эндоканнабиноидная (eCB) система представляет собой нейромодулирующую систему в головном мозге, которая точно формирует возбудимость нейронов и синаптическую пластичность (Chevaleyre et al. , 2006; Kano et al., 2009).). Система eCB сильно активируется во время судорог (Stella et al., 1997), и как экзогенные, так и эндогенные каннабиноиды резко ослабляют повышенную возбудимость нейронов и экспрессию судорог (Marsicano et al., 2003; Wallace et al., 2003; Colangeli et al., 2017, 2019). Система eCB состоит из рецепторов каннабиноидов типа 1 (CB1) и каннабиноидов типа 2 (CB2), двух основных эндогенных каннабиноидов анандамида (AEA) и 2-арахидоноилглицерина (2-AG), а также ферментов, ответственных за их синтез и деградацию. (Кано и др., 2009 г.). Сообщалось об изменениях молекулярных элементов eCB в головном мозге как животных с моделями эпилепсии, так и людей с височной эпилепсией (Ludányi et al., 2008; Di Maio et al., 2019), а в спинномозговой жидкости наблюдалось снижение уровня AEA. людей с височной эпилепсией (Romigi et al., 2010). Более того, усиление передачи сигналов AEA восстанавливает индуцированное судорогами нарушение кратковременной и долговременной синаптической пластичности в зубчатой извилине после киндлинга перфорантного пути у крыс (Colangeli et al.
, 2006; Kano et al., 2009).). Система eCB сильно активируется во время судорог (Stella et al., 1997), и как экзогенные, так и эндогенные каннабиноиды резко ослабляют повышенную возбудимость нейронов и экспрессию судорог (Marsicano et al., 2003; Wallace et al., 2003; Colangeli et al., 2017, 2019). Система eCB состоит из рецепторов каннабиноидов типа 1 (CB1) и каннабиноидов типа 2 (CB2), двух основных эндогенных каннабиноидов анандамида (AEA) и 2-арахидоноилглицерина (2-AG), а также ферментов, ответственных за их синтез и деградацию. (Кано и др., 2009 г.). Сообщалось об изменениях молекулярных элементов eCB в головном мозге как животных с моделями эпилепсии, так и людей с височной эпилепсией (Ludányi et al., 2008; Di Maio et al., 2019), а в спинномозговой жидкости наблюдалось снижение уровня AEA. людей с височной эпилепсией (Romigi et al., 2010). Более того, усиление передачи сигналов AEA восстанавливает индуцированное судорогами нарушение кратковременной и долговременной синаптической пластичности в зубчатой извилине после киндлинга перфорантного пути у крыс (Colangeli et al. , 2017).
, 2017).
Передача сигналов eCB является важным регулятором тревожноподобного поведения и памяти о страхе у грызунов (Bedse et al., 2014; Morena et al., 2016a). Нарушение передачи сигналов eCB в миндалевидном теле вызывает тревожно-подобное поведение (Haller et al., 2004) и вызывает изменения динамики памяти о страхе у грызунов (Marsicano et al., 2002). Поэтому мы проверили гипотезу о том, что повторные приступы миндалины вызывают дисфункцию эмоционального поведения за счет стойкого неадекватного изменения передачи сигналов eCB в базолатеральной миндалине (БЛА).
Материалы и методы
Животные
Самцов крыс Long-Evans (LE) с капюшоном (LE) (300–350 г; Charles River) в начале поведенческих или электрофизиологических экспериментов помещали индивидуально в термостатируемую (23 ± 1°C) камеру. ) помещение вивария и поддерживается в определенных условиях отсутствия патогенов и 12/12-часовом цикле свет/темнота (с 7:00 до 19:00 свет включен). Еда и вода были доступны вволю . Все тесты проводились во время легкой фазы цикла. Все экспериментальные процедуры соответствовали протоколам, одобренным Комитетом по уходу за животными Университета Калгари, и рекомендациям Канадского совета по уходу за животными. Были предприняты все усилия, чтобы свести к минимуму страдания животных и сократить количество используемых животных.
Все тесты проводились во время легкой фазы цикла. Все экспериментальные процедуры соответствовали протоколам, одобренным Комитетом по уходу за животными Университета Калгари, и рекомендациям Канадского совета по уходу за животными. Были предприняты все усилия, чтобы свести к минимуму страдания животных и сократить количество используемых животных.
Хирургия
Постоянно имплантированные электроды для процедуры растопки были изготовлены из проволоки из нержавеющей стали с тефлоновым покрытием диаметром 178 мкм (AM Systems). Концы проводов были очищены от тефлона и соединены с позолоченными амфенольными контактами. Крыс (как ложных, так и разожженных) анестезировали 5% изофлураном и поддерживали от 1% до 2%. Затем животных помещали в стереотаксическую раму (David Kopf Instruments) и один биполярный электрод хронически имплантировали в правую БЛА (координаты: переднезадняя, -2,8 мм, и медиолатеральная, -4,8 мм от брегмы; дорсовентральная, -8,0 мм от твердой мозговой оболочки). ). Электроды были прикреплены к черепу с помощью стоматологического цемента и трех винтов из нержавеющей стали. Один из трех винтов служил заземляющим электродом. Крысам давали бупренорфин, и им давали возможность восстановиться после операции в течение одной недели до начала процедур растопки.
). Электроды были прикреплены к черепу с помощью стоматологического цемента и трех винтов из нержавеющей стали. Один из трех винтов служил заземляющим электродом. Крысам давали бупренорфин, и им давали возможность восстановиться после операции в течение одной недели до начала процедур растопки.
Лекарства
Ингибитор гидролиза AEA URB597 (0,5 мг/кг; Cayman Chemical) или его носитель (5% полиэтиленгликоль, 5% Tween 80 и 90% физиологический раствор) вводили внутрибрюшинно (2 мл/кг), 60 мин до эксперимента с приподнятым крестообразным лабиринтом (EPM), или сразу после сеанса кондиционирования страха, или за 60 мин до подготовки срезов к электрофизиологическим экспериментам. Доза была выбрана на основе ранее опубликованных работ (Kathuria et al., 2003) и пилотных экспериментов, проведенных в нашей лаборатории. Все растворы лекарственных средств были свежеприготовлены перед каждым экспериментом.
Экспериментальный план и методика
Процедура разжигания
Процедура разжигания состояла из 20 электрических стимуляций правой миндалины (1 мс, двухфазные прямоугольные импульсы, 60 Гц в течение 1 с) с током стимуляции 20% выше начального порога послеразряда (Farrell et al. , 2016). Электрическая стимуляция проводилась два раза в день в течение 10 дней с перерывом не менее 4 часов между стимуляциями, чтобы рассеять постиктальные эффекты (Farrell et al., 2016, 2017). Так же, как и пламенных крыс, ложных животных помещали в клетку для растопки два раза в день на 10 дней и подключали к стимулятору, но ток не подавали. Поведенческие судороги, вызванные каждой стимуляцией, оценивали по шкале Расина (Racine, 1972). Классы были оперативно определены как класс 1: неподвижность; класс 2: орофациальные автоматизмы с киванием головой; класс 3: односторонний клонус передних конечностей; класс 4: выращивание с двусторонним клонусом передних конечностей; и класс 5: поднимание с двусторонним клонусом передних конечностей с последующим падением. При использовании этой системы классификации крысы считаются «возбужденными» после трех последовательных судорог 5-го класса (Pinel and Rovner, 1978).
, 2016). Электрическая стимуляция проводилась два раза в день в течение 10 дней с перерывом не менее 4 часов между стимуляциями, чтобы рассеять постиктальные эффекты (Farrell et al., 2016, 2017). Так же, как и пламенных крыс, ложных животных помещали в клетку для растопки два раза в день на 10 дней и подключали к стимулятору, но ток не подавали. Поведенческие судороги, вызванные каждой стимуляцией, оценивали по шкале Расина (Racine, 1972). Классы были оперативно определены как класс 1: неподвижность; класс 2: орофациальные автоматизмы с киванием головой; класс 3: односторонний клонус передних конечностей; класс 4: выращивание с двусторонним клонусом передних конечностей; и класс 5: поднимание с двусторонним клонусом передних конечностей с последующим падением. При использовании этой системы классификации крысы считаются «возбужденными» после трех последовательных судорог 5-го класса (Pinel and Rovner, 1978).
EPM
Через неделю после последнего сеанса растопки крыс тестировали на тревожное поведение в EPM. Перед тестированием ЭПМ животных брали на руки по 1 мин в течение трех дней подряд. Для приучения животных к комнате поведенческого тестирования крыс переводили в поведенческую комнату, а их домашние клетки помещали в звукоизолирующие, вентилируемые и освещаемые шкафы не менее чем на 30 мин до и после обработки и на 9 мин.0 мин до тестирования EPM. Аппарат EPM (Med Associates Inc.) состоял из двух открытых рукавов (50 × 10 × 75 см 3 ) и двух закрытых рукавов (50 × 10 × 75 см 3 ), которые выходили из общей центральной платформы (10 × 10 см 2 ). Лабиринт располагался в звукоизолированной комнате в условиях красного освещения (открытые рукава, 15 лк; закрытые рукава, 5 лк). Была использована слегка модифицированная процедура, описанная нами ранее (Morena et al., 2016b, 2019). Каждую крысу помещали в центральную часть аппарата головой к закрытому рукаву. Воздействие длилось 5 минут, и поведение регистрировалось с помощью видеокамеры, расположенной над экспериментальной установкой, и анализировалось с помощью программного обеспечения Noldus EthoVision XT (RRID: SCR_000441).
Перед тестированием ЭПМ животных брали на руки по 1 мин в течение трех дней подряд. Для приучения животных к комнате поведенческого тестирования крыс переводили в поведенческую комнату, а их домашние клетки помещали в звукоизолирующие, вентилируемые и освещаемые шкафы не менее чем на 30 мин до и после обработки и на 9 мин.0 мин до тестирования EPM. Аппарат EPM (Med Associates Inc.) состоял из двух открытых рукавов (50 × 10 × 75 см 3 ) и двух закрытых рукавов (50 × 10 × 75 см 3 ), которые выходили из общей центральной платформы (10 × 10 см 2 ). Лабиринт располагался в звукоизолированной комнате в условиях красного освещения (открытые рукава, 15 лк; закрытые рукава, 5 лк). Была использована слегка модифицированная процедура, описанная нами ранее (Morena et al., 2016b, 2019). Каждую крысу помещали в центральную часть аппарата головой к закрытому рукаву. Воздействие длилось 5 минут, и поведение регистрировалось с помощью видеокамеры, расположенной над экспериментальной установкой, и анализировалось с помощью программного обеспечения Noldus EthoVision XT (RRID: SCR_000441). В качестве индикаторов тревожноподобного поведения и двигательной активности анализировали следующие параметры: процент времени, проведенного в открытых рукавах (время открытого %), выраженное как [(секунд, проведенных в открытых ответвлениях лабиринта/секунд, проведенных в открытых + закрытых оружия) × 100]; процент записей в открытых рукавах (открытые входы %), выраженный как [(количество входов в открытые рукава/количество входов в открытые + закрытые рукава) × 100]; общее пройденное расстояние, выраженное в сантиметрах (расстояние). После каждого использования лабиринт очищали 70% раствором этанола.
В качестве индикаторов тревожноподобного поведения и двигательной активности анализировали следующие параметры: процент времени, проведенного в открытых рукавах (время открытого %), выраженное как [(секунд, проведенных в открытых ответвлениях лабиринта/секунд, проведенных в открытых + закрытых оружия) × 100]; процент записей в открытых рукавах (открытые входы %), выраженный как [(количество входов в открытые рукава/количество входов в открытые + закрытые рукава) × 100]; общее пройденное расстояние, выраженное в сантиметрах (расстояние). После каждого использования лабиринт очищали 70% раствором этанола.
Слуховое формирование страха
Через неделю после последнего сеанса растопки отдельные когорты животных использовали для экспериментов по слуховому условию страха с использованием слегка измененной ранее описанной процедуры (Morena et al., 2019). Для приучения животных к комнате поведенческого тестирования крыс переводили в поведенческую комнату, а их домашние клетки помещали в звукоизолирующие, вентилируемые и освещаемые шкафы не менее чем на 30 мин до и после манипуляций в 1-й и 2-й дни, а в течение 90 мин до и после тестирования, последующие дни. Крыс обрабатывали в течение 1 мин с каждой. В 1-й и 2-й дни сразу после процедуры хендлинга животных приучали к камерам кондиционирования страха (контекст А, 1-й день и контекст Б, 2-й день) в течение 10 мин. Слуховое обусловливание страха (день 3) проводили в контексте А. После 5-минутного периода акклиматизации все крысы подвергались трем испытаниям на обусловливание. Каждая проба на обусловливание включала предъявление условного стимула (УС; 80 дБ, тон 4 Гц) в течение 30 с, одновременно заканчивающееся односекундным безусловным стимулом (УЗ; 0,9).удар током 0 мА). Межпробный интервал (ITI) между двумя последовательными парами CS-US составлял 3 минуты. После кондиционирования каждую крысу возвращали в ее домашнюю клетку. На 4-й день крысы получали 24-часовой сеанс сохранения памяти, состоящий из 18 презентаций CS с ITI между CS 2 мин в контексте B. На 5-й день крысы получали 48-часовой сеанс удержания памяти в контексте B, состоящий из 5 CS (2-минутный ITI). Камеры кондиционирования страха (30,5 × 24,1 × 21 см 3 ) были оборудованы настилом из металлических стержней из нержавеющей стали, соединенным с генератором тока.
Крыс обрабатывали в течение 1 мин с каждой. В 1-й и 2-й дни сразу после процедуры хендлинга животных приучали к камерам кондиционирования страха (контекст А, 1-й день и контекст Б, 2-й день) в течение 10 мин. Слуховое обусловливание страха (день 3) проводили в контексте А. После 5-минутного периода акклиматизации все крысы подвергались трем испытаниям на обусловливание. Каждая проба на обусловливание включала предъявление условного стимула (УС; 80 дБ, тон 4 Гц) в течение 30 с, одновременно заканчивающееся односекундным безусловным стимулом (УЗ; 0,9).удар током 0 мА). Межпробный интервал (ITI) между двумя последовательными парами CS-US составлял 3 минуты. После кондиционирования каждую крысу возвращали в ее домашнюю клетку. На 4-й день крысы получали 24-часовой сеанс сохранения памяти, состоящий из 18 презентаций CS с ITI между CS 2 мин в контексте B. На 5-й день крысы получали 48-часовой сеанс удержания памяти в контексте B, состоящий из 5 CS (2-минутный ITI). Камеры кондиционирования страха (30,5 × 24,1 × 21 см 3 ) были оборудованы настилом из металлических стержней из нержавеющей стали, соединенным с генератором тока. Каждая камера была заключена в вентилируемые и шумопоглощающие боксы (Med Associates Inc.). Были использованы два разных контекста (контексты A и B). Контекст А состоял из камеры с решетчатым полом, задней и боковыми металлическими стенами, передней дверью и потолком из прозрачного плексигласа и белым светом. Контекст А очищали 70% этанолом между крысами. Чтобы создать новый контекст тестирования (контекст B), первоначальный решетчатый пол был покрыт белой непрозрачной пластиковой вставкой, а боковые металлические стенки были закрыты белой пластиковой панелью, чтобы создать изогнутую стену. Контекст Б очищали раствором Виркона между крысами. Видеокамера, расположенная на внутренней двери бокса для кондиционирования, записывала видео, и полученный результат анализировали на предмет замирания (т. е. отсутствия движений, кроме дыхания) с использованием программного обеспечения Video Freeze (Med Associates Inc.; RRID: SCR_014574). .
Каждая камера была заключена в вентилируемые и шумопоглощающие боксы (Med Associates Inc.). Были использованы два разных контекста (контексты A и B). Контекст А состоял из камеры с решетчатым полом, задней и боковыми металлическими стенами, передней дверью и потолком из прозрачного плексигласа и белым светом. Контекст А очищали 70% этанолом между крысами. Чтобы создать новый контекст тестирования (контекст B), первоначальный решетчатый пол был покрыт белой непрозрачной пластиковой вставкой, а боковые металлические стенки были закрыты белой пластиковой панелью, чтобы создать изогнутую стену. Контекст Б очищали раствором Виркона между крысами. Видеокамера, расположенная на внутренней двери бокса для кондиционирования, записывала видео, и полученный результат анализировали на предмет замирания (т. е. отсутствия движений, кроме дыхания) с использованием программного обеспечения Video Freeze (Med Associates Inc.; RRID: SCR_014574). .
Электрофизиология
Через неделю после последнего вызванного приступа крыс декапитировали под глубокой анестезией изофлураном и транскардиально перфузировали ледяным раствором для срезов, содержащим следующее: 87 мМ NaCl, 2,5 мМ KCl, 25 мМ NaHCO 3 , 0,5 мМ CaCl 2 , 7 мМ MgCl 2 , 1,25 мМ NaH 2 PO 4 , 25 мМ D-глюкозы и 75 мМ сахарозы. Коронарные срезы головного мозга БЛА (толщиной 300 мкм) были приготовлены с помощью слайсера Leica VT1000S (RRID: SCR_01649).5) в ледяном растворе для нарезки. Для восстановления срезы инкубировали в камере для выдержки с нормальной искусственной спинномозговой жидкостью (ACSF), содержащей следующее: 126 мМ NaCl, 2,5 мМ KCl, 2,5 мМ CaCl 2 , 1 мМ MgSO 4 , 1,25 мМ NaH 2 PO 4 , 26 мМ NaHCO 3 и 10 мМ глюкозы при рН 7,4 в течение 30 мин при 32°С, а затем при комнатной температуре не менее 30 мин. После инкубации срезы помещали в регистрационную камеру и перфузировали ACSF со скоростью потока 1,5 мл/мин. Во всех электрофизиологических экспериментах использовалась только левая сторона БЛА, противоположная стороне имплантации. Все эксперименты проводили при комнатной температуре (22–25°С). Для записей пэтч-клэмп целых клеток основные нейроны BLA отличались от интернейронов в соответствии с их морфологическими и электрофизиологическими свойствами (Washburn and Moises, 19).
Коронарные срезы головного мозга БЛА (толщиной 300 мкм) были приготовлены с помощью слайсера Leica VT1000S (RRID: SCR_01649).5) в ледяном растворе для нарезки. Для восстановления срезы инкубировали в камере для выдержки с нормальной искусственной спинномозговой жидкостью (ACSF), содержащей следующее: 126 мМ NaCl, 2,5 мМ KCl, 2,5 мМ CaCl 2 , 1 мМ MgSO 4 , 1,25 мМ NaH 2 PO 4 , 26 мМ NaHCO 3 и 10 мМ глюкозы при рН 7,4 в течение 30 мин при 32°С, а затем при комнатной температуре не менее 30 мин. После инкубации срезы помещали в регистрационную камеру и перфузировали ACSF со скоростью потока 1,5 мл/мин. Во всех электрофизиологических экспериментах использовалась только левая сторона БЛА, противоположная стороне имплантации. Все эксперименты проводили при комнатной температуре (22–25°С). Для записей пэтч-клэмп целых клеток основные нейроны BLA отличались от интернейронов в соответствии с их морфологическими и электрофизиологическими свойствами (Washburn and Moises, 19). 92). Эксперименты по основным внутренним свойствам, регистрации ЭПСК и соотношению ингибирования/возбуждения (И-Э) проводили с электродами из боросиликатного стекла (4–6 МОм), заполненными стандартным внутриклеточным раствором следующего состава: 108 мМ K-глюконата, 8 мМ Na-глюконата, 2 мМ MgCl 2 , 8 мМ KCl, 4 мМ K2-ATP, 0,3 мМ Na-GTP, 1 мМ EGTA и 10 мМ HEPES, рН 7,4, отрегулированный KOH. ИПСК, опосредованные рецептором ГАМК-А, регистрировали с помощью стеклянных электродов (3–5 МОм), заполненных раствором, содержащим следующее: 90 мм CsCH 3 SO 3 , 50 мм CsCl, 1 мм EGTA, 10 мм HEPES, 4,6 мм MgCl 2 , 0,1 мм CaCl 2 , 5 мм QX314, 0,3 мм 4 мм Na-GTP и Mg-АТФ, рН 7,3, доведенный до CsOH. 40 мкм d-(-)-2-амино-5-фосфонопентановой кислоты (d-AP-5) и 20 мкм динатрия 6,7-динитрохиноксалин-2,3-диона (DNQX) добавляли в ванну для блокирования глутаматергическая передача. В экспериментах IE IPSC были зарегистрированы с использованием стандартного внутриклеточного раствора, позволяющего регистрировать как положительно направленные IPSC, так и отрицательные EPSC в одной и той же клетке в течение одного и того же эксперимента, где мы не могли фармакологически изолировать передачу глутамата или ГАМК.
92). Эксперименты по основным внутренним свойствам, регистрации ЭПСК и соотношению ингибирования/возбуждения (И-Э) проводили с электродами из боросиликатного стекла (4–6 МОм), заполненными стандартным внутриклеточным раствором следующего состава: 108 мМ K-глюконата, 8 мМ Na-глюконата, 2 мМ MgCl 2 , 8 мМ KCl, 4 мМ K2-ATP, 0,3 мМ Na-GTP, 1 мМ EGTA и 10 мМ HEPES, рН 7,4, отрегулированный KOH. ИПСК, опосредованные рецептором ГАМК-А, регистрировали с помощью стеклянных электродов (3–5 МОм), заполненных раствором, содержащим следующее: 90 мм CsCH 3 SO 3 , 50 мм CsCl, 1 мм EGTA, 10 мм HEPES, 4,6 мм MgCl 2 , 0,1 мм CaCl 2 , 5 мм QX314, 0,3 мм 4 мм Na-GTP и Mg-АТФ, рН 7,3, доведенный до CsOH. 40 мкм d-(-)-2-амино-5-фосфонопентановой кислоты (d-AP-5) и 20 мкм динатрия 6,7-динитрохиноксалин-2,3-диона (DNQX) добавляли в ванну для блокирования глутаматергическая передача. В экспериментах IE IPSC были зарегистрированы с использованием стандартного внутриклеточного раствора, позволяющего регистрировать как положительно направленные IPSC, так и отрицательные EPSC в одной и той же клетке в течение одного и того же эксперимента, где мы не могли фармакологически изолировать передачу глутамата или ГАМК. Амплитуды и частоты спонтанных IPSC (sIPSC) и спонтанных EPSC (sEPSC) были обнаружены путем непрерывной записи в течение 300 с с использованием программного обеспечения Mini Analysis 6.0.7 (Synaptosoft) с использованием пороговых значений, в пять раз превышающих среднеквадратичное значение уровней шума. Нейроны удерживали при -60 мВ для записи EPSC и при -70 мВ для записи IPSC. Для парадигмы IE вызванные PSC (ePSC) были вызваны в режиме фиксации напряжения путем изменения мембранного потенциала зарегистрированного нейрона для выделения токов, связанных с глутаматергическими или ГАМКергическими токами. eEPSC были зарегистрированы путем фиксации напряжения клетки на уровне -60 мВ, когда движущая сила тормозного компонента эффективно устраняется, оставляя только токи возбуждения. Внешние eIPSC были зарегистрированы с той же настройкой стимуляции, что и для eEPSC, путем ограничения напряжения клетки до 0 мВ, где вклад AMPA-опосредованных PSCs незначителен. Затем было рассчитано и проанализировано сравнение соответствующих токов при каждом из этих потенциалов (отношение амплитуд eIPSCS/eEPSC).
Амплитуды и частоты спонтанных IPSC (sIPSC) и спонтанных EPSC (sEPSC) были обнаружены путем непрерывной записи в течение 300 с с использованием программного обеспечения Mini Analysis 6.0.7 (Synaptosoft) с использованием пороговых значений, в пять раз превышающих среднеквадратичное значение уровней шума. Нейроны удерживали при -60 мВ для записи EPSC и при -70 мВ для записи IPSC. Для парадигмы IE вызванные PSC (ePSC) были вызваны в режиме фиксации напряжения путем изменения мембранного потенциала зарегистрированного нейрона для выделения токов, связанных с глутаматергическими или ГАМКергическими токами. eEPSC были зарегистрированы путем фиксации напряжения клетки на уровне -60 мВ, когда движущая сила тормозного компонента эффективно устраняется, оставляя только токи возбуждения. Внешние eIPSC были зарегистрированы с той же настройкой стимуляции, что и для eEPSC, путем ограничения напряжения клетки до 0 мВ, где вклад AMPA-опосредованных PSCs незначителен. Затем было рассчитано и проанализировано сравнение соответствующих токов при каждом из этих потенциалов (отношение амплитуд eIPSCS/eEPSC). Все эксперименты проводились при постоянном присутствии AP-5 в ванне для блокирования токов, опосредованных NMDA. Емкость ячейки и сопротивление доступа (исходное значение <25 МОм) контролировали во время экспериментов, и записи принимались для анализа, если какая-либо переменная не изменялась более чем на 20%. Регистрацию потенциала внеклеточного поля проводили с помощью стеклянных микроэлектродов (1–2 МОм), заполненных ACSF. Для стимуляции предполагаемых глутаматергических или ГАМКергических волокон использовались монополярные пипетки пластыря, заполненные ACSF (1 МОм), помещенные в BLA непосредственно медиальнее внешней капсулы. Долговременную депрессию тормозной передачи (iLTD) индуцировали путем подачи тормозной низкочастотной стимуляции (iLFS), состоящей из 100 стимулов с частотой 1 Гц, через стимулирующий электрод. Для высокочастотной стимуляции (ВЧС) в латеральную миндалину (ЛП) применяли четыре серии по 100 импульсов с частотой 100 Гц и межстимульным интервалом 20 с. Для регистрации потенциала внеклеточного поля интенсивность стимула регулировали для получения полумаксимальных ответов.
Все эксперименты проводились при постоянном присутствии AP-5 в ванне для блокирования токов, опосредованных NMDA. Емкость ячейки и сопротивление доступа (исходное значение <25 МОм) контролировали во время экспериментов, и записи принимались для анализа, если какая-либо переменная не изменялась более чем на 20%. Регистрацию потенциала внеклеточного поля проводили с помощью стеклянных микроэлектродов (1–2 МОм), заполненных ACSF. Для стимуляции предполагаемых глутаматергических или ГАМКергических волокон использовались монополярные пипетки пластыря, заполненные ACSF (1 МОм), помещенные в BLA непосредственно медиальнее внешней капсулы. Долговременную депрессию тормозной передачи (iLTD) индуцировали путем подачи тормозной низкочастотной стимуляции (iLFS), состоящей из 100 стимулов с частотой 1 Гц, через стимулирующий электрод. Для высокочастотной стимуляции (ВЧС) в латеральную миндалину (ЛП) применяли четыре серии по 100 импульсов с частотой 100 Гц и межстимульным интервалом 20 с. Для регистрации потенциала внеклеточного поля интенсивность стимула регулировали для получения полумаксимальных ответов. Сигналы собирали с помощью усилителя Axopatch 200B (Molecular Devices). ЗАЖИМ 9программное обеспечение (Molecular Devices) использовалось для сбора данных. Сигналы фильтровались на частоте 1 кГц и оцифровывались на частоте 10 кГц.
Сигналы собирали с помощью усилителя Axopatch 200B (Molecular Devices). ЗАЖИМ 9программное обеспечение (Molecular Devices) использовалось для сбора данных. Сигналы фильтровались на частоте 1 кГц и оцифровывались на частоте 10 кГц.
Экстракция и анализ ЭХБ
Через неделю после последнего спровоцированного припадка ложных и разожженных крыс умерщвляли путем быстрого обезглавливания. И правую, и левую миндалины быстро вырезали и хранили при -80°C. Процесс экстракции липидов проводили, как описано ранее (Morena et al., 2015; Qi et al., 2015). Мозговую ткань взвешивали и помещали в культуральные пробирки из боросиликатного стекла, содержащие 2 мл ацетонитрила (ACN) с 5 пмоль [2H8]AEA и 5 нмоль [2H8]2-AG для экстракции, и гомогенизировали стеклянной палочкой. Ткань обрабатывали ультразвуком в течение 30 мин в ледяной воде и инкубировали в течение ночи при -20°C для осаждения белков, затем центрифугировали при 1500×9.1050 г для удаления твердых частиц. Супернатанты переносили в новую стеклянную пробирку и выпаривали досуха в атмосфере газа N 2 . Образцы восстанавливали в 300 мкл ACN и снова сушили в атмосфере газа N 2 . Экстракты липидов суспендировали в 20 мкл АЦН и хранили при –80°С до проведения анализа. Анализ AEA и 2-AG проводили с помощью жидкостной хроматографии в сочетании с масс-спектрометрией (ЖХ-МС/МС), как подробно описано ранее (Qi et al., 2015).
Образцы восстанавливали в 300 мкл ACN и снова сушили в атмосфере газа N 2 . Экстракты липидов суспендировали в 20 мкл АЦН и хранили при –80°С до проведения анализа. Анализ AEA и 2-AG проводили с помощью жидкостной хроматографии в сочетании с масс-спектрометрией (ЖХ-МС/МС), как подробно описано ранее (Qi et al., 2015).
Статистический анализ
Данные были проанализированы с помощью GraphPad Prism 6 (RRID: SCR_002798). Различия между группами анализировали с использованием критерия Стьюдента t для непарных данных, двустороннего или повторного измерения (RM) ANOVA с последующим апостериорным критерием Тьюки для множественных сравнений, когда это уместно. Значимость была принята при p <0,05. Статистический план для каждого эксперимента подробно описан в легенде к соответствующему рисунку.
Результаты
Повторяющиеся приступы миндалины вызывают стойкое межприступное усиление тревожноподобного поведения и нарушение сохранения памяти о страхе
Kindling моделирует патофизиологию височной эпилепсии и аберрантную нейронную пластичность, которая способствует межприступным эмоциональным и когнитивным сопутствующим заболеваниям (Teskey and Corcoran, 2009, 2015). Здесь, чтобы проверить, постоянно ли возбуждение миндалины изменяет эмоциональность, мы вызвали двадцать электрически вызванных электрографических припадков в течение двух недель, а затем мы оценили эмоциональное поведение у крыс, у которых не было приступов в течение одной недели, чтобы рассеять постиктальные эффекты. В этом исследовании все крысы, подвергшиеся киндлингу, показали по крайней мере три последовательных поведенческих припадка пятой стадии в конце двадцати стимуляций миндалевидного тела (данные не показаны). Мы оценивали тревожно-подобное поведение у ложных или кинженных крыс с помощью ЭРМ (рис. 19).1050 A ), в котором уменьшение времени, проведенного в распростертых объятиях EPM, интерпретируется как повышенная тревожность. Влияние киндлинга миндалины на тревожноподобное поведение ранее давало неоднозначные результаты, поскольку сообщалось как о анксиолитическом, так и анксиогенном эффекте киндлинговых припадков (Helfer et al., 1996; Adamec et al., 2004; Hannesson et al.
Здесь, чтобы проверить, постоянно ли возбуждение миндалины изменяет эмоциональность, мы вызвали двадцать электрически вызванных электрографических припадков в течение двух недель, а затем мы оценили эмоциональное поведение у крыс, у которых не было приступов в течение одной недели, чтобы рассеять постиктальные эффекты. В этом исследовании все крысы, подвергшиеся киндлингу, показали по крайней мере три последовательных поведенческих припадка пятой стадии в конце двадцати стимуляций миндалевидного тела (данные не показаны). Мы оценивали тревожно-подобное поведение у ложных или кинженных крыс с помощью ЭРМ (рис. 19).1050 A ), в котором уменьшение времени, проведенного в распростертых объятиях EPM, интерпретируется как повышенная тревожность. Влияние киндлинга миндалины на тревожноподобное поведение ранее давало неоднозначные результаты, поскольку сообщалось как о анксиолитическом, так и анксиогенном эффекте киндлинговых припадков (Helfer et al., 1996; Adamec et al., 2004; Hannesson et al. , 2008; Chen). и др., 2016). Здесь мы обнаружили, что крысы, подвергшиеся киндлингу миндалевидного тела, показали значительно сниженный процент времени, проведенного в открытых руках (рис. 1 B 9).1051) и записи с открытой рукой (рис. 1 C ) по отношению к фиктивной контрольной группе, что указывает на повышенное тревожное поведение. Эти эффекты не были связаны с изменением двигательной активности, поскольку киндлинг не влиял на общее расстояние, пройденное за 5-минутное задание (рис. 1 D ). Затем мы проверили динамику памяти о страхе на другой группе крыс. В отличие от тревожного поведения, интериктальные последствия воспламенения памяти о страхе подвергались гораздо меньшему количеству экспериментов. Ранее было показано, что возбуждение BLA ухудшает восстановление условных воспоминаний о страхе (Fournier et al., 2013; Botterill et al., 2014). Однако в этих исследованиях поведенческие оценки проводились через 1 день после последней киндлинговой стимуляции, когда постиктальное состояние еще могло влиять на поведенческие реакции.
, 2008; Chen). и др., 2016). Здесь мы обнаружили, что крысы, подвергшиеся киндлингу миндалевидного тела, показали значительно сниженный процент времени, проведенного в открытых руках (рис. 1 B 9).1051) и записи с открытой рукой (рис. 1 C ) по отношению к фиктивной контрольной группе, что указывает на повышенное тревожное поведение. Эти эффекты не были связаны с изменением двигательной активности, поскольку киндлинг не влиял на общее расстояние, пройденное за 5-минутное задание (рис. 1 D ). Затем мы проверили динамику памяти о страхе на другой группе крыс. В отличие от тревожного поведения, интериктальные последствия воспламенения памяти о страхе подвергались гораздо меньшему количеству экспериментов. Ранее было показано, что возбуждение BLA ухудшает восстановление условных воспоминаний о страхе (Fournier et al., 2013; Botterill et al., 2014). Однако в этих исследованиях поведенческие оценки проводились через 1 день после последней киндлинговой стимуляции, когда постиктальное состояние еще могло влиять на поведенческие реакции. Кроме того, авторы выполнили форму долговременной растопки, состоящую из 99 стимуляций. Здесь мы протестировали ложных и кинжированных крыс, используя классическую парадигму слухового условного рефлекса страха (рис. 1—) через неделю после последнего из двадцати вызванных приступов. В этой задаче животные учатся ассоциировать CS, тон, с US, электрическим током, и последующее повторное воздействие тона приводит к реакциям страха, которые включают замирание. Мы обнаружили, что киндлинг не изменяет приобретение реакции страха, поскольку вызванное КС-УЗ замирание во время кондиционирования выявило значительный эффект презентации КС-УЗ с течением времени в обеих группах, но между группами не было различий (рис. 1–9).1050 F ), что указывает на то, что все крысы приобрели вызванные CS реакции страха. Тем не менее, киндлинг значительно нарушил выученное выражение страха и восстановление памяти о страхе, поскольку у разожженных крыс наблюдалось снижение среднего замирания во время презентации CS при тестировании как через 24 часа (рис.
Кроме того, авторы выполнили форму долговременной растопки, состоящую из 99 стимуляций. Здесь мы протестировали ложных и кинжированных крыс, используя классическую парадигму слухового условного рефлекса страха (рис. 1—) через неделю после последнего из двадцати вызванных приступов. В этой задаче животные учатся ассоциировать CS, тон, с US, электрическим током, и последующее повторное воздействие тона приводит к реакциям страха, которые включают замирание. Мы обнаружили, что киндлинг не изменяет приобретение реакции страха, поскольку вызванное КС-УЗ замирание во время кондиционирования выявило значительный эффект презентации КС-УЗ с течением времени в обеих группах, но между группами не было различий (рис. 1–9).1050 F ), что указывает на то, что все крысы приобрели вызванные CS реакции страха. Тем не менее, киндлинг значительно нарушил выученное выражение страха и восстановление памяти о страхе, поскольку у разожженных крыс наблюдалось снижение среднего замирания во время презентации CS при тестировании как через 24 часа (рис. 1 G ), так и через 48 часов после кондиционирования (рис. 1 H ) по сравнению с с фиктивной группой.
1 G ), так и через 48 часов после кондиционирования (рис. 1 H ) по сравнению с с фиктивной группой.
Рис. 1.
Разожженные крысы проявляют тревожно-подобный фенотип с нарушением сохранения памяти о страхе. Схематическое изображение плана эксперимента по оценке влияния судорог на тревожноподобное поведение (9).0165 А ). Возбуждение миндалевидного тела увеличивало показатели тревожноподобного поведения в EPM ( B–D ). Влияние повторных припадков миндалины на процент времени, проведенного в открытых рукавах ( B , непарный тест t : t (19) = 2,28, p (50 6 записей) = 0,034), открытые руки C , непарный t тест: t (19) = 2,52, p = 0,021), и общее пройденное расстояние ( Д , непарный т тест: т (19) = 0,97, р = 0,34). Схематическое изображение плана эксперимента по оценке влияния судорог на динамику памяти о страхе ( E ). Повторные приступы миндалевидного тела уменьшали выражение страха через 24 и 48 ч после кондиционирования ( F – H ). Процентное замирание во время условного рефлекса страха ( F , двусторонний RM ANOVA, эффект судорог: Ф (1,25) = 1,32, р = 0,26; Эффект КС-УЗ: F (3,75) = 41,15, p < 0,0001; взаимодействие: F (3,75) = 0,36, p = 0,78), сохранение памяти о страхе в течение 24 ч ( G , двухфакторный RM ANOVA, эффект судорог: F 11 ,25) = 7,22, p = 0,013; влияние КС: F (6,150) = 21,43, p < 0,0001; взаимодействие: F (6,150) = 0,90, p = 0,50) и 48-часовое сохранение памяти о страхе ( H , двухфакторный RM ANOVA, эффект судорог: F = 7,496) , P = 0,011; CS эффект: F (2,50) = 42,35, P <0,0001; взаимодействие: F (2,50) = 3,06, P = 0,056) = 3,06, P = 0,056) .
Схематическое изображение плана эксперимента по оценке влияния судорог на динамику памяти о страхе ( E ). Повторные приступы миндалевидного тела уменьшали выражение страха через 24 и 48 ч после кондиционирования ( F – H ). Процентное замирание во время условного рефлекса страха ( F , двусторонний RM ANOVA, эффект судорог: Ф (1,25) = 1,32, р = 0,26; Эффект КС-УЗ: F (3,75) = 41,15, p < 0,0001; взаимодействие: F (3,75) = 0,36, p = 0,78), сохранение памяти о страхе в течение 24 ч ( G , двухфакторный RM ANOVA, эффект судорог: F 11 ,25) = 7,22, p = 0,013; влияние КС: F (6,150) = 21,43, p < 0,0001; взаимодействие: F (6,150) = 0,90, p = 0,50) и 48-часовое сохранение памяти о страхе ( H , двухфакторный RM ANOVA, эффект судорог: F = 7,496) , P = 0,011; CS эффект: F (2,50) = 42,35, P <0,0001; взаимодействие: F (2,50) = 3,06, P = 0,056) = 3,06, P = 0,056) . Данные выражены как среднее ± SEM * p < 0,05; ✦ p < 0,05 указывает на основной эффект судорог; n = 10–15 на группу.
Данные выражены как среднее ± SEM * p < 0,05; ✦ p < 0,05 указывает на основной эффект судорог; n = 10–15 на группу.
Повторяющиеся приступы миндалевидного тела не изменяют свойства мембран и внутреннюю возбудимость, но изменяют синаптическую активность пирамидных нейронов БЛА неделю после последнего спровоцированного припадка (рис. 2
A ). Сначала мы изучили внутренние свойства пирамидных нейронов BLA (рис. 29).1050 B ) и обнаружили, что все зарегистрированные пирамидные нейроны в обеих группах показали сходные профили мембран в ответ на серию шагов тока (рис. 2 B ), как показано на графиках ток-напряжение (I-V) (рис. 2 С ). Мембранные потенциалы покоя (рис. 2 D ), порог потенциала действия (рис. 2 E ), минимальный ток, необходимый для возбуждения потенциала действия (реобаза; рис. 2 F ) и число действий. потенциалов в ответ на токовые шаги (рис. 2 G ) были сходными между кинженными и ложными крысами. Таким образом, основные внутренние свойства нейронов BLA не были изменены повторной судорожной активностью, что согласуется с предыдущим сообщением (Shoji et al., 1998).
Таким образом, основные внутренние свойства нейронов BLA не были изменены повторной судорожной активностью, что согласуется с предыдущим сообщением (Shoji et al., 1998).Рисунок 2.
Повторяющиеся приступы миндалевидного тела изменяют синаптическую активность, не влияя на внутренние свойства пирамидных нейронов BLA. Схематическое изображение плана эксперимента по оценке влияния повторных припадков на электрофизиологические свойства пирамидных нейронов БЛА ( А ). Типичные мембранные ответы на этапы соматического тока ложных (вверху) и активированных (внизу) пирамидных нейронов BLA ( B ). Кривые ВАХ, показывающие отсутствие различий между фиктивными ( n = 10 клеток, 9 крыс) и активными ( n = 10 клеток, 10 крыс) группами (двусторонний RM ANOVA с током F (6,108) = 81,26, p < 0,0001 и судороги F (1,18) = 0,16, p = 0,69 как факторы, взаимодействие: Ф (6,108) = 0,67, р = 0,68; С ). Мембранный потенциал покоя (фиктивный: -71,7 ± 1,97, n = 10 клеток, 9 крыс vs киндированный: -69,40 ± 1,89 мВ, n = 10 клеток, 10 крыс; D ), потенциал действия порог (ложный: -38,90 ± 2,38 мВ, n = 10, 9 по сравнению с зажженным: -36,67 ± 1,24 мВ, n = 10, 10; E ) и реобаза (ложный: 284,09 ± A , д = 10, 9 против воспламененных: 186,2 ± 12,75 пА, n = 10, 10; F ) были одинаковыми в обеих группах; p > 0,05, тест Стьюдента t . G , Резюме кривых возбуждения тока, показывающих, что количество вызванных потенциалов действия в ответ на шаги соматического тока было одинаковым в пирамидных нейронах ложных ( n = 10, 9) и разожженных крыс ( n = 10, 10). Двусторонний RM ANOVA с током F (13,234) = 49,35, p < 0,0001 и изъятия F (1,18) = 0,17, p = 0,69 как факторы; взаимодействие: F (13,234) = 0,94, р = 0,51.
Мембранный потенциал покоя (фиктивный: -71,7 ± 1,97, n = 10 клеток, 9 крыс vs киндированный: -69,40 ± 1,89 мВ, n = 10 клеток, 10 крыс; D ), потенциал действия порог (ложный: -38,90 ± 2,38 мВ, n = 10, 9 по сравнению с зажженным: -36,67 ± 1,24 мВ, n = 10, 10; E ) и реобаза (ложный: 284,09 ± A , д = 10, 9 против воспламененных: 186,2 ± 12,75 пА, n = 10, 10; F ) были одинаковыми в обеих группах; p > 0,05, тест Стьюдента t . G , Резюме кривых возбуждения тока, показывающих, что количество вызванных потенциалов действия в ответ на шаги соматического тока было одинаковым в пирамидных нейронах ложных ( n = 10, 9) и разожженных крыс ( n = 10, 10). Двусторонний RM ANOVA с током F (13,234) = 49,35, p < 0,0001 и изъятия F (1,18) = 0,17, p = 0,69 как факторы; взаимодействие: F (13,234) = 0,94, р = 0,51. Representative traces and histograms ( H , I ) show that relative to sham rats, kindled rats display increased sEPSC frequency ( H , sham n = 12, 7, kindled n = 12, 6; непарные T Тест: T (22) = 5,35, P <0,0001) без существенного влияния на амплитуду SEPSC ( H , Sham N = 12, 7, Kindled N . , 6; непарный t тест: t (22) = 0,78, p = 0,44) и сниженная парная скважность ЭПСК ( I , имитация 19105 n = 10, 5, непарный t тест: t (18) = 2,38, р = 0,029). Representative traces and histograms ( J , K ) show that relative to sham rats, kindled rats display decreased sIPSC frequency ( J , sham n = 12, 5, kindled n = 12, 6; непарный t тест: t (22) = 3,91, p = 0,0008) без значимого влияния на амплитуду сТПСТ ( J , фиктивный н = 12, 5, зажженный н = 12, 6; Непарная T Тест: T (22) = 0,58, P = 0,57) и увеличение парного импульса IPSC ( K , SHAM N = 16, n , SHAM N = 16, .
Representative traces and histograms ( H , I ) show that relative to sham rats, kindled rats display increased sEPSC frequency ( H , sham n = 12, 7, kindled n = 12, 6; непарные T Тест: T (22) = 5,35, P <0,0001) без существенного влияния на амплитуду SEPSC ( H , Sham N = 12, 7, Kindled N . , 6; непарный t тест: t (22) = 0,78, p = 0,44) и сниженная парная скважность ЭПСК ( I , имитация 19105 n = 10, 5, непарный t тест: t (18) = 2,38, р = 0,029). Representative traces and histograms ( J , K ) show that relative to sham rats, kindled rats display decreased sIPSC frequency ( J , sham n = 12, 5, kindled n = 12, 6; непарный t тест: t (22) = 3,91, p = 0,0008) без значимого влияния на амплитуду сТПСТ ( J , фиктивный н = 12, 5, зажженный н = 12, 6; Непарная T Тест: T (22) = 0,58, P = 0,57) и увеличение парного импульса IPSC ( K , SHAM N = 16, n , SHAM N = 16, . = 12, 7; непарный т тест: т (26) = 2,32, р = 0,028). Данные выражены как среднее ± SEM; * p < 0,05 и *** p < 0,001.
= 12, 7; непарный т тест: т (26) = 2,32, р = 0,028). Данные выражены как среднее ± SEM; * p < 0,05 и *** p < 0,001.
Затем мы выдвинули гипотезу, что, несмотря на то, что внутренняя возбудимость пирамидных нейронов BLA не изменилась у разожженных крыс, sEPSC, опосредованные АМРА-рецептором (рис. 2 H ) на пирамидных нейронах BLA, будут подвержены повторной судорожной активности. Мы заметили, что частота sEPSCs значительно увеличилась в разожженной группе по сравнению с фиктивной группой (рис. 2 H ). Поскольку мы не обнаружили различий в амплитуде вВПСК между группами (рис. 2 H ), мы предположили, что индуцированное киндлингом изменение возбуждающей синаптической активности может быть связано с пресинаптической реакцией. Для проверки этой гипотезы мы измеряли вЭПСТ в ответ на парную стимуляцию с межстимульным интервалом 50 мс (рис. 2–9).1050 I ). Как сообщалось ранее (Acharjee et al. , 2018), парные стимуляции у ложных крыс вызывали ответы eEPSC с аналогичной амплитудой (Fig. 2 I ). Наоборот, у кинженных крыс было снижено соотношение парных импульсов, при этом второй ответ вЭПСТ был значительно меньше, чем первый (рис. 2, -1), и вместе с данными об увеличении частоты вПСТ, но не амплитуды, это согласуется с повышенным вероятность высвобождения нейротрансмиттера пресинаптическими глутаматергическими окончаниями на БЛА у кинженных крыс. Пирамидные нейроны также находятся под выраженным тоническим тормозным контролем со стороны ГАМКергических тормозных цепей, что побудило нас определить, как sIPSCs (рис. 2-9)1050 J ) подвержены повторной судорожной активности. Мы наблюдали снижение частоты, но не амплитуды sIPSCs (рис. 2 J ) в разожженной группе, что сопровождалось увеличением ГАМКергической парной частоты импульсов (рис. 2 K ), предполагая, что повторяющаяся судорожная активность вызывала снижение вероятности высвобождения ГАМК из ГАМКергических терминалей, нацеленных на главные нейроны BLA.
, 2018), парные стимуляции у ложных крыс вызывали ответы eEPSC с аналогичной амплитудой (Fig. 2 I ). Наоборот, у кинженных крыс было снижено соотношение парных импульсов, при этом второй ответ вЭПСТ был значительно меньше, чем первый (рис. 2, -1), и вместе с данными об увеличении частоты вПСТ, но не амплитуды, это согласуется с повышенным вероятность высвобождения нейротрансмиттера пресинаптическими глутаматергическими окончаниями на БЛА у кинженных крыс. Пирамидные нейроны также находятся под выраженным тоническим тормозным контролем со стороны ГАМКергических тормозных цепей, что побудило нас определить, как sIPSCs (рис. 2-9)1050 J ) подвержены повторной судорожной активности. Мы наблюдали снижение частоты, но не амплитуды sIPSCs (рис. 2 J ) в разожженной группе, что сопровождалось увеличением ГАМКергической парной частоты импульсов (рис. 2 K ), предполагая, что повторяющаяся судорожная активность вызывала снижение вероятности высвобождения ГАМК из ГАМКергических терминалей, нацеленных на главные нейроны BLA.
Повторяющиеся припадки миндалевидного тела снижают содержание AEA в BLA, а системное усиление передачи сигналов AEA устраняет эмоциональную дисфункцию, вызванную припадком
Хорошо известно, что передача сигналов eCB играет важную роль в модуляции эмоционального поведения благодаря жесткому контролю высвобождения пресинаптических нейротрансмиттеров в BLA (Morena et al., 2016a). Таким образом, мы спросили, изменяют ли повторные припадки стойкие уровни двух основных лигандов eCB в миндалевидном теле (рис. 3 —). Через неделю после последнего спровоцированного припадка уровни АЭА были значительно ниже в группе киндлинга по сравнению с группой имитации, в то время как содержание 2-AG не изменилось (рис. 3–9).1050 В , С ). Это подняло вопрос о том, были ли сниженные уровни AEA причиной изменения поведения. Таким образом, мы попытались восстановить повторяющееся вызванное судорогами изменение эмоционального поведения путем усиления передачи сигналов AEA посредством ингибирования его катаболического фермента амидгидролазы жирных кислот (FAAH; Kathuria et al. , 2003). В первом эксперименте мы лечили киндлинированных и ложных крыс системным введением либо ингибитора FAAH URB597 (0,5 мг/кг), либо его носителя за 1 ч до тестирования на ПКП (рис. 4–9).1050 А ). Мы снова наблюдали, что повторные припадки усиливали тревожноподобное поведение, так как группа разожженных, получавших носитель, проводила значительно меньше времени в открытых рукавах (рис. 4 B ) и выполняла значительно меньше входов в открытые рукава (рис. 4 C ). ) по сравнению с группой имитации/носителя. Аугментация AEA полностью избавила от этих изменений. Действительно, у разжиженных крыс, которым вводили URB597, были проценты времени, проведенного в открытых рукавах (рис. 4 B ) и входов в открытые рукава (рис. 49).1050 C ), эквивалентные таковым у ложных крыс, которым вводили либо носитель, либо URB597. Кроме того, ни судороги миндалины, ни медикаментозное лечение не влияли на двигательную активность крыс (рис. 4 D ). Затем мы спросили, может ли аугментация АЕА также спасти нарушение памяти о страхе у разожженных крыс (рис.
, 2003). В первом эксперименте мы лечили киндлинированных и ложных крыс системным введением либо ингибитора FAAH URB597 (0,5 мг/кг), либо его носителя за 1 ч до тестирования на ПКП (рис. 4–9).1050 А ). Мы снова наблюдали, что повторные припадки усиливали тревожноподобное поведение, так как группа разожженных, получавших носитель, проводила значительно меньше времени в открытых рукавах (рис. 4 B ) и выполняла значительно меньше входов в открытые рукава (рис. 4 C ). ) по сравнению с группой имитации/носителя. Аугментация AEA полностью избавила от этих изменений. Действительно, у разжиженных крыс, которым вводили URB597, были проценты времени, проведенного в открытых рукавах (рис. 4 B ) и входов в открытые рукава (рис. 49).1050 C ), эквивалентные таковым у ложных крыс, которым вводили либо носитель, либо URB597. Кроме того, ни судороги миндалины, ни медикаментозное лечение не влияли на двигательную активность крыс (рис. 4 D ). Затем мы спросили, может ли аугментация АЕА также спасти нарушение памяти о страхе у разожженных крыс (рис. 4 E ). Через неделю после последнего спровоцированного ложного или киндированного припадка крыс обеих групп, не получавших лекарств, обучали ассоциировать нейтральный стимул с ударом током по ступням во время сеанса кондиционирования (рис. 4–9).1050Е ). В конце сеанса кондиционирования мы обрабатывали фиктивных и кинженных крыс либо носителем, либо URB597 (0,5 мг/кг), а после лечения мы помещали крыс обратно в их домашнюю клетку. Мы решили вводить крысам сразу после кондиционирования, чтобы усилить передачу сигналов AEA во время фазы консолидации памяти о страхе, основываясь на предыдущих выводах, показывающих, что eCB, особенно AEA, модулируют консолидацию эмоциональной памяти (Morena et al., 2014, 2019). Как показано на Фигуре 4 F , во время кондиционирования не было ранее существовавших различий в замирании у крыс до лечения лекарственным средством. У разожженных крыс, получавших носитель, наблюдалось нарушение сохранения памяти о страхе как через 24 ч (рис.
4 E ). Через неделю после последнего спровоцированного ложного или киндированного припадка крыс обеих групп, не получавших лекарств, обучали ассоциировать нейтральный стимул с ударом током по ступням во время сеанса кондиционирования (рис. 4–9).1050Е ). В конце сеанса кондиционирования мы обрабатывали фиктивных и кинженных крыс либо носителем, либо URB597 (0,5 мг/кг), а после лечения мы помещали крыс обратно в их домашнюю клетку. Мы решили вводить крысам сразу после кондиционирования, чтобы усилить передачу сигналов AEA во время фазы консолидации памяти о страхе, основываясь на предыдущих выводах, показывающих, что eCB, особенно AEA, модулируют консолидацию эмоциональной памяти (Morena et al., 2014, 2019). Как показано на Фигуре 4 F , во время кондиционирования не было ранее существовавших различий в замирании у крыс до лечения лекарственным средством. У разожженных крыс, получавших носитель, наблюдалось нарушение сохранения памяти о страхе как через 24 ч (рис. 4–9,1050 G ) и через 48 часов (Фиг. 4 H ) после кондиционирования, поскольку мы наблюдали в этой группе снижение среднего замирания во всех проявлениях CS по сравнению с имитационной группой, получавшей носитель. URB597 восстанавливал изменения памяти о страхе в группе, получавшей возбуждение, поскольку мы больше не наблюдали различий в среднем замирании во время презентаций CS между группами имитация/носитель, имитация/URB и возбуждение/URB как через 24, так и через 48 часов после сеанса кондиционирования (рис. 4 ). Г , Н ).
4–9,1050 G ) и через 48 часов (Фиг. 4 H ) после кондиционирования, поскольку мы наблюдали в этой группе снижение среднего замирания во всех проявлениях CS по сравнению с имитационной группой, получавшей носитель. URB597 восстанавливал изменения памяти о страхе в группе, получавшей возбуждение, поскольку мы больше не наблюдали различий в среднем замирании во время презентаций CS между группами имитация/носитель, имитация/URB и возбуждение/URB как через 24, так и через 48 часов после сеанса кондиционирования (рис. 4 ). Г , Н ).
Рисунок 3.
Повторяющиеся припадки миндалины снижают содержание AEA в миндалине, оставляя уровни 2-AG неизменными. План эксперимента по определению влияния повторных припадков миндалины на уровни AEA и 2-AG в миндалине ( A ). Разожженные крысы ( n = 8) показали снижение уровня AEA (пмоль/г ткани) в миндалине по сравнению с ложными крысами ( n = 7; B , непарный тест t : t (13) = 2,27, р = 0,041). Различий в уровнях 2-АГ миндалины (нмоль/г ткани) между двумя экспериментальными группами не было ( С , непарные t тест: t (13) = 0,068, p 90,951 ). Данные выражены как среднее ± SEM; * р < 0,05.
Различий в уровнях 2-АГ миндалины (нмоль/г ткани) между двумя экспериментальными группами не было ( С , непарные t тест: t (13) = 0,068, p 90,951 ). Данные выражены как среднее ± SEM; * р < 0,05.
Рисунок 4.
Ингибитор FAAH URB597 нейтрализует пагубные эффекты повторяющихся припадков миндалины на тревожно-подобное поведение и 24- и 48-часовую задержку памяти о страхе. Схематическое изображение плана эксперимента для оценки URB597 влияние на повторяющееся тревожное поведение, вызванное припадками миндалины ( A ). Лечение URB597 обращало вспять повторяющееся, вызванное припадками миндалевидного тела усиление тревожноподобного поведения в EPM ( B-D ). Влияние возбуждения миндалины у крыс, получавших носитель или URB597, на процент времени, проведенного в открытых рукавах ( B , двусторонний ANOVA, эффект судорог: F (1,36) = 5,49, p = 0,025, эффект препарата: Ф (1,36) = 0,59, р = 0,45; взаимодействие: F (1,36) = 4,23, p = 0,047), входы в открытые рукава ( C , двусторонний дисперсионный анализ, эффект судорог: F () = 6,18, p = 0,018, эффект препарата: F (1,36) = 2,40, p = 0,13, взаимодействие: F (1,36) , = , = 1,36, 0,13), и общее пройденное расстояние ( D , двухфакторный дисперсионный анализ, эффект судорог: F (1,36) = 0,11, p = 0,74; лекарственное действие: F (1,36) = 1,64, p = 0,21; взаимодействие: F (1,36) = 0,38, p = 0,54). Схематическое изображение плана эксперимента для оценки влияния URB597 на повторяющиеся изменения динамики памяти о страхе, вызванные приступами миндалины ( E ). Лечение URB597 восстанавливало повторные нарушения выражения страха, вызванные припадками миндалевидного тела, через 24 и 48 часов после кондиционирования (9).0165 F–H ). Процентное замирание во время условного рефлекса страха ( F , двухсторонний RM ANOVA, эффект препарата: F (3,32) = 0,45, p = 0,72; эффект CS-US: F 1 (3,96) = 119,2, p < 0,0001; взаимодействие: F (9,96) = 0,49, p = 0,88), 24-часовая память о страхе (
Схематическое изображение плана эксперимента для оценки влияния URB597 на повторяющиеся изменения динамики памяти о страхе, вызванные приступами миндалины ( E ). Лечение URB597 восстанавливало повторные нарушения выражения страха, вызванные припадками миндалевидного тела, через 24 и 48 часов после кондиционирования (9).0165 F–H ). Процентное замирание во время условного рефлекса страха ( F , двухсторонний RM ANOVA, эффект препарата: F (3,32) = 0,45, p = 0,72; эффект CS-US: F 1 (3,96) = 119,2, p < 0,0001; взаимодействие: F (9,96) = 0,49, p = 0,88), 24-часовая память о страхе (  Данные выражены как среднее ± SEM. Критерий множественных сравнений Тьюки: * р < 0,05, ** р < 0,01; Критерий множественного сравнения Тьюки для эффекта препарата: ✦ p <0,05, ✦✦ p <0,01 воспламененный носитель по сравнению с ложным носителем; •• p < 0,01 зажженный носитель по сравнению с зажженным URB597; n = 8–12 на группу.
Данные выражены как среднее ± SEM. Критерий множественных сравнений Тьюки: * р < 0,05, ** р < 0,01; Критерий множественного сравнения Тьюки для эффекта препарата: ✦ p <0,05, ✦✦ p <0,01 воспламененный носитель по сравнению с ложным носителем; •• p < 0,01 зажженный носитель по сравнению с зажженным URB597; n = 8–12 на группу.
Системное усиление передачи сигналов АЕА восстанавливает повторяющиеся вызванные приступом изменения глутаматергической, но не ГАМКергической передачи, и восстанавливает баланс ИЭ в БЛА
Более низкие уровни AEA были связаны с дисфункцией тревожноподобного поведения, и усиление передачи сигналов AEA с помощью URB597 обращало эти изменения вспять. Затем мы попытались обратить вспять изменения синаптической физиологии, вызванные судорогами, используя тот же подход. Чтобы сохранить последовательный экспериментальный протокол с поведенческими тестами, мы выполнили in vivo введение либо URB597 (0,5 мг/кг), либо носителя, а затем, через 1 час после инъекции, приступили к умерщвлению и сбору срезов мозга для электрофизиологические опыты.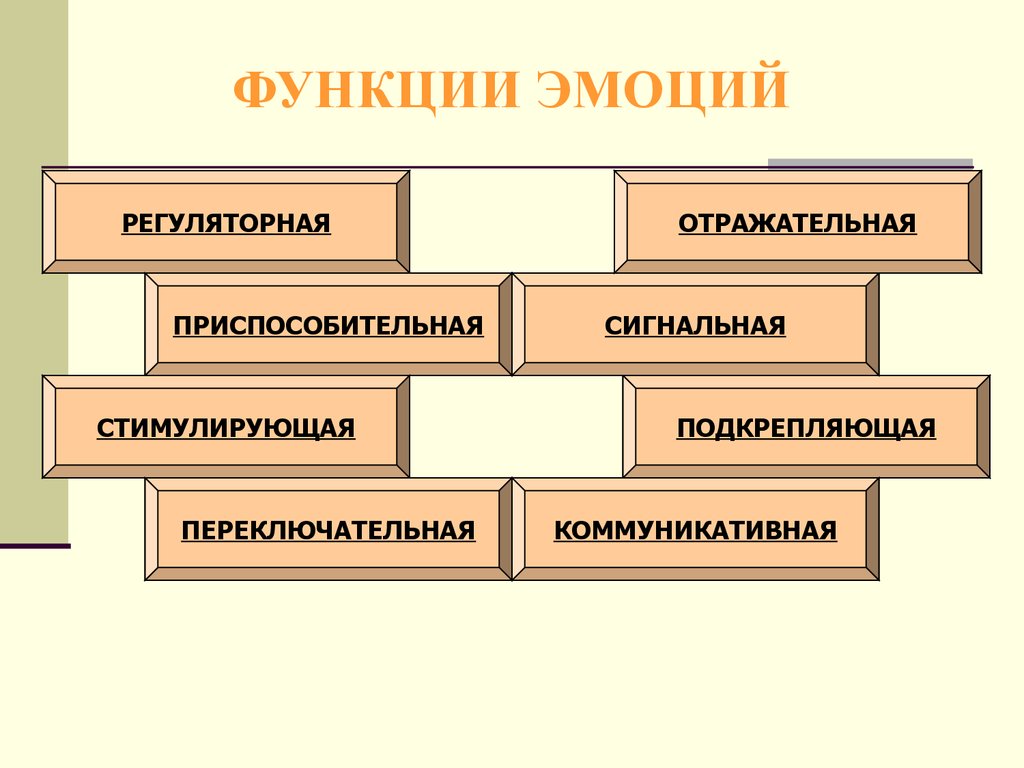 Поскольку как глутаматергическая, так и ГАМКергическая передача находятся под контролем системы eCB (Yoshida et al., 2011; Hill and Tasker, 2012), мы исследовали эффект системного введения либо URB59, либо7, или его носитель как на sEPSC, так и на sIPSC, зарегистрированных в пирамидных нейронах BLA в активированных и ложных группах. Не было обнаружено существенной разницы ни в средней частоте, ни в амплитуде sEPSCs пирамидных нейронов BLA, зарегистрированных у ложных крыс, предварительно обработанных URB597, по сравнению с ложными крысами, предварительно обработанными носителем (фиг. 5 A-C ). Тем не менее, мы обнаружили, что URB597 вызывал значительное снижение частоты сВПСТ у крыс, подвергшихся стимуляции, по сравнению с группой, подвергшейся стимуляции, которой вводили носитель (рис. 5–9).1050 B ), так что после введения URB597 не было никакой разницы между ложными и разожженными крысами. Таким образом, системное увеличение передачи сигналов AEA не изменяет высвобождение глутамата в BLA per se, но способно ослаблять вызванное судорогами повышенное высвобождение глутамата в основных нейронах BLA. Чтобы дополнительно подтвердить это открытие, мы обнаружили, что повышение уровня AEA восстанавливало соотношение парных импульсов eEPSC у разожженных крыс (рис. 5 D ). Действительно, мы не обнаружили различий в ответах eEPSC на парные стимулы при тестировании пирамидных нейронов BLA у киндлингированных крыс, которым вводили URB59.7 по отношению к группам, зарегистрированным в группе, имитирующей носитель, в то время как соотношение парных импульсов статистически отличалось между группой, подвергнутой возбуждению, обработанной носителем, и группой, подвергнутой возбуждению, обработанной URB597 (фиг. 5 E ). Поскольку рецептор CB1 также сильно экспрессируется на ГАМКергических окончаниях в BLA (Yoshida et al., 2011), мы исследовали влияние URB597 на передачу ГАМК BLA (Fig. 5 F ). Мы не обнаружили различий ни в частоте sIPSCs, ни в амплитуде sIPSCs, зарегистрированных в пирамидных нейронах ложной группы, обработанной URB597 или транспортное средство (рис. 5 G , H ). Более того, URB597 не влиял на индуцированное киндлингом снижение частоты sIPSC, поскольку мы не обнаружили различий между sIPSC клеток, зарегистрированных в группе, обработанной URB597 или носителем (фиг. 5 G ). Соотношение парных импульсов eIPSC (рис. 5 I ) также не отличалось в разожженной группе, получавшей либо URB597, либо носитель (рис. 5 J ). Эти данные демонстрируют, что увеличение AEA ослабляло индуцированное судорогами повышенное высвобождение пресинаптического глутамата, но не оказывало существенного влияния на индуцированное судорогами нарушение пресинаптического высвобождения ГАМК. Затем мы исследовали, является ли модулирующий эффект URB597 на глутаматергические, но не на ГАМКергические окончания, было достаточно для восстановления баланса IE, нарушенного повторными приступами миндалевидного тела. Как и ожидалось, пирамидальные нейроны в разожженной группе демонстрировали значительно сниженное соотношение IE-I по сравнению с фиктивной группой, которое восстанавливалось после увеличения AEA у разожженных крыс (рис. 5 K , L ).
Поскольку как глутаматергическая, так и ГАМКергическая передача находятся под контролем системы eCB (Yoshida et al., 2011; Hill and Tasker, 2012), мы исследовали эффект системного введения либо URB59, либо7, или его носитель как на sEPSC, так и на sIPSC, зарегистрированных в пирамидных нейронах BLA в активированных и ложных группах. Не было обнаружено существенной разницы ни в средней частоте, ни в амплитуде sEPSCs пирамидных нейронов BLA, зарегистрированных у ложных крыс, предварительно обработанных URB597, по сравнению с ложными крысами, предварительно обработанными носителем (фиг. 5 A-C ). Тем не менее, мы обнаружили, что URB597 вызывал значительное снижение частоты сВПСТ у крыс, подвергшихся стимуляции, по сравнению с группой, подвергшейся стимуляции, которой вводили носитель (рис. 5–9).1050 B ), так что после введения URB597 не было никакой разницы между ложными и разожженными крысами. Таким образом, системное увеличение передачи сигналов AEA не изменяет высвобождение глутамата в BLA per se, но способно ослаблять вызванное судорогами повышенное высвобождение глутамата в основных нейронах BLA. Чтобы дополнительно подтвердить это открытие, мы обнаружили, что повышение уровня AEA восстанавливало соотношение парных импульсов eEPSC у разожженных крыс (рис. 5 D ). Действительно, мы не обнаружили различий в ответах eEPSC на парные стимулы при тестировании пирамидных нейронов BLA у киндлингированных крыс, которым вводили URB59.7 по отношению к группам, зарегистрированным в группе, имитирующей носитель, в то время как соотношение парных импульсов статистически отличалось между группой, подвергнутой возбуждению, обработанной носителем, и группой, подвергнутой возбуждению, обработанной URB597 (фиг. 5 E ). Поскольку рецептор CB1 также сильно экспрессируется на ГАМКергических окончаниях в BLA (Yoshida et al., 2011), мы исследовали влияние URB597 на передачу ГАМК BLA (Fig. 5 F ). Мы не обнаружили различий ни в частоте sIPSCs, ни в амплитуде sIPSCs, зарегистрированных в пирамидных нейронах ложной группы, обработанной URB597 или транспортное средство (рис. 5 G , H ). Более того, URB597 не влиял на индуцированное киндлингом снижение частоты sIPSC, поскольку мы не обнаружили различий между sIPSC клеток, зарегистрированных в группе, обработанной URB597 или носителем (фиг. 5 G ). Соотношение парных импульсов eIPSC (рис. 5 I ) также не отличалось в разожженной группе, получавшей либо URB597, либо носитель (рис. 5 J ). Эти данные демонстрируют, что увеличение AEA ослабляло индуцированное судорогами повышенное высвобождение пресинаптического глутамата, но не оказывало существенного влияния на индуцированное судорогами нарушение пресинаптического высвобождения ГАМК. Затем мы исследовали, является ли модулирующий эффект URB597 на глутаматергические, но не на ГАМКергические окончания, было достаточно для восстановления баланса IE, нарушенного повторными приступами миндалевидного тела. Как и ожидалось, пирамидальные нейроны в разожженной группе демонстрировали значительно сниженное соотношение IE-I по сравнению с фиктивной группой, которое восстанавливалось после увеличения AEA у разожженных крыс (рис. 5 K , L ).
Рисунок 5.
Ингибитор FAAH URB597 восстанавливает вызванное судорогами изменение глутаматергической, но не ГАМКергической передачи, и восстанавливает соотношение IE-E в BLA. Репрезентативные следы ( A ) и гистограммы ( B , C ) показывают влияние URB597 на sEPSCs у ложных и активированных крыс. Лечение URB597 восстанавливало повторяющееся индуцированное судорогами миндалевидного тела увеличение частоты sEPSC ( B , имитация носителя, n = 12, 9, активированный носитель, n = 10, 9, имитация URB597, = , 9 7, зажженный URB597, n = 13, 8, двухфакторный дисперсионный анализ, эффект препарата: F (1,41) = 14,18, р = 0,0005; судорожный эффект: F (1,41) = 27,39, p < 0,0001; взаимодействие: F (1,41) = 11,96, p = 0,0013) без значимого влияния на амплитуду сВПСТ ( C , ложный носитель, n = носитель 12, 9, 1 кинлед = 10, 9, фиктивный URB597, n = 10, 7, воспламененный URB597, n = 13, 8, двухфакторный дисперсионный анализ, эффект препарата: F (1,41) = 1,83, р = 0,18; судорожный эффект: F (1,41) = 0,048, р = 0,83; взаимодействие: F (1,41) = 0,057, р = 0,81). Кроме того, лечение URB597 восстанавливало повторное индуцированное приступом миндалевидного тела снижение частоты парных импульсов EPSC ( D , E , фиктивный носитель, n = 10, 8, зажженный носитель, n 9105, 6, фиктивный URB597, n = 13, 8, зажженный URB597, н = 13, 7; двухсторонний ANOVA, эффект препарата: F (1,42) = 5,81, p = 0,020; судорожный эффект: F (1,42) = 8,40, p = 0,0059; взаимодействие: F (1,42) = 2,79, p = 0,10). Репрезентативные кривые ( F ) и гистограммы ( G , H ) показывают влияние лечения URB597 на sIPSCs у ложных и разожженных крыс. УРБ597 лечение не влияло на повторное снижение частоты sIPSC, вызванное судорожным припадком миндалины ( G , имитация носителя, n = 12, 9, активированный носитель, n = 12, 9, имитация URB597, n = 12, 8, зажженный URB597, n = 13, 8; двухсторонний ANOVA, эффект препарата: F (1,45) = 1,81, p = 0,19; эффект судорог: F (51
51
51 1,45)
= 20,09, p < 0,0001 взаимодействие: F (1,45) = 3,03, p = 0,089) и не влияли на амплитуду сТПСТ ( H , фиктивный наполнитель, n = 12, 9, зажженный наполнитель, n = 12,9 9, фиктивный URB597, n = 12, 8, активированный URB597, n = 13, 8, двухфакторный дисперсионный анализ, эффект препарата: F (1,45) = 0,55, p = 0,55 эффект судорог: F (1,45) = 0,20, p = 0,65 взаимодействие: F (1,45) = 0,87, р = 0,36). Лечение URB597 не влияло на повторное индуцированное припадком миндалевидного тела повышение частоты парных импульсов IPSC ( I , J , фиктивный носитель, n = 21,13, активированный носитель, n 9105, 7, фиктивный URB597, n = 17, 9, активированный URB597, n = 23,15, двухфакторный дисперсионный анализ, эффект препарата: F (1,68) = 1,73, p = судорожный эффект: F (1,68) = 21,59, р < 0,0001; взаимодействие: F (1,68) = 0,40, p = 0,53). Репрезентативные кривые ( K ) показывают влияние повторяющихся припадков миндалины и ингибирования FAAH на отношение I-V для чисто возбуждающих (заблокированных 20 мкМ DNQX, ниже) и чистых тормозных (заблокированных 20 мкМ бикукуллина, вверху) вызванные синаптические токи. Ответы регистрировали при удерживающих потенциалах -60 и 0 мВ соответственно. URB597 восстанавливал индуцированное повторяющимся припадком миндалевидного тела снижение соотношения IE-E у разожженных крыс (9).0165 L , фиктивный ТС, н = 11, 6, пламенный ТС, н = 11, 6, фиктивный УРБ597, н = 9, 5, факельный УРБ597, = 7, 0501 н; двухсторонний ANOVA, эффект препарата: F (1,33) = 0,92, p = 0,35; судорожный эффект: F (1,33) = 1,34, p = 0,26; взаимодействие: F (1,33) = 10,70, p = 0,0025). Данные выражены как среднее ± SEM. Тест множественных сравнений Тьюки: * р < 0,05, ** р < 0,01, *** р < 0,001.
Повторяющиеся приступы миндалевидного тела изменяют как тоническую, так и фазическую передачу сигналов eCB
Тоническая передача сигналов eCB была зарегистрирована в различных областях мозга (Oliet et al., 2007; Lee et al., 2015), а также в BLA (Zhu and Lovinger, 2005). Поскольку мы обнаружили стойкое изменение содержания AEA в миндалевидном теле, мы проверили, модулирует ли тоническая передача сигналов eCB высвобождение пресинаптических нейротрансмиттеров в BLA и влияет ли подавление AEA на тоническую активность eCB. Тоническую передачу сигналов eCB можно наблюдать как увеличение базальной синаптической активности после фармакологической блокады рецепторов CB1. Здесь мы регистрировали либо eEPSC, либо eIPSC до и после применения в ванне антагониста рецептора CB1 AM251 (2 мкм) у ложных и разожженных крыс. Мы обнаружили, что AM251 вызывает значительное увеличение амплитуды eEPSC (рис. 6-9).1050 A–C ), не влияя на амплитуду eIPSC (рис. 6 D–F ) пирамидных нейронов, зарегистрированных в фиктивной группе, что указывает на то, что передача сигналов eCBs тонически контролирует базальную передачу глутамата, но не передачу ГАМК в BLA. Интересно, что применение AM251 в ванне не приводило к значительному увеличению амплитуды eEPSC в разожженной группе (рис. 6 B ), что указывает на то, что тонический контроль eCB за высвобождением глутамата в BLA изменяется повторными приступами миндалевидного тела.
Рисунок 6.
Повторяющиеся приступы миндалевидного тела изменяют как тоническую, так и фазовую передачу сигналов eCB. Графики ( A , B ) и кривые образцов ( C ) eEPSC показывают, что применение в ванне антагониста рецептора CB1 AM251 увеличивало амплитуду eEPSC 1006 крыс, но не генерировало 1006 B , фиктивный, n = 8, 6, воспламененный, n = 9, 7, двусторонний RM ANOVA, эффект захватов: F (1,15) = 5,48, р = 0,034; Эффект АМ251: F (1,15) = 17,04, р = 0,0009; взаимодействие: F (1,15) = 5,48, p = 0,034). Графики ( D , E ) и кривые выборки ( F ) eIPSC, показывающие, что применение в ванне антагониста рецептора CB1 AM251 не влияло на амплитуду eIPSC ни у крыс, симулированных, ни у крыс E , фиктивный, н = 7, 7, раскаленный, н = 7, 7; двухсторонний RM ANOVA, эффект судорог: F (1,12) = 0,73, p = 0,41; Эффект АМ251: F (1,12) = 0,063, p = 0,81; взаимодействие: F (1,12) = 0,73, p = 0,41). Графики ( G , H ) и прослеживаемые образцы ( I ) eIPSC показывают, что индукция LTD была блокирована у крыс, подвергшихся возбуждению с помощью носителя ( n = 8, 8), по сравнению с ложными крысами-носителями ( n = 9, 9) и восстанавливался у киндлинированных крыс, получавших URB597 ( H , ложный URB597, n = 7, 6, зажженный URB597, n = 8, 8; двухфакторный RM ANOVA, эффект препарата: F (3,28) = 4,66, p = 0,009; эффект LFS: F (1,28) ) = 85,45, p < 0,0001 Взаимодействие: F (3,28) = 4,66, p = 0,009). Данные в B , E показывают среднее значение первых 5 минут записи до применения ванны AM251 (до AM) и от 21 до 25 минут записи (после AM). Данные в H показывают среднее значение первых 10 минут записи до LFS (до LFS) и от 31 до 35 минут записи (после LFS). Данные выражены как среднее ± SEM. Критерий множественного сравнения Тьюки: ** p < 0,01, *** p < 0,001 имитация (носитель) по сравнению с воспламененным (носитель), • p < 0,05 зажженный автомобиль по сравнению с зажженным URB597.
Фазовый режим мобилизации eCB, зависящий от активности, представляет собой основной механизм, с помощью которого eCB опосредуют кратковременные и долговременные формы пластичности как в возбуждающих, так и в тормозных синапсах. В то время как вызванное деполяризацией кратковременное подавление высвобождения нейротрансмиттера осуществляется исключительно действием 2-AG на пресинаптические рецепторы CB1 во многих областях мозга (Kano et al., 2009), включая BLA (Yoshida et al., 2011), iLTD в BLA, вероятно, опосредуется AEA (Marsicano et al., 2002; Azad et al., 2004). В соответствии с предыдущими отчетами (Marsicano et al., 2002) мы обнаружили, что низкочастотной стимуляции (iLFS; 100 импульсов с частотой 1 Гц) внешней капсулы было достаточно, чтобы вызвать LTD eIPSCs (iLTD), зарегистрированных в пирамидных нейронах BLA от ложный носитель крыс (рис. 6 G–I ). Напротив, iLFS не вызывал какой-либо значимой iLTD в разожженной группе при инъекции носителя, в то время как предварительная обработка URB597 была способна восстановить индукцию iLTD у разожженных крыс (рис. 6 G-I ). Вместе эти данные свидетельствуют о том, что в BLA как тоническая, так и фазическая передача сигналов eCB нарушаются после повторяющейся судорожной активности.
Усиление передачи сигналов AEA восстанавливает опосредованную eCB долгосрочную синаптическую пластичность в BLA после повторяющихся припадков миндалины
Синаптическая пластичность в миндалевидном теле имеет решающее значение для консолидации памяти о страхе (Nabavi et al., 2014), и считается, что ЛП является местом хранения ассоциаций CS-US с базальными ядрами (BA), которые представляют собой важный ретранслятор потенцированных активность ЛА. Было показано, что путь LA-BA является важным участком пластичности (Rammes et al., 2000; Azad et al., 2004), особенно в модуляции консолидации памяти о страхе (Anglada-Figueroa and Quirk, 2005; Amano et al. , 2011). Более того, было показано, что передача сигналов eCB влияет на пластичность этого пути (Marsicano et al., 2002; Azad et al., 2004). Здесь мы проверили, влияют ли повторяющиеся приступы миндалины на способность синапсов BLA подвергаться долговременной потенциации (LTP) пути LA-BA. Мы наблюдали, что ВЧС (четыре серии по 100 импульсов частотой 100 Гц) в ЛП вызывала небольшую, но достоверную ДВП амплитуды поля ВПСП (пВПСП) в БА (рис. 7-9).1050 А–С ). Тем не менее, ДП может быть вызвана и у разжиженных крыс, что свидетельствует о том, что повторяющаяся судорожная активность не влияет на индукцию и поддержание ДП (рис. 7, , А-С, ). Сообщалось, что LTP может модулироваться с помощью eCB-опосредованной iLTD во многих областях мозга (Chevaleyre et al., 2006), включая BLA (Azad et al., 2004). Поскольку мы действительно обнаружили, что повторяющаяся судорожная активность в миндалевидном теле ухудшает iLTD (рис. 6 G-I ), мы попытались индуцировать LTP в BLA, когда ему предшествовала iLFS-индуцирующая iLTD (100 импульсов с частотой 1 Гц). Azad и его коллеги показали, что в BLA величина LTP значительно увеличивается, если HFS предшествует iLFS (Azad et al., 2004). Соответственно, мы наблюдали, что когда iLFS индуцировали за 10 минут до HFS, получающаяся в результате LTP была значительно повышена у ложных крыс/крыс-носителей (рис. 7–9).1050 D – F ). Тем не менее, iLTD-индуцированное усиление LTP полностью отсутствовало в разжиженной группе, получавшей носитель (рис. 7 G-I ). Поскольку iLTD может быть восстановлен путем предварительной обработки URB597, как показано выше (рис. 6 G-I ), мы проверили, может ли увеличение AEA восстановить надлежащее iLTD-опосредованное усиление LTP. Действительно, мы обнаружили, что iLTD-опосредованная усиленная LTP может быть индуцирована в разожженной группе, получавшей URB597 (рис. 7, , J-L, ), что указывает на то, что повторная судорожная активность в миндалевидном теле влияет на фазическую передачу сигналов AEA для передачи ГАМК в BLA. что, следовательно, изменяет величину LTP в синапсах LA-BA.
Рисунок 7.
Ингибитор FAAH восстанавливает долговременную синаптическую пластичность в BLA после повторных приступов миндалевидного тела. Графики ( A , B ) и выборочные кривые ( C ) fEPSP, записанные в BLA, показывающие, что HFS индуцирует LTP в обоих плацебо-носителях (710050 n n n n n ) , и зажженный носитель ( n = 5, 5) крысы ( B ; двусторонний RM ANOVA, судорожный эффект F (1,11) = 0,75, р = 0,40; Эффект HFS F (1,11) = 26,88, p = 0,0003; взаимодействие F (1,11) = 0,78, р = 0,40; Множественный сравнительный тест Тьюки: фиктивный носитель до ВЧС по сравнению с пост-ВЧС, p <0,05; зажженный носитель до ВФС по сравнению с пост-ВФС, p <0,01). Графики ( D , E , G , H ) и образцы трасс ( F , I ) показывают, что при применении iLFS за 10 мин до HFS результирующая LTP значительно увеличивается в ложном растворителе ( E , n два 9,051; -7 путь RM ANOVA, эффект iLFS: F (1,13) = 6,63, p = 0,023, эффект HFS: F (1,13) = 24,64, p 90,0051; взаимодействие = 3; F (1,13) = 6,70, p = 0,023), но не у кинледных крыс ( Н , н = 6, 6; двухсторонний RM ANOVA, эффект iLFS: F (1,9) = 2,68, p = 0,14; Эффект ВЧС: F (1,9) = 43,34, p = 0,0001; взаимодействие: F (1,9) = 2,54, p = 0,15). Графики ( J , K ) и кривые образцов ( L ) показывают, что обработка URB597 значительно увеличивала LTP у разожженных крыс, когда iLFS применяли за 10 минут до HFS (9). 0165 К , фиктивный УРБ597, н = 7, 4, зажженный УРБ597 н = 8, 5; двухсторонний RM ANOVA, эффект препарата: F (3,24) = 3,38, p = 0,035; эффект iLFS+HFS: F (1,24) = 76,08, p < 0,0001; взаимодействие: F (3,24) = 3,32, p = 0,037). Данные в B , E , H , K показывают среднее значение первой 10-минутной записи исходного уровня до HFS или iLFS (до) и последней 10-минутной записи после HFS (после). Данные ложного транспортного средства и зажженного транспортного средства повторяются на графиках, поскольку они относятся к одному и тому же экспериментальному плану. Данные выражены как среднее ± SEM. Критерий множественных сравнений Тьюки: * p < 0,05; Множественный сравнительный тест Тьюки на эффект препарата: ✦✦✦ p <0,001 воспламененный носитель по сравнению с ложным носителем; • p < 0,05 зажженный автомобиль по сравнению с зажженным URB597.
Дискуссия
Здесь мы впервые показали, что повторяющаяся судорожная активность в миндалевидном теле вызывает стойкое и сохраняющееся усиление тревожноподобного поведения и снижение ретенции памяти о страхе, что связано с устойчивым подавлением АЭА в миндалевидное тело. Это снижение передачи сигналов AEA привело к потере тонического торможения глутаматергических синапсов и потере фазового контроля над ГАМКергическими синапсами, что привело к эмоциональным изменениям. Более того, ингибирование FAAH нормализовало как поведенческую, так и электрофизиологическую дисфункцию у крыс с повторными приступами.
Базальные уровни AEA жестко управляют пресинаптической активностью нейронов BLA и, следовательно, эмоциональным поведением (Morena et al., 2019). В то время как уровни AEA вместе с 2-AG кратковременно повышаются во время острого припадка в области мозга, участвующей в судорожной активности (Marsicano et al., 2003; Wallace et al., 2003), хроническое влияние повторной судорожной активности на содержание eCB получил ограниченное внимание в животных моделях эпилепсии. Чен и его коллеги (Chen et al., 2003) обнаружили, что в модели фебрильных судорог через неделю после судорог не происходит изменений в уровнях eCB в гиппокампе. И наоборот, мы обнаружили, что уровни AEA, но не 2-AG, постоянно снижаются в миндалевидном теле через неделю после последнего приступа. В соответствии с нашими выводами аналогичные результаты были получены в клиническом исследовании, в котором содержание AEA было значительно ниже в спинномозговой жидкости людей с височной эпилепсией (Romigi et al., 2010). Наши данные свидетельствуют о том, что хотя передача сигналов АЕА после припадка задействуется для подавления чрезмерной активности нейронов и восстановления гомеостатического состояния, предшествующего приступу (Marsicano et al., 2003), повторяющаяся судорожная активность вызывает стойкое аллостатическое подавление передачи сигналов АЕА в миндалевидном теле. , которые могут вызывать пресинаптические изменения пирамидных нейронов BLA и впоследствии интериктальную эмоциональную дисфункцию. Действительно, в соответствии с предыдущими выводами мы обнаружили, что повторяющиеся изменения основных нейронов BLA, вызванные судорогами, ограничиваются синаптическим уровнем, в то время как внутренние свойства нейронов не затрагиваются повторяющейся судорожной активностью (Shoji et al., 19).98; Шуберт и др., 2005). В дополнение к этому, здесь мы показали, что стойкие синаптические изменения, вызванные повторяющимися припадками, ограничиваются пресинаптическими глутаматергическими и ГАМКергическими окончаниями, вызывая несбалансированную передачу IE на пирамидные нейроны BLA. Важно отметить, что сообщалось, что пресинаптические изменения передачи глутамата миндалевидного тела с последующей аномальной гиперактивностью BLA связаны с тревожноподобными фенотипами (Aroniadou-Anderjaska et al., 2012; Lowery-Gionta et al., 2018) и низким AEA. содержание в миндалевидном теле связано с повышенной активностью BLA, что приводит к тревожному поведению (Gray et al., 2015). Тот факт, что наши данные показали, что системное введение ингибитора FAAH URB597 ослаблял как глутаматергическую пресинаптическую гиперактивность, так и тревожное поведение у разожженных крыс, убедительно указывает на то, что дефицит передачи сигналов AEA в BLA способствует гипервозбудимости BLA и развитию межприступной тревоги. Это открытие представляет собой новый потенциальный механизм, вызывающий коморбидную тревогу при расстройствах, связанных с судорогами.
Передача сигналов eCB осуществляет пресинаптический тонический контроль высвобождения как глутамата, так и ГАМК, формируя синаптическую силу нейронных сетей (Zhu and Lovinger, 2005; Oliet et al., 2007). Таким образом, снижение содержания AEA в BLA после киндлинга, вероятно, ограничивает тоническое действие eCB на высвобождение глутаматергического нейротрансмиттера, вызывая гиперактивность BLA. Хотя сообщалось, что тоническая передача сигналов eCB происходит в ГАМКергических синапсах в гиппокампе (Lee et al., 2015), также было продемонстрировано тоническое действие eCB на возбуждающую передачу в BLA (Yasmin et al., 2020). В соответствии с этим последним исследованием мы наблюдали, что у фиктивных крыс блокада рецептора CB1 выявила тоническое действие eCB на глутамат, но не на нейротрансмиссию ГАМК в BLA. Более того, мы обнаружили, что этот тонический контроль ЭХБ при возбуждающей передаче нарушен у кинженных крыс. Эти данные вместе с выводами о том, что ингибитор FAAH ослабляет индуцированную судорогами гиперактивность глутамата, не влияя на передачу ГАМК, дают убедительные доказательства того, что аллостатическая дезадаптация передачи сигналов AEA в миндалевидном теле ухудшает опосредованный eCB тонический контроль над высвобождением глутамата, что, в свою очередь, вызывает усиленное возбуждение BLA и, в конечном итоге, тревожноподобный фенотип. Важно отметить, что вместе с AEA 2-AG служит тонической передаче сигналов eCB (Lee et al., 2015). Однако недавнее исследование показало, что ингибирование синтеза 2-AG в BLA не вызывает изменений в базальной передаче глутамата и ГАМК, что исключает возможную роль 2-AG в опосредованном eCB тоническом контроле высвобождения нейротрансмиттера на Пирамидные нейроны BLA (Yasmin et al., 2020). С другой стороны, конститутивная активность рецептора CB1 в отсутствие эндогенного лиганда, который тонически регулирует высвобождение нейротрансмиттера, также не может быть исключена (Lee et al. , 2015). Тем не менее, наши данные свидетельствуют о решающей роли тонической потери сигналов AEA после индуцированных судорогами изменений в физиологии и эмоциональном поведении BLA, которые параллельны исследованиям, показывающим, как воздействие стресса приводит к снижению передачи сигналов AEA миндалевидного тела, вызывая усиление пресинаптической глутаматергической передачи BLA. , что приводит к тревожному поведению (Gray et al., 2015; Yasmin et al., 2020).
Передача сигналов AEA миндалевидного тела также играет решающую роль в консолидации памяти о страхе (Morena et al., 2014), а нарушения памяти о страхе связаны с изменением передачи сигналов AEA (Morena et al., 2019). LTP и LTD в миндалевидном теле широко считаются синаптической основой, лежащей в основе ассоциативной памяти, включая эмоциональную память (Nabavi et al., 2014), и изменения синаптической пластичности в миндалевидном теле сильно влияют на динамику памяти о страхе (Marsicano et al., 2002; Nabavi). и др., 2014). Широко продемонстрирована центральная роль eCBs в модуляции долговременной синаптической пластичности (Chevaleyre et al., 2006). В частности, в гиппокампе и миндалевидном теле передача сигналов AEA участвует как в LTP (Zimmermann et al., 2019) и LTD (Азад и др., 2004). Судорожная активность изменяет синаптическую пластичность в гиппокампе (Colangeli et al., 2017) и в миндалевидном теле (Schubert et al., 2005), а недавнее исследование показало, что усиление AEA способно восстанавливать как краткосрочные, так и долгосрочные синаптическая пластичность в зубчатой извилине после киндлинга перфорантного пути (Colangeli et al., 2017). Следовательно, возможно, что межприступная дисфункция памяти о страхе может быть вызвана изменениями AEA-зависимой долговременной пластичности в BLA. LTD передачи ГАМК (iLTD) представляет собой форму синаптической пластичности, заключающуюся в длительном снижении высвобождения медиатора из ГАМКергических терминалей (Chevaleyre et al., 2006). В BLA iLTD опосредуется передачей сигналов AEA и играет важную роль в настройке величины LTP, индуцированной HFS (Azad et al. , 2004). Здесь мы обнаружили, что, несмотря на то, что у кинженных крыс наблюдается нарушение формирования памяти о страхе, индукция LTP в миндалевидном теле остается неизменной после повторной судорожной активности. И наоборот, мы обнаружили, что как индукция iLTD, так и модуляция LTP, опосредованная iLTD, значительно ухудшаются после повторных приступов миндалины, что указывает на то, что повторная активность приступов изменяет синаптическую пластичность, особенно в синапсах ГАМК. Было показано, что структурная и функциональная пластичность ГАМКергических синапсов в БА возникает во время кондиционирования страха (Kasugai et al., 2019).), таким образом, можно предположить, что изменения пластичности ГАМК, вызванные судорогами, вероятно, объясняют изменение сохранения памяти о страхе у разожженных крыс. Однако предыдущие исследования показывают, что стойкое снижение базальной ГАМКергической активности вместе с устойчивым увеличением возбудительного влечения в BLA обычно связано с усилением выражения страха (Ehrlich et al. , 2009). Возможное объяснение этого несоответствия заключается в том, что синаптическая пластичность, возникающая во время ассоциативного обучения, требует определенного во времени нарушения баланса возбуждения и торможения (Letzkus et al., 2015), таким образом, стойкое и неспецифическое растормаживание, вызванное повторяющимися приступами, вероятно, влияет на височную специфичность локальной тормозной сети и точная активация глутаматергической активности для управления поведенческими реакциями. В поддержку этой интерпретации мы обнаружили, что ингибирование FAAH нормализовало баланс IE в BLA и восстанавливало зависимую от активности eCB-индуцированную депрессию локальной тормозной сети у разожженных крыс. Таким образом, это предполагает, что физиологическая роль передачи сигналов AEA в BLA заключается в поддержании гомеостатического тонического контроля над глутаматергической передачей при одновременном осуществлении точного фазового контроля над передачей сигналов GABA, что обеспечивает индукцию пластичности. Таким образом, наши данные свидетельствуют о том, что судороги постоянно изменяют тоническую передачу сигналов AEA на глутаматергических окончаниях, вызывая гипервозбудимость BLA, что может способствовать повышению тревожно-подобного поведения. Более того, повторяющиеся приступы миндалевидного тела нарушают зависящую от активности передачу сигналов AEA при ГАМКергической передаче в BLA, что может объяснять нарушение сохранения памяти о страхе. В поддержку этой гипотезы усиление передачи сигналов AEA за счет ингибирования FAAH восстанавливало межприступные изменения эмоционального поведения. В то время как доступная литература поддерживает гипотезу о том, что эти эффекты, вероятно, опосредованы, по крайней мере, частично, через усиление передачи сигналов рецептора AEA/CB1 в собственно BLA, важно отметить, что в этом исследовании ингибитор FAAH был назначен системно, поэтому нельзя исключать влияние увеличения AEA в других областях мозга, важных для памяти о страхе и тревоге. Действительно, было показано, что модулирование передачи сигналов AEA в гиппокампе или префронтальной коре может влиять на консолидацию памяти о страхе и поведение, подобное тревоге (Rubino et al. , 2008; Campos et al., 2010; Morena et al., 2014; Lisboa et al. и др., 2015 г. Хартманн и др., 2019 г.; Циммерманн и др., 2019).
В заключение, наши данные показывают, что аллостатическая дезадаптация передачи сигналов AEA играет решающую роль в развитии интериктальной эмоциональной дисфункции, указывая на передачу сигналов AEA и фермент FAAH в качестве главной мишени для развертывания новых терапевтических вариантов коморбидных эмоциональных изменений, происходящих в лица с эпилепсией. Действительно, клинические исследования показали, что генетические полиморфизмы, которые снижают активность FAAH и повышают базальные уровни AEA, связаны со снижением тревожности, облегчением консолидации эмоциональных воспоминаний и защитой от негативных эмоциональных последствий после воздействия стресса (Spagnolo et al., 2016; Mayo et al. ., 2020b), и аналогичные результаты были получены у людей, получавших ингибиторы FAAH (Mayo et al., 2020a). Важно отметить, что ингибиторы FAAH, подходящие для использования человеком, безопасные, хорошо переносимые и не вызывающие злоупотребления, уже разработаны (Li et al. , 2012; Mayo et al., 2020a) и потенциально могут использоваться в клинических испытаниях. для лечения интериктальных эмоциональных сопутствующих заболеваний. Более того, учитывая потенциальный противосудорожный эффект ингибиторов FAAH (Colangeli et al., 2017), эти данные теперь указывают на то, что ингибиторы FAAH могут иметь потенциальный двухсторонний терапевтический подход к расстройствам, связанным с судорогами, будучи способными как снижать тяжесть судорог, себя, а также облегчить развитие сопутствующих эмоциональных изменений. Этот двойной подход к лечению предлагает совершенно новый подход к терапии расстройств, связанных с судорогами.
Сноски
Эта работа была поддержана грантами Канадского института исследований в области здравоохранения (CIHR) PJT-162204 и FRN-162177 (для GCT и M.N.H.) и PJT-159454 (для QJP). Р.К. получил поддержку заработной платы от постдокторской стипендии Eyes High. М.М. получил поддержку заработной платы от Alberta Innovates и постдокторские стипендии CIHR.
Авторы заявляют об отсутствии конкурирующих финансовых интересов.
- Корреспонденцию следует направлять Roberto Colangeli по адресу roberto.colangeli{at}ucalgary.ca
Каталожные номера
- ↵
- Acharjee S,
- Verbeek M,
- Gomez CD,
- Bisht K,
- Lee B,
- Benoit L,
- Sharkey KA,
- Benediktsson A,
- Tremblay ME,
- Pittman QJ
(2018) Снижение активности микроглии и усиление передачи глутамата в базолатеральной миндалине при раннем аутоиммунитете ЦНС. Дж. Нейроски 38:9019–9033. doi:10.1523/JNEUROSCI.0398-18.2018 pmid:30185466
- ↵
- Adamec R,
- Blundell J,
- Burton P
(2004) Анксиолитические эффекты разжигающей роли анатомического расположения разжигающего электрода в ответ на разжигание правой базолатеральной миндалины.
Мозг Res 1024:44–58. doi:10.1016/j.brainres.2004.06.074 pmid:15451366 - ↵
- Амано Т,
- Duvarci S,
- Popa D,
- Paré D
(2011) Новый взгляд на контур страха: вклад ядер базального миндалевидного тела в условный страх. J Neurosci 31:15481–15489. doi:10.1523/JNEUROSCI.3410-11.2011 pmid:22031894
- ↵
- Anglada-Figueroa D,
- Quirk GJ
(2005) Поражения базальной миндалины блокируют выражение условного страха, но не угасание. J Neurosci 25:9680–9685. doi:10.1523/JNEUROSCI.2600-05.2005 pmid:16237172
- ↵
- Aroniadou-Anderjaska V,
- Fritsch B,
- Qashu F,
- Braga MF
(2008) Патология и патофизиология миндалевидного тела при эпилептогенезе и эпилепсии Эпилепсия Res 78:102–116.
doi:10.1016/j.eplepsyres.2007.11.011 pmid:18226499 - ↵
- Арониаду-Андерьяска В,
- Pidoplichko VI,
- FigueIredo TH,
- Almeida-Suhett CP,
- Prager EM,
- Braga MF
(2012). функция рецепторов GluK1. Неврология 221:157–169. doi:10.1016/j.neuroscience.2012.07.006 pmid:227
- ↵
- ТЦ Азад,
- Монори К,
- Marsicano G,
- Cravatt BF,
- Lutz B,
- Zieglgänsberger W,
- Rammes G
(2004) Схема включает сигнальную асоциативную пластичность. J Neurosci 24:9953–9961. doi:10.1523/JNEUROSCI.2134-04.2004 pmid:15525780
- ↵
- Bedse G,
- Colangeli R,
- Lavecchia AM,
- Romano A,
- Altieri F,
- Cifani C,
- Cassano T,
- Gaetani S
(2014) Роль базолатеральной миндалины в опосредовании эффектов ингибитора амидгидролазы жирных кислот URB597 на стресс-ответ.
Eur Neuropsychopharmacol 24:1511–1523. doi:10.1016/j.euroneuro.2014.07.005 pmid:25106694 - ↵
- Botterill JJ,
- Fournier NM,
- Guskjolen AJ,
- Люссье А.Л.,
- Маркс В.Н.,
- Калинчук Л.Е.
(2014) Разжигание миндалевидного тела нарушает отслеживание и задержку кондиционирования страха с параллельными изменениями экспрессии белка Fos во всем лимбическом мозге. Неврология 265:158–171. doi:10.1016/j.neuroscience.2014.01.040 pmid:24486965
- ↵
- Campos AC,
- Ferreira FR,
- Guimarães FS,
- Lemos JI
(2010) Усиление эндоканнабиноидных эффектов в вентральном гиппокампе модулирует тревожное поведение в зависимости от предыдущего опыта стресса. Неврология 167:238–246. doi:10.1016/j.neuroscience.2010.
01.062 pmid:20167262 - ↵
- Чен К.,
- Рацлифф А.,
- Хильгенберг Л.,
- Гуляш А.,
- Фройнд Т.Ф.,
- Смит М.,
- Динх Т.П.,
- 8
- Mackie K,
- Soltesz I
(2003) Долгосрочная пластичность эндоканнабиноидной передачи сигналов, вызванная фебрильными судорогами развития. Нейрон 39: 599–611. doi:10.1016/s0896-6273(03)00499-9 pmid:12
5 - ↵
- Chen SD,
- Wang YL,
- Liang SF,
- Shaw FZ. Front Behav Neurosci 10:129. doi:10.3389/fnbeh.2016.00129 pmid:27445726
- ↵
- Chevaleyre V,
- Takahashi KA,
- Castillo PE
(2006) Эндоканнабиноид-опосредованная синаптическая пластичность в ЦНС. Annu Rev Neurosci 29:37–76. doi:10.
1146/annurev.neuro.29.051605.112834 pmid:16776579 - ↵
- Коланджели Р,
- Пьеруччи М,
- Benigno A,
- Campiani G,
- Butini S,
- Di Giovanni G
(2017) Ингибитор FAAH URB597 подавляет вызванные длительной синаптической пластичностью зубчатой кости нарушения после разрядки кратковременных судорог и восстанавливает гиппокамп . Научный представитель 7:11152. doi:10.1038/s41598-017-11606-1 pmid:288
- ↵
- Коланджели Р,
- Ди Майо Р,
- Пьеруччи М,
- Deidda G,
- Casarrubea M,
- Di Giovanni G
(2019) Синергетическое действие рецепторов CB1 и 5-HT2B в предотвращении пилокарпин-индуцированного эпилептического статуса у крыс. Neurobiol Dis 125:135–145. doi:10.1016/j.nbd.2019.01.026 pmid:30716469
- ↵
- Di Maio R,
- Colangeli R,
- Di Giovanni G
(2019) WIN 55,212-2 реверсивный пилокарпин-индуцированный эпилептический статус ранние изменения взаимодействия между 5-HT2C/NMDA-рецепторами у крыс гиппокамп.
ACS Chem Neurosci 10:3296–3306. doi:10.1021/acschemneuro.9b00080 pmid:304 - ↵
- Eddy CM,
- Rickards HE,
- Cavanna AE
(2011) Влияние противоэпилептических препаратов на когнитивные функции. Ther Adv Neurol Disord 4: 385–407. doi:10.1177/1756285611417920 pmid:22164192
- ↵
- Эрлих I,
- Юмо Ю,
- Гренье Ф,
- Ciocchi S,
- Herry C,
- Lüthi A
(2009) Тормозные цепи миндалевидного тела и контроль памяти о страхе. Нейрон 62:757–771. doi:10.1016/j.neuron.2009.05.026 pmid:1
45
- ↵
- Farrell JS,
- Gaxiola-Valdez I,
- Wolff MD,
- David LS,
- Dika HI,
- Geeraert BL,
- Rachel Wang 9h,
- Sing X,
- 0018
- Spanswick SC,
- Dunn JF,
- Antle MC,
- Federico P,
- Teskey GC
(2016) .
Elife 5. doi:10.7554/eLife.19352 - ↵
- Farrell JS,
- Colangeli R,
- Wolff MD,
- Wall AK,
- Phillips TJ,
- George A,
- Federico P,
- Teskey GC
(2017) Постиктальная гипоперфузия/гипоксия обеспечивает основу для единой теории аномалий мозга, вызванных приступами, и поведенческой дисфункции. Эпилепсия 58:1493–1501. doi:10.1111/epi.13827 pmid:28632329
- ↵
- Фурнье Н.М.,
- Боттерилл Дж.Дж.,
- Маркс В.Н.,
- Гускьолен А.Я.,
- Калинчук Л.Е.
(2013) Нарушение рекрутирования нейронов, генерируемых припадками, в сети функциональной памяти взрослой зубчатой извилины после долговременного киндлинга миндалевидного тела. Exp Neurol 244:96–104. doi:10.1016/j.expneurol.2012.
11.031 pmid:23247116 - ↵
- Серый JM,
- Веккиарелли HA,
- Морена М,
- Ли ТТ,
- Хермансон ДиДжей,
- Ким АБ,
- Маклафлин Р.Дж.,
0021 Hassan Ki, - Kühne C,
- Wotjak CT,
- Deussing JM,
- Patel S,
- Hill Mn
(2015). J Neurosci 35:3879–3892. doi:10.1523/JNEUROSCI.2737-14.2015 pmid:25740517
- Haller J,
- Varga B,
- Ledent C,
- Barna I,
- Freund TF
(2004) Контекстно-зависимые эффекты нарушения гена каннабиноидов CB1 на тревожное и социальное поведение у мышей. Eur J Neurosci 19: 1906–1912. doi:10.1111/j.1460-9568.2004.03293.x pmid:15078564
- Hannesson DK,
- Pollock MS,
- Howland JG,
- Mohapel P,
- Wallace AE,
- Corcoran ME
(2008). у крыс. Эпилепсия Поведение 13:52–61. doi:10.1016/j.yebeh.2008.02.007 pmid:18337178
- Hartmann A,
- FASSINI A,
- SCOPINHO A,
- Correa FM,
- GuiMarães FS,
- LISBOA SF,
- RESSTEL LB 90ALOCAL 9.CLOCAL 9.GALCAL 9.GALCAL 9.GALCAL 9.GALCAL
- 9.SIPPOCAL
- .SIPPOCAL. в кардиоваскулярных изменениях и отсроченном тревожноподобном эффекте, вызванном острым иммобилизационным стрессом у крыс. J Psychopharmacol 33:606–614. doi:10.1177/0269881119827799 pmid:30789299
- ↵
- Helfer V,
- Deransart C,
- Marescaux C,
- Depaulis A
(1996) Возбуждение миндалевидного тела у крыс: анксиогенные последствия. Неврология 73:971–978. doi:10.1016/0306-4522(96)00081-4 pmid:8809816
- ↵
- Hill MN,
- Tasker JG
(2012) Эндоканнабиноидная передача сигналов, глюкокортикоид-опосредованная отрицательная обратная связь и регуляция гипоталамо-гипофизарно-надпочечниковой оси.
Неврология 204:5–16. doi:10.1016/j.neuroscience.2011.12.030 pmid:22214537 - ↵
- Каннер А.М.
(2016) Лечение психических и неврологических сопутствующих заболеваний при эпилепсии. Nat Rev Neurol 12:106–116. doi:10.1038/nrneurol.2015.243 pmid:26782334
- ↵
- Kanner AM,
- Barry JJ,
- Gilliam F,
- Hermann B,
- Meador KJ
(2010) качество жизни больных эпилепсией? Эпилепсия 51:1152–1158. doi:10.1111/j.1528-1167.2010.02582.x pmid:20477847
- ↵
- Кано М.,
- Оно-Шосаку Т.,
- Хашимотодани Ю.,
- Учигасима М.,
- Ватанабэ М.
(2009) Эндоканнабиноидно-медикаментозный контроль передачи. Physiol Rev 89:309–380.
doi:10.1152/physrev.00019.2008 pmid:160 - ↵
- Kasugai Y,
- Vogel E,
- Hörtnagl H,
- Schönherr S,
- Paradiso E,
- Hauschild M,
- Göbel G,
- Миленкович I,
- Peterschmitt Y,
- Tasan R,
- Sperk G,
- Shigemoto R,
- Sieghart w,
(2019) Структурное и функциональное ремоделирование ГАМКергических синапсов миндалины при обучении ассоциативному страху. Нейрон 104:781–794.e4. doi:10.1016/j.neuron.2019.08.013 pmid:31543297
- Kathuria S,
- Gaetani S,
- Fegley D,
- Valiño F,
- Duranti A,
- Tontini A,
- Mor M,
- Tarzia G,
- La Rana G,
- Calignano A ,
- Джустино А. ,
- Таттоли М.,
- Палмери М.,
- Куомо В.,
- Пиомелли Д.
(2003) Модуляция тревоги посредством блокады гидролиза анандамида. Nat Med 9:76–81. doi:10.1038/nm803 pmid:12461523
- LeDoux JE
(2000) Цепи эмоций в мозге. Annu Rev Neurosci 23:155–184. doi:10.1146/annurev.neuro.23.1.155 pmid:10845062
- Lee SH,
- Ledri M,
- Tóth B,
- Marchionni I,
- Henstridge CM,
- Dudok B,
- Kenesei K,
- Barna L,
- Szabó SI,
- Renkecz T,
- Oberoi M,
- Watanabe M,
- Limoli CL,
- Horvai G,
- Soltesz I,
- Katona I
(2015 ) Множественные формы эндоканнабиноидной и эндованиллоидной передачи сигналов регулируют тонический контроль высвобождения ГАМК. J Neurosci 35:10039–10057. doi:10.1523/JNEUROSCI.4112-14.2015 pmid:26157003
- Letzkus JJ,
- Wolff SB,
- Lüthi A
(2015) Растормаживание, схема механизма ассоциативного обучения и памяти. Нейрон 88: 264–276. doi:10.1016/j.neuron.2015.09.024 pmid:264
- Li GL,
- Winter H,
- Arends R,
- Jay GW,
- LE V,
- Young T,
- Huggins JP
- 1
(2012) Оценка Pharmacom 04457845, необратимый ингибитор гидролазы амидов жирных кислот-1, у здоровых людей. Br J Clin Pharmacol 73:706–716. doi:10.1111/j.1365-2125.2011.04137.x pmid:22044402
- ↵
- Lisboa SF,
- Borges AA,
- Nejo P,
- FASSINI A,
- GUIMARã FS FS,
- RESSTEL LB
(20151).
подобное поведение у крыс: дополнительные доказательства. Прог Нейропсихофармакол Биол Психиатрия 59:76–83. doi:10.1016/j.pnpbp.2015.01.005 pmid:255 - ↵
- Lowery-Gionta EG,
- Crowley NA,
- Bukalo O,
- Silverstein S,
- Holmes A,
- Kash TL
(2018). Нейрофармакология 139:68–75. doi:10.1016/j.neuropharm.2018.06.032 pmid:297
- ↵
- Ludányi A,
- Eross L,
- Czirják S,
- Vajda J,
- Halász P,
- Watanabe M,
- Palkovits M,
- Maglóczky Z,
- Freund TF,
- Katona I
(2008)Подавление каннабиноидного рецептора CB1 и родственных молекулярных элементов эндоканнабиноидной системы в эпилептическом гиппокампе человека. J Neurosci 28:2976–2990.
doi:10.1523/JNEUROSCI.4465-07.2008 pmid:18354002 - ↵
- Marsicano G,
- Wotjak CT,
- Azad SC,
- Bisogno T,
- Rammes G,
- Cascio MG,
- Hermann H,
- Tang J,
- Hofmann C,
- Zieglgänsberger W,
- Di Marzo V,
- Lutz B
(2002) Эндогенная каннабиноидная система контролирует угасание аверсивных воспоминаний. Природа 418:530–534. дои: 10.1038 / природа00839pmid:12152079
- ↵
- Marsicano G,
- Goodenough S,
- Monory K,
- Hermann H,
- Eder M,
- Cannich A,
- Azad SC,
- Cascio MG,
- Gutiérrez SO,
- van der Stelt M,
- Лопес-Родригес ML,
- Казанова E,
- Schütz G,
- Zieglgänsberger W,
- Di Marzo V,
- Behl C,
- Lutz B
(2003) Каннабиноидные рецепторы CB1 и защита по требованию от эксайтотоксичности.
Наука 302:84–88. doi:10.1126/science.1088208 pmid:14526074 - ↵
- Mayo LM,
- Asratian A,
- Lindé J,
- Morena M,
- Haataja R,
- Hammar V,
- Augier G,
- Hill MN,
- Heilig M
(2020a ) Повышенный уровень анандамида, усиление воспоминаний об исчезновении страха и ослабление стрессовых реакций после ингибирования амидгидролазы жирных кислот: рандомизированное контролируемое экспериментальное медицинское исследование. Биол Психиатрия 87:538–547. doi:10.1016/j.biopsych.2019.07.034 pmid:315
- ↵
- Mayo LM,
- Asratian A,
- Lindé J,
- Holm L,
- Nätt D,
- Augier G,
- Stensson N,
- Vecchiarelli HA,
- Balsevich G,
- Aukema RJ,
- Гафури Б. ,
- Спаньоло, штат Пенсильвания,
- Ли, штат Миннесота,
- Хилл, Миннесота,
- Heilig M
(2020b) Защитные эффекты повышенного содержания анандамида в отношении поведения, связанного со стрессом и страхом: трансляционные данные о людях и мышах. Мол Психиатрия 25:993–1005. doi:10.1038/s41380-018-0215-1 pmid:30120421
- ↵
- Морена М,
- Розендал Б,
- Трецца В,
- Ратано П,
- Пелосо А,
- Хауэр Д,
- Ацак П,
- Cuomo V,
- McGaugh JL,
- Schelling G,
- Campolongo P
(2014) Высвобождение эндогенных каннабиноидов в префронтально-лимбических путях влияет на консолидацию памяти во время эмоциональной тренировки. Proc Natl Acad Sci USA 111:18333–18338. doi:10.1073/pnas.1420285111 pmid:25489086
- ↵
- Морена М,
- Де Кастро V,
- Серый JM,
- Палмери М,
- Trezza V,
- Roozendaal B,
- Hill MN,
- Campolongo P
(2015) Связанное с обучением эмоциональное возбуждение формирует эндоканнабиноидную модуляцию пространственной памяти у крыс.
J Neurosci 35:13962–13974. doi:10.1523/JNEUROSCI.1983-15.2015 pmid:26468197 - ↵
- Morena M,
- Patel S,
- Bains JS,
- Hill MN
(2016a) Нейробиологические взаимодействия между стрессом и эндоканнабиноидной системой. Нейропсихофармакология 41:80–102. doi:10.1038/npp.2015.166 pmid:26068727
- ↵
- Morena M,
- Leitl KD,
- Vecchiarelli HA,
- Gray JM,
- Campolongo P,
- Hill MN
(2016b) Emotional arousal state influences the ability of amygdalar endocannabinoid signaling to modulate anxiety. Нейрофармакология 111:59–69. doi:10.1016/j.neuropharm.2016.08.020 pmid:27553121
- ↵
- Morena M,
- Aukema RJ,
- Leitl KD,
- Rashid AJ,
- Vecchiarelli HA,
- Josselyn SA,
- Hill MN
(2019) Upregulation of anandamide hydrolysis in the basolateral complex of миндалевидное тело снижает экспрессию памяти о страхе и показатели стресса и тревоги.
J Neurosci 39:1275–1292. doi:10.1523/JNEUROSCI.2251-18.2018 pmid:30573646 - ↵
- Nabavi S,
- Fox R,
- Proulx CD,
- Lin JY,
- Tsien RY,
- Malinow R
Engineering с памятью ATP и LTD (201). Природа 511:348–352. doi:10.1038/nature13294 pmid:248
- ↵
- Oliet SH,
- Баймухаметова Д.В.,
- Piet R,
- Bains JS
(2007) Ретроградная регуляция передачи ГАМК посредством тонического выброса окситоцина и эндоканнабиноидов. J Neurosci 27:1325–1333. doi:10.1523/JNEUROSCI.2676-06.2007 pmid:17287507
- ↵
- Pinel JP,
- Rovner LI
(1978) Экспериментальный эпилептогенез: индуцированная киндлингом эпилепсия у крыс.
Exp Neurol 58:190–202. doi:10.1016/0014-4886(78)-4 pmid:618743 - ↵
- Qi M,
- Morena M,
- Vecchiarelli HA,
- Hill MN,
- Schriemer DC
(2015) Надежный метод капиллярной жидкостной хроматографии/тандемной масс-спектрометрии для количественного определения нейромодулирующих эндоканнабиноидов. Rapid Commun Mass Spectro 29: 1889–1897. doi:10.1002/rcm.7277 pmid:26411510
- ↵
- Racine RJ
(1972) Изменение судорожной активности с помощью электрической стимуляции. II. Моторный приступ. Электроэнцефалогр Клин Нейрофизиол 32: 281–294. doi:10.1016/0013-4694(72)-0 pmid:4110397
- ↵
- rammes G,
- Steckler T,
- Kresse A,
- Schütz G,
- Zieglgänsberger W,
- Lutz B
(2000).
белок, связывающий элементы ответа (CREB) в переднем мозге. Eur J Neurosci 12:2534–2546. дои: 10.1046/j.1460-9568.2000.00108.x pmid:108 - ↵
- Romigi A,
- Bari M,
- Placidi F,
- Marciani Mg,
- Malaponti M,
- Torelli F,
- Izzi F,
- 111111111111111111111111111111111111111111111111111.8111111111111111111118. ............
(2010) Уровни эндоканнабиноидного анандамида в спинномозговой жидкости снижаются у пациентов с недавно диагностированной височной эпилепсией, не получавших лечения. Эпилепсия 51:768–772. doi:10.1111/j.1528-1167.2009.02334.x
- Rubino T,
- Realini N,
- Castiglioni C,
- Guidali C,
- Viganó D,
- Marras E,
- Petrosino S,
- Perletti G,
- Maccarrone M,
- Di Marzo V ,
- Parolaro D
(2008)Роль в тревожном поведении эндоканнабиноидной системы в префронтальной коре. Кора головного мозга 18:1292–1301. doi:10.1093/cercor/bhm161 pmid:179
- Schubert M,
- Siegmund H,
- Pape HC,
- Albrecht D
(2005) Изменения пластичности миндалевидного тела и гиппокампа, вызванные киндлингом. Выучить память 12:520–526. doi:10.1101/lm.4205 pmid:16204204
- Сёдзи Ю,
- Танака Э,
- Yamamoto S,
- Maeda H,
- Higashi H
(1998) Механизмы, лежащие в основе усиления возбуждающей синаптической передачи в базолатеральных нейронах миндалевидного тела разжигающей крысы. J Нейрофизиол 80:638–646. doi:10.1152/jn.1998.80.2.638 pmid: 57
- Spagnolo PA,
- Ramchandani VA,
- Schwandt ML,
- Kwako LE,
- George DT,
- Mayo LM,
- Hillard CJ,
- Heilig M
(2016) Вариация гена FAAH снижает реакцию на стресс и тяжесть симптомов у пациентов с посттравматическим стрессовым расстройством и сопутствующей алкогольной зависимостью. Alcohol Clin Exp Res 40:2426–2434. doi:10.1111/acer.13210 pmid:27716956
- Stella N,
- Schweitzer P,
- Piomelli D
(1997) Второй эндогенный каннабиноид, модулирующий долгосрочное потенцирование. Природа 388:773–778. doi:10.1038/42015 pmid:
- Teskey GC,
- Corcoran ME
(2009) Межприступные поведенческие сопутствующие заболевания в модели эпилепсии. В: Энциклопедия основных исследований эпилепсии (Schwartzkroin P, ed), стр. 1254–1260. Оксфорд: Эльзевир.
- Teskey GC,
- Corcoran ME
(2015) Kindling. В: Международная энциклопедия социальных и поведенческих наук (Wright JD, ed), Ed 2, pp 32–35. Оксфорд: Эльзевир.
- Wallace MJ,
- Blair RE,
- Falenski KW,
- Martin BR,
- DeLorenzo RJ
(2003) J Pharmacol Exp Ther 307:129–137. дои: 10.1124/jpet.103.051920:120
- Washburn MS,
- Moises HC
(1992) Электрофизиологические и морфологические свойства базолатеральных миндалевидных нейронов крыс in vitro. J Neurosci 12:4066–4079. pmid:1403101
- Ясмин Ф.,
- Коланджели Р.,
- Морена М.,
- Филипски С.,
- Ван дер Стелт М.,
- Питтман К.Дж.,
- Hillard CJ,
- Teskey GC,
- McEwen BS,
- Hill MN,
- Chattarji S
(2020). Proc Natl Acad Sci USA 117:650–655. doi:10.1073/pnas.1
- Ёсида Т,
- Учигасима М,
- Ямасаки М,
- Katona I,
- Yamazaki M,
- Sakimura K,
- Kano M,
- Yoshioka M,
- Watanabe M
(2011) Unique inhibitory synapse with particularly rich endocannabinoid signaling machinery on pyramidal neurons in basal миндалевидное ядро. Proc Natl Acad Sci USA 108:3059–3064. doi:10.1073/pnas.1012875108 pmid:21282604
- Чжу П.Дж.,
- Любовник DM
(2005) Ретроградная эндоканнабиноидная передача сигналов в препарате постсинаптического нейрона/синаптического бутона из базолатеральной миндалины. J Neurosci 25:6199–6207. doi:10.1523/JNEUROSCI.1148-05.2005 pmid:15987949
- Zimmermann T,
- Bartsch JC,
- Beer A,
- Lomazzo E,
- Guggenhuber S,
- Lange MD,
- Bindila 9
Pa,
- 80021 Lutz B
(2019) Нарушение передачи сигналов анандамида/пальмитоилэтаноламида в глутаматергических нейронах гиппокампа изменяет синаптическую пластичность, обучение и эмоциональные реакции. Нейропсихофармакология 44:1377–1388. doi:10.1038/s41386-018-0274-7 pmid:30532004
View Abstract
Вернуться к началу
Выражения лица не выдают эмоций
Движения вашего лица передают ваши эмоции другим людям? Если вы думаете, что ответ положительный, подумайте еще раз. Этот вопрос находится в стадии ожесточенных дискуссий. Некоторые эксперты утверждают, что люди во всем мире делают определенные, узнаваемые лица, которые выражают определенные эмоции, например улыбаются от счастья, хмурятся от гнева и задыхаются с расширенными глазами от страха. Они ссылаются на сотни исследований, демонстрирующих, что улыбка, хмурый взгляд и т. д. являются универсальным выражением эмоций на лице. Они также часто цитируют книгу Чарльза Дарвина 1872 года 9.1050 The Expression of the Emotions in Man and Animals , чтобы поддержать утверждение о том, что универсальные выражения возникли в результате естественного отбора.
Другие ученые указывают на массу контрдоказательств, показывающих, что движения лица во время эмоций слишком сильно различаются, чтобы быть универсальными маяками эмоционального значения. Люди могут улыбаться с ненавистью, когда замышляют падение своего врага, и хмуриться от восторга, когда слышат плохой каламбур. В меланезийской культуре задыхающееся лицо с широко раскрытыми глазами является символом агрессии, а не страха. Эти эксперты говорят, что предполагаемые универсальные выражения просто представляют собой культурные стереотипы. Чтобы было ясно, обе стороны в дебатах признают, что движения лица меняются в зависимости от данной эмоции; разногласие заключается в том, достаточно ли единообразия, чтобы определить, что кто-то чувствует.
Этот спор не только академический; результат имеет серьезные последствия. Сегодня вам могут отказать в приеме на работу, потому что так называемая система чтения эмоций, наблюдающая за вами на камеру, использует искусственный интеллект для неблагоприятной оценки ваших движений лица во время интервью. В суде США судья или присяжные иногда могут вынести более суровый приговор, вплоть до смертной казни, если посчитают, что на лице подсудимого не было раскаяния. Детей в дошкольных учреждениях по всей стране учат распознавать улыбки как счастье, хмурые взгляды как гнев и другие выразительные стереотипы из книг, игр и плакатов с бестелесными лицами. А для детей в спектре аутизма, некоторые из которых с трудом воспринимают эмоции других, эти учения не способствуют лучшему общению.
Так кто же прав? Ответ включает в себя невольного врача, научную ошибку и многовековую неверную интерпретацию работ Дарвина. По иронии судьбы, его собственные наблюдения предлагают мощное решение, которое трансформирует современное понимание эмоций.
Предположение об универсальных выражениях лица восходит к нескольким источникам, в первую очередь к набору фотографий французского врача XIX века Гийома-Бенжамина-Амана Дюшенна. На заре фотографии Дюшен электрически стимулировал лицевые мышцы людей и фотографировал их сокращения.
Его фотографии вдохновили Дарвина на предположение в Expression , что определенные движения лица являются универсальными признаками эмоций. В счастье, писал Дарвин, люди улыбаются. В печали они хмурятся. Как обычно рассказывают эту историю, Дарвин обнаружил, что эмоции имеют врожденные, биологически обоснованные выражения, которые создаются и признаются повсеместно и разделяются с другими животными. В этой истории движения лица представлены как своего рода сигнальная система, с помощью которой вы можете смотреть на лицо человека, определять его эмоциональное состояние и получать важную информацию, которая поможет вам — и им — быть живыми и здоровыми.
По крайней мере, так может показаться. Большинство свидетельств показывает, что Дарвин был не прав, и его ошибка была дурацкой. В реальной жизни люди выражают данную эмоцию с огромной вариативностью. В гневе, например, люди в городских культурах хмурятся (или делают некоторые мимики для хмурого взгляда) только около 35 процентов времени, согласно метаанализу исследований, измеряющих движения лица во время эмоции. Хмурый взгляд также не связан с гневом, потому что люди хмурятся по другим причинам, например, когда они концентрируются или когда у них газы. Такая же огромная вариация наблюдается для каждой изучаемой эмоции — и для любого другого измерения, которое якобы говорит нам о чьем-то эмоциональном состоянии, будь то его физиология, голос или активность мозга.
Эмоции Системы искусственного интеллекта, таким образом, не обнаруживают эмоции. Они обнаруживают физические сигналы, такие как движения мышц лица, а не психологическое значение этих сигналов. Смешение движения и смысла глубоко укоренилось в западной культуре и науке. Примером может служить недавнее громкое исследование, в котором машинное обучение применялось к более чем шести миллионам интернет-видео с лицами. Оценщиков-людей, обучавших систему искусственного интеллекта, попросили маркировать движения лица в видео, но единственные метки, которые им давали использовать, были словами эмоций, такими как «сердитый», а не физическими описаниями, такими как «хмурый взгляд». Более того, не было объективного способа подтвердить, что чувствовали в эти моменты анонимные люди на видео.
Имеются также убедительные доказательства того, что движения лица являются лишь одним из многих сигналов в гораздо большем массиве контекстуальной информации, которую воспринимает наш мозг. Покажите людям гримасничающее лицо в одиночестве, и они могут ощутить боль или разочарование. Но покажите такое же лицо бегуну, пересекающему финишную черту забега, и та же самая гримаса передаст триумф. Лицо часто является более слабым сигналом внутреннего состояния человека, чем другие сигналы массива.
Выражение Дарвина предполагает, что случаи определенной эмоции, такой как гнев, имеют общую неизменную физическую причину или состояние — сущность, — которая делает их похожими, даже если они имеют поверхностные различия. Ученые предложили множество сущностей, некоторые из которых легко увидеть, например, движения лица, а другие, такие как сложные, переплетенные закономерности частоты сердечных сокращений, дыхания и температуры тела, наблюдаемые только с помощью специализированных приборов. Эта вера в сущности, называемая эссенциализмом, является неотразимой интуицией. Еще и потому, что практически невозможно доказать, что сущности не существует. Люди, которые верят в сущности, но не соблюдают их, несмотря на неоднократные попытки, часто все равно продолжают в них верить. Исследователи, в частности, склонны оправдывать свою веру, предполагая, что инструментов и методов еще недостаточно для обнаружения сущности, которую они ищут.
Решение этой головоломки можно найти в более известной книге Дарвина «Происхождение видов », написанной за 13 лет до «Выражение ». По иронии судьбы, он прославился тем, что помог биологии «избежать парализующей хватки эссенциализма», по словам известного биолога Эрнста Майра. До публикации Origin ученые считали, что каждый биологический вид имеет идеальную форму, созданную Богом, с определяющими свойствами — сущностью — которые отличают его от всех других видов. Думайте об этом как о версии биологии для «выставки собак». На выставке собак каждый участник оценивается в сравнении с гипотетической идеальной собакой. Отклонение от идеала считается ошибкой. Дарвина Происхождение радикально предположил, что вид представляет собой обширную популяцию различных индивидуумов, не имеющих сущности в своей основе. Идеальной собаки не существует — это статистическая сводка множества разных собак. Вариация — это не ошибка; это необходимый ингредиент для естественного отбора со стороны окружающей среды. Однако когда дело дошло до эмоций, Дарвин стал жертвой эссенциализма, проигнорировав свое самое важное открытие.
Сила эссенциализма привела Дарвина к некоторым прекрасно смешным идеям об эмоциях, в том числе к тому, что эмоциональный дисбаланс может вызывать вьющиеся волосы и что насекомые выражают страх и гнев, лихорадочно потирая части тела друг о друга.
Эссенциализм, похоже, также соблазняет разработчиков эмоциональных систем искусственного интеллекта следовать за Дарвином по этому удобному пути, предполагая, что эмоции развились в результате естественного отбора для выполнения важных функций. Но если вы действительно прочитаете Выражение , вы обнаружите, что Дарвин почти не упоминал естественный отбор. Он также не писал, что мимика является функциональным продуктом эволюции. На самом деле он писал обратное: улыбки, хмурые взгляды, расширение глаз и другие физические выражения были «бесцельными» — рудиментарными движениями, которые больше не выполняют никакой функции. Он делал это заявление более 10 раз в Выражение . Для Дарвина выражение эмоций было убедительным доказательством того, что люди — это животные и что мы эволюционировали. По его логике, если мы разделяем выражения с другими животными, но эти выражения функционально бесполезны для нас, они, должно быть, произошли от давно ушедшего общего предка, для которого выражения были полезны.
Выражение неправильно цитируется уже более 100 лет. Как это случилось? Я нашел ответ в работах психолога начала 20-го века Флойда Олпорта. В свои 19В книге «Социальная психология » Оллпорт сделал радикальный вывод из сочинений Дарвина, утверждая, что выражения у новорожденных начинаются как рудиментарные, но быстро приобретают полезные социальные функции. Он писал: «Вместо биологически полезной реакции, имеющейся у предка, и выразительного остатка у потомка, мы рассматриваем обе эти функции как имеющиеся у потомка, причем первая служит основой, из которой развивается последняя».
Идея Олпорта, хотя и неверная, была приписана Дарвину и охотно принята учеными-единомышленниками. Теперь они могли писать о мимике как об универсальной и претендовать на звание наследников неприступного Чарльза Дарвина. Одним предложением Олпорт исказил западное понимание эмоций не только в науке, но и в юриспруденции, медицине, в глазах общественности, а теперь и в системах искусственного интеллекта эмоций.
Тем не менее, эта научная сказка имеет счастливый конец, потому что есть название для вида вариаций, которые мы наблюдаем в реальных проявлениях эмоций. Это то же самое изменение, которое сам Дарвин наблюдал у животных. В Origin, Дарвин описал вид животных как совокупность различных индивидуумов, не имеющих биологической сущности в своей основе. Это ключевое наблюдение стало широко известно как популяционное мышление, и оно подтверждается современными исследованиями в области генетики.
Популяционное мышление произвело революцию в биологии за последнее столетие, а теперь оно революционизирует науку об эмоциях. Подобно виду, данная эмоция, такая как страх, горе или восторг, представляет собой обширную совокупность разнообразных экземпляров. Люди действительно могут широко раскрывать глаза и задыхаться от страха, но они также могут хмуриться от страха, плакать от страха, смеяться в лицо страху и, в некоторых культурах, даже засыпать в страхе. Сути нет. Изменчивость — это норма, и она тесно связана с физиологией и ситуацией человека, точно так же, как изменчивость вида связана с окружающей средой, в которой живут его представители9.0003
Все большее число исследователей эмоций более серьезно относятся к мышлению населения и выходят за рамки эссенциалистских идей прошлого. Пришло время сторонникам искусственного интеллекта эмоций и компаниям, которые производят и продают эти продукты, прекратить шумиху и признать, что движения лицевых мышц не всегда сопоставляются с конкретными эмоциями. Очевидно, что одна и та же эмоция может сопровождать разные движения лица и что одни и те же движения лица могут иметь разное (или не иметь) эмоциональное значение. Разнообразие, а не единообразие, является правилом.
Выражение Дарвина лучше всего рассматривать как исторический текст, а не как исчерпывающее научное руководство. Это приводит к более глубокому уроку: наука не является истиной авторитетом. Наука — это количественная оценка сомнения путем многократного наблюдения в различных контекстах. Даже самые выдающиеся ученые могут ошибаться. К счастью, ошибки являются частью научного процесса. Это возможности для открытий.
Это статья с мнением и анализом, и взгляды, выраженные автором или авторами, не обязательно совпадают с мнениями Научный американец.
ОБ АВТОРАХ
Лиза Фельдман Барретт — профессор психологии Северо-восточного университета. Она является автором нескольких книг, в том числе «Как создаются эмоции: тайная жизнь мозга» . Подпишитесь на нее в Твиттере @LFeldmanBarrett.
Последние статьи Лизы Фельдман Барретт
- О природе страха
Лимбическая система: миндалевидное тело (раздел 4, глава 6) Neuroscience Online: электронный учебник по нейронаукам | Кафедра нейробиологии и анатомии
6.1 Миндалевидное тело – общие положения
Миндалевидное тело является интегративным центром эмоций, эмоционального поведения и мотивации. Если мозг перевернуть вверх дном, конец структуры, продолжающейся с гиппокампом, называется крючком. Если вы оторвете ункус, вы обнаружите миндалевидное тело, которое примыкает к передней части гиппокампа. Как и в случае с гиппокампом, основные пути взаимодействуют в двух направлениях и содержат как эфферентные, так и афферентные волокна.
Рисунок 6.1 |
6. 2 Входы в миндалевидное тело
Рисунок 6.2 |
Как и в случае с гиппокампом, волокна, несущие входы в миндалину, практически во всех случаях сочетаются с волокнами, несущими выходы из миндалины.
Миндалевидное тело получает информацию от всех органов чувств, а также от внутренних органов. Поскольку миндалевидное тело играет очень важную роль в эмоциональном обучении, неудивительно, что висцеральные входы являются основным источником входных данных. Висцеральные входы исходят от гипоталамуса, области перегородки, орбитальной коры и парабрахиального ядра. Обонятельная сенсорная информация поступает из обонятельной луковицы. Слуховая, зрительная и соматосенсорная информация поступает из височной и передней поясной коры.
Рисунок 6.3 |
6.3 Основные выходные пути миндалевидного тела
- Вентральный миндалевидный путь
- Терминальная полоска
- Непосредственно в гиппокамп
- Непосредственно в энторинальную кору
- Непосредственно к дорсомедиальному ядру таламуса
6.4 Вентральный миндалевидный путь
Вентральный миндалевидный путь. Термин «фугал» происходит от слова «фуге» — отгонять — как в слове «беглец». Этот путь продолжается к переднему обонятельному ядру, переднему продырявленному веществу, грушевидной коре, орбитофронтальной коре, передней поясной коре и вентральному полосатому телу. Вентральный стриатум включает в себя часть хвостатого ядра, скорлупу и прилежащее ядро перегородки (ядро, лежащее на перегородке). Отростки вентрального полосатого тела являются звеньями цепи базальных ганглиев, которые важны для ассоциативного обучения «стимул-реакция». Вентральный миндалевидно-фугальный путь также соединяется с гипоталамусом и ядром перегородки, но основная связь миндалевидного тела с гипоталамусом и ядром перегородки осуществляется через терминальную полоску.
Вентральный миндалевидный путь важен, потому что он является связующим звеном, посредством которого мотивация и побуждения через лимбическую систему могут влиять на реакцию. Это также связь, по которой узнаются ответы. В данном случае это звено, посредством которого происходит ассоциативное обучение. Именно здесь ответы связаны с аппетитными и аверсивными последствиями, то есть с вознаграждением и наказанием.
Три упрощения:
- Конечная полоска по форме, функциям и расположению аналогична своду пути гиппокампа. Таким образом, по аналогии можно сказать, что конечная полоска относится к миндалевидному телу так же, как свод к гиппокампу. Stria — это латинское слово, означающее линию, канавку или полосу. Относится к слову «полосатый».
- Конечная полоска соединяется только с подкорковыми структурами. (Соединение со структурами коры осуществляется через вентральный миндалевидный путь.)
- Конечная полоска перекрывается с вентральным миндалевидным путем в том смысле, что он также соединяется с ядрами перегородки и гипоталамусом и, таким образом, образует петлю.
Подробнее о сходстве со сводом:
Как и свод, терминальная полоска имеет прекомиссуральные и посткомиссуральные ответвления относительно передней комиссуры. Прекомиссуральная ветвь идет в септальную область. Это именно то, что делает fornix. Посткомиссуральная ветвь идет к гипоталамусу. Это именно то, что делает fornix. В то время как постспаечная ветвь свода проецируется на сосцевидные тела гипоталамуса, посткомиссуральная ветвь концевой полоски проецируется на латеральное ядро и вентрально-медиальное ядро гипоталамуса.
Как и в своде, некоторые волокна входят в переднюю комиссуру креста на противоположной стороне. Как и в случае двух гиппокампов, сообщающихся друг с другом через переднюю спайку, две миндалины сообщаются друг с другом через переднюю спайку.
Конечная полоска также проецируется на хабенулу, которая является частью эпиталамуса.
Центральное ядро миндалевидного тела производит вегетативные компоненты эмоций (например, изменения частоты сердечных сокращений, артериального давления и дыхания) главным образом через выходные пути к латеральному гипоталамусу и стволу головного мозга.
Центральное ядро миндалевидного тела также обеспечивает сознательное восприятие эмоций, главным образом, через вентральный выходной путь миндалевидного тела к передней поясной коре, орбитофронтальной коре и префронтальной коре.
6.5 Подробнее о функциях миндалевидного тела
Стимуляция миндалевидного тела вызывает сильные эмоции, такие как агрессия или страх.
Ирритативные поражения височной эпилепсии имеют эффект стимуляции миндалевидного тела. В своей крайней форме ирритативные поражения височной эпилепсии могут вызывать панические атаки. Панические атаки — это короткие спонтанно повторяющиеся приступы ужаса, которые вызывают ощущение надвигающейся катастрофы без четко определенной причины. ПЭТ-сканирование показало увеличение притока крови к парагиппокампальной извилине, начиная с правой парагиппокампальной извилины. Аналогичное, но ослабленное увеличение кровотока происходит во время приступов тревоги.
Деструктивные поражения, такие как абляция миндалины, вызывают эффект, противоположный раздражающим поражениям височной эпилепсии. Деструктивные поражения миндалины вызывают у животных укрощенность, а у человека — безмятежное спокойствие, характеризующееся уплощением аффекта. Поражения миндалевидного тела могут возникать в результате болезни Урбаха-Вите, когда кальций откладывается в миндалевидном теле. Если это заболевание возникает в раннем возрасте, то эти пациенты с двусторонним поражением миндалины не могут различать эмоции по выражению лица, но их способность идентифицировать лица сохраняется. Анатомическая область распознавания лиц и памяти находится в мультимодальной ассоциативной области нижневисочной коры. Это хороший пример того, как эмоции в одной области (миндалевидное тело) связаны с восприятием в другой области (нижневисочной коре), создавая интенсивную эмоционально заряженную память.
Рисунок 6.4. Результаты фМРТ |
Уплощение аффекта является одним из симптомов ранее упомянутого синдрома Клювера-Бьюси, при котором у обезьян удаляли все височные доли. На самом деле было показано, что именно поражения миндалевидного тела в первую очередь ответственны за уплощение аффекта. Эта работа в конечном итоге привела к психохирургической технике префронтальной лоботомии. Вспомните фильм с Джеком Николсоном «Пролетая над гнездом кукушки». Префронтальная кора входит в миндалевидное тело. Разъединение этого входа вызывает уплощение аффекта, которое считалось желательным у пациентов с шизофренией, которые были агрессивно агрессивны или эмоционально возбуждены.
Миндалевидное тело объединяет множество различных сенсорных сигналов. Подобно гиппокампу, он сочетает в себе внешние и внутренние раздражители. Каждая сенсорная модальность имеет вход. Они интегрированы с соматосенсорными и висцеральными входами — здесь вы получаете свою «инстинктивную реакцию». Связь между префронтальной корой, областью перегородки, гипоталамусом и миндалевидным телом, вероятно, дает нам наши внутренние ощущения, эти субъективные ощущения, о том, что хорошо, а что плохо.
Здесь также соединяются память и эмоции. Когда награда особенно сладка, такое поведение и ассоциация могут длиться всю жизнь. Так же и травма и унижение наказания могут запомниться надолго.
6.6 Обусловливание страхом: пример роли миндалевидного тела в обучении
Другим примером связи эмоций с некоторым перцептивным опытом является обусловливание страхом. В этом примере сенсорный опыт является слуховым, а не визуальным, как в эмоциях лиц. Многое из того, что мы знаем об миндалевидном теле и его роли в эмоциональном обучении и памяти, исходит из обусловленности страхом, в основном, но не исключительно, с животными. Это пример классической обусловленности или павловской обусловленности. В классических опытах, проведенных Павловым на рубеже веков, звучал нейтральный раздражитель — звонок, и через короткий промежуток времени собаке клали в рот пищевой порошок — безусловный раздражитель. После нескольких таких пар у собаки выделялась слюна на звук колокольчика. Важнейшим аспектом классического обусловливания является то, что это сочетание двух стимулов. Для получения награды ответ не требуется. При обусловливании страха организм слышит шум или видит визуальный стимул. Через несколько секунд он получает легкий шок. Реакции включают замирание, повышенное кровяное давление и частоту сердечных сокращений, и он становится нервным — легко вздрагивает.
Рисунок 6.5
| ||
Рисунок 6.6 (вверху) и 6.7 (внизу) |
Пути от таламуса к миндалевидному телу особенно важны для эмоционального обучения. Выходные пути из центрального ядра миндалевидного тела образуют обширные связи со стволом мозга для эмоциональных реакций и обширные связи с областями коры через базальное ядро. Считается, что холинергические проекции от базального ядра к коре возбуждают кору.
Следующая диаграмма предоставляет дополнительную информацию о выходных сигналах, контролируемых миндалевидным телом во время кондиционирования страха.
Рисунок 6. |
Были обнаружены некоторые пути формирования условного рефлекса страха, и это горячая тема для исследований в нейробиологии. Если, например, поврежден путь слуховой коры, базовое обусловливание страха не изменяется, но изменяется различение. В процедуре различения один звук сочетается с ударом, а другой звук не сочетается с ударом. Животные должны были полагаться исключительно на таламус и миндалевидное тело для обучения, и они не могли научиться различению; по-видимому, эти два стимула были неразличимы.
Итак, кора головного мозга не нужна для простого обусловливания страха; вместо этого он позволяет нам узнавать объект по зрению или звуку — интерпретировать окружающую среду.
Таким образом, пути от сенсорного таламуса обеспечивают лишь грубое восприятие мира, но поскольку они включают только одно нервное звено, они являются быстрыми путями. Почему FAST может быть важен? Нужна быстрая реакция на потенциальную опасность. Путь таламус-миндалевидное тело дает нам это, а также может подготовить миндалину к получению более тщательно обработанной информации от коры.
С другой стороны, проводящие пути, идущие от коры, дают подробные и точные представления об окружающей среде. Поскольку эти пути имеют несколько нейронных связей, по сравнению с ними они медленные.
Если, например, мы видим тонкую изогнутую фигуру за деревом, гораздо лучше отпрыгнуть назад и позже узнать, что это садовый шланг, чем не суметь быстро отпрыгнуть назад, если это была змея. Позже у нас будет достаточно времени, чтобы подумать о том, как глупо было испугаться в нашем собственном безопасном саду, где нет змей.
Рисунок 6.9 | Рисунок 6.10 |
Корковые и подкорковые пути обусловливания страха. | |
Зрительные стимулы, вызывающие страх, быстро обрабатываются таламусом, и эта информация передается в миндалевидное тело (красное), производя быструю реакцию (зеленое) на опасность. Таламус также передает информацию в кору, чтобы можно было сделать более осторожные (и более медленные) суждения о реальной потенциальной опасности.
Миндалевидное тело вовлечено в эмоциональное обучение, приносящее удовольствие, а также в эмоциональное обучение, связанное со страхом. Рассмотрим инструментальное обучение. В отличие от классического обусловливания, когда два стимула объединены в пары, при инструментальном обусловливании реакции сопровождаются вознаграждением, а ассоциации стимул-реакция изучаются. Таким образом, есть три события: стимул, реакция и вознаграждение. Стало ясно, что все три парные комбинации усваиваются в инструментальной обусловленности. В дело вступает миндалевидное тело, так как поражения базолатеральных ядер миндалины нарушают связь со стимулом и атрибутами вознаграждения пищи.
Эта система памяти миндалевидного тела служит примером систем памяти в целом. Создание воспоминаний является функцией всей сети, а не какого-либо отдельного компонента. Миндалевидное тело участвует в примитивной эмоциональной памяти, которая, вероятно, сохранилась в ходе эволюции. Согласно схеме систем памяти (например, Нольте, стр. 577), декларативная память опосредована гиппокампом и корой. Но, как и в случае с корой, поражения гиппокампа мало влияют на формирование условного рефлекса страха, за исключением различения внешних раздражителей.
Исследование пациентов с повреждением миндалевидного тела, гиппокампа или того и другого ясно демонстрирует отличительную роль этих двух структур в памяти. Этим пациентам показывали слайды зеленого, синего, желтого или красного цветов. После нескольких цветов раздался громкий и пугающий звук рога. Регистрировались вегетативные ответы (через записи GSR) для определения обучения. Пациенты с миндалевидным телом не привыкли к цветам, сопровождаемым громким рогом. Но когда их спросили, сколько цветов было представлено и за какими следовал рог, их воспоминания были правильными. То есть у них была явная память о событиях. С другой стороны, пациенты с гиппокампом демонстрировали обучение и обусловливание цветов, за которыми следовал рог, но не могли вспомнить, какие именно. То есть у них была имплицитная память о событиях. Пациенты с обоими типами поражений не демонстрировали никакой обусловленности и не имели явных воспоминаний о том, за какими цветами следует рог. Глава «Обучение и память» расскажет больше о явной памяти и гиппокампе.
Тревога диссоциирует адаптивные функции сенсорных и моторных реакций на социальные угрозы
Аннотация
Эффективное обнаружение и реакция на негативные сигналы в окружающей среде необходимы для выживания. В социальных ситуациях эти сигналы часто неоднозначны и могут подразумевать разные уровни угрозы для наблюдателя, что делает их распознавание восприимчивым к контекстуальным сигналам, таким как направление взгляда при оценке выражений эмоций на лице. Однако механизмы, лежащие в основе таких контекстуальных эффектов, остаются плохо изученными. Путем компьютерного моделирования человеческого поведения и электрической активности мозга мы демонстрируем, что направление взгляда повышает перцептивную чувствительность к эмоциям, сигнализирующим об угрозе — гнев в паре с прямым взглядом и страх в паре с отведенным взглядом. Этот эффект возникает одновременно в вентральной лицевой и дорсальной моторной коре через 200 мс после предъявления лица, диссоциирует у разных людей в зависимости от тревоги и не отражает повышенного внимания к эмоциям, сигнализирующим об угрозе. Эти результаты показывают, что угроза настраивает нейронную обработку быстрым, избирательным, но независимым от внимания образом в сенсорных и двигательных системах для различных адаптивных целей.
https://doi.org/10.7554/eLife.10274.001
Дайджест eLife
Выражения лица могут передавать важные социальные сигналы, и понимание этих сигналов может иметь важное значение для выживания в угрожающих ситуациях. Прошлые исследования выявили изменения в активности и поведении мозга в ответ на определенные социальные угрозы, но неясно, как мозг обрабатывает информацию о выражениях лица других людей, чтобы идентифицировать эти угрозы. Здесь Эль Зейн, Вайарт и Грез стремились определить, как сигналы угрозы представлены в человеческом мозгу.
В эксперименте использовался метод, называемый электроэнцефалографией, для записи мозговой активности у здоровых людей-добровольцев, когда они изучали выражения гнева и страха на лицах. Эль Зейн, Вайарт и Грез обнаружили, что эмоции, сигнализирующие наблюдателю об угрозе, лучше представлены в определенных областях мозга, включая те, которые контролируют действия, в течение доли секунды после того, как добровольцу было показано выражение лица. Более того, реакция областей мозга, контролирующих действия, была выше у добровольцев с более высоким уровнем тревожности, что подчеркивает роль тревожности в быстром реагировании на социальные угрозы в окружающей среде.
Выводы Эль Зейна, Вайарта и Грезеса показывают, что социальные угрозы могут изменять активность мозга очень быстро и более избирательно, чем считалось ранее. Будущая задача состоит в том, чтобы выяснить, могут ли другие аспекты угрожающей среды стимулировать аналогичное увеличение мозговой активности.
https://doi.org/10.7554/eLife.10274.002
Введение
Решения о восприятии основаны на сочетании слабых и/или неоднозначных образцов сенсорных данных. Точность этого процесса принятия решений особенно важна для интерпретации негативных сигналов, которые требуют быстрой и адаптивной реакции. В социальной сфере определение эмоционального состояния сородича – например, он/она зол или напуган? – редко зависит исключительно от черт лица, которые обычно неоднозначны и могут подразумевать разные уровни угрозы для наблюдателя. Известно, что окружающие сигналы, такие как направление взгляда и поза тела, действуют как контекстуальная информация во время распознавания эмоций (Righart and de Gelder, 2008; Barrett and Kensinger, 2010; Aviezer et al. , 2011). В частности, обнаружение гнева представляет непосредственную угрозу для наблюдателя в сочетании с прямым взглядом; напротив, именно в сочетании с отведенным взглядом страх отмечает присутствие (и, возможно, локализацию) угрозы в окружающей среде (Sander et al., 2007). Было показано, что эти сигнализирующие об угрозе комбинации направления взгляда и эмоций лучше распознаются и оцениваются как более интенсивные, чем другие комбинации (Adams and Kleck, 2003, 2005; Graham and LaBar, 2007; Sander et al., 2007; Bindemann et al. ., 2008 г.; Н’Диайе и др., 2009 г.), и это зависит от уровня тревожности людей (Ewbank et al., 2010). Однако вычислительные механизмы, лежащие в основе приоритизации информации, сигнализирующей об угрозе, остаются неустановленными.
Классическая теория принятия решений различает два класса механизмов, с помощью которых контекстная информация, такая как направление взгляда, может влиять на распознавание негативных эмоций. Направление взгляда могло исказить интерпретацию негативных выражений лица в пользу эмоции, сигнализирующей о более высокой угрозе в данном контексте — гнева для прямого взгляда, страха для отведенного взгляда. В теоретических терминах обнаружения сигналов (Green and Swets, 1966; Macmillan and Creelman, 2004), этот эффект будет соответствовать аддитивному сдвигу критерия принятия решения в зависимости от направления взгляда. Однако направление взгляда может также увеличить перцептивную чувствительность к чертам лица, диагностирующим эмоцию, сигнализирующую о более высокой угрозе. В отличие от первой версии, этот эффект будет соответствовать мультипликативному усилению сигналов, сигнализирующих об угрозе, в процессе принятия решений. В то время как эти два описания предсказывают сходное влияние направления взгляда на распознавание эмоций, сигнализирующих об угрозе, эффект предвзятости будет максимальным для нейтральных (безэмоциональных) выражений, тогда как эффект чувствительности будет максимальным при низкой силе эмоций (рис. 1).
Прогнозы модели влияния направления взгляда на категоризацию эмоций.
Левая панель: предсказание влияния направления взгляда на предвзятость решения. Верхняя левая панель: если направление взгляда искажает интерпретацию негативных выражений лица в пользу эмоций, сигнализирующих о более высокой угрозе, прямой взгляд дополнительно смещает выбор выбора в сторону гнева. Нижняя левая панель: прогнозируемая психометрическая функция будет соответственно смещена влево для прямого взгляда, поскольку участники будут склонны интерпретировать лица, демонстрирующие прямой взгляд, как сердитые. Максимальные эффекты будут проявляться для нейтральных (безэмоциональных) выражений, выделенных заштрихованной серой областью на оси эмоций, которая представляет собой разницу между двумя психометрическими функциями для прямого и отведенного взгляда. Правая панель: предсказание влияния направления взгляда на перцептивную чувствительность. Верхняя правая панель: если направление взгляда увеличивает чувствительность к чертам лица, диагностирующим эмоцию, сигнализирующую о более высокой угрозе, прямой взгляд теперь будет мультипликативно повышать обработку гневного выражения, отображающего прямой взгляд. Нижняя правая панель: предсказанная психометрическая функция теперь будет показывать повышенный наклон для эмоций, сигнализирующих об угрозе, с максимальным эффектом при низкой силе эмоций (как показано в заштрихованной серой области на оси эмоций).
Здесь мы провели арбитраж между этими двумя возможными версиями, записав сигналы электроэнцефалограммы (ЭЭГ) человека, в то время как участники классифицировали выражение лица как выражение гнева или страха. Мы манипулировали силой эмоций, представляя «трансформированные» выражения лица в диапазоне от нейтрального до сильного гнева или страха, а также контекстную информацию, сочетая выражения лица с прямым или отведенным взглядом. Параметрический контроль над силой эмоций позволил подобрать теоретические модели принятия решений к поведенческим и нейронным данным, чтобы выбрать между предвзятостью и чувствительностью оценок влияния угроз на распознавание эмоций. На нейронном уровне в предыдущих исследованиях сообщалось о взаимодействии между эмоциями и направлением взгляда через 200 мс после предъявления лица (Sato et al., 2004; N’Diaye et al., 2009).; Адамс и др., 2012 г.; Conty et al., 2012), но не смог охарактеризовать вычислительный механизм, ответственный за эти эффекты. Здесь мы применили управляемые моделью регрессии сигналов ЭЭГ в одной попытке, чтобы определить, усиливается ли нейронное «кодирование» эмоций, сигнализирующих об угрозе, в вентральных лицевых избирательных и/или дорсальных моторных областях (El Zein et al., 2015). и опосредовано ли это улучшение повышенным нисходящим вниманием к чертам лица, сигнализирующим об угрозе. Поскольку люди с высокой тревожностью проявляют повышенную чувствительность к угрозам, а также к негативным сигналам в целом (Bishop, 2007; Cisler and Koster, 2010), мы дополнительно оценили нейронные механизмы, с помощью которых тревога влияет на обнаружение социальных угроз и реакцию на них.
Результаты
Поведение
В каждом испытании участникам предъявляли лица, выражающие страх или гнев различной силы эмоции (7 уровней силы эмоции для каждой эмоции), и они должны были классифицировать отображаемую эмоцию (рис. 2). Важно отметить, что направлением взгляда (прямым или отведенным) манипулировали независимо от отображаемой эмоции совершенно имплицитным образом, поскольку оно никогда не упоминалось испытуемым и не имело отношения к задаче категоризации эмоций. Тем не менее, в дополнение к ожидаемому увеличению производительности категоризации в зависимости от силы эмоций (F 6,138 = 187,3, p<0,001), направление взгляда сильно взаимодействовало с отображаемой эмоцией при выполнении (F 1,23 = 21,2, p<0,001). Выражения гнева лучше классифицируются в паре с прямым взглядом ( t 23 = 4,3, p<0,001), тогда как выражение страха лучше классифицируется в паре с отведенным взглядом ( t 23 = — 3,4, р<0,01; рис. 3а). Эти комбинации взгляда и эмоций, гнева в паре с прямым взглядом и страха в паре с отведенным взглядом связаны с более высокой угрозой для наблюдателя (Сандер и др., 2007), хотя и разной природы. В случае гнева направление взгляда указывает на цель угрозы, а в случае страха направление взгляда сигнализирует об ее источнике. Тем не менее, точно так же, как сочетание гнева с прямым взглядом является более угрожающим/уместным, чем с отведенным взглядом, страх более опасен в сочетании с отведенным взглядом, чем с прямым взглядом. Эти две комбинации, прямой гнев и предотвращенный страх, будут, таким образом, помечены как комбинации УГРОЗА+, в отличие от комбинаций УГРОЗА- (т. е. гнев в паре с отведенным взглядом и страх в паре с прямым взглядом).
Стимулы и методика эксперимента.
( a ) Примеры морфированных выражений для одной идентичности: морфы от нейтральных до интенсивных испуганных/сердитых выражений, свидетельствующих о той или иной эмоции. Стимулы отображали либо отведенный, либо прямой взгляд. Условия УГРОЗА+ (оранжевые) соответствуют комбинациям взгляда и эмоций, которые сигнализируют о более высокой угрозе для наблюдателя по сравнению с условиями УГРОЗА- (зелеными). ( b ) После фиксации выражение лица появлялось на 250 мс, после чего участник должен был в течение 2 с указать, выражает ли лицо гнев или страх. Никакой обратной связи не было предоставлено после ответа.
Повышенная точность распознавания и чувствительность восприятия к эмоциям, сигнализирующим об угрозе.
( a ) Доля правильных ответов для (слева направо) предотвращение/гнев, прямое/гнев, предотвращение/страх и прямое/страх. УГРОЗА + комбинации взгляда и эмоций (выделены оранжевым цветом) были связаны с повышенной точностью распознавания. ( b ) Психометрическая функция, представляющая долю реакций «гнева» в зависимости от признаков гнева (процентная трансформация, 0 = нейтральная, отрицательная по отношению к страху и положительная по отношению к гневу) для УГРОЗА+ (оранжевый) и УГРОЗА- (зеленый) ) сочетания взгляда и эмоций. Точки и прикрепленные планки погрешностей указывают данные человека (среднее значение ± стандартная ошибка среднего). Линии и заштрихованные полосы погрешностей указывают прогнозы наиболее подходящей модели. ( c ) Параметр оценки наклона психометрической кривой (соответствующей эмоциональной чувствительности) для комбинаций УГРОЗА+ и УГРОЗА-. **р < 0,01, ***р < 0,001.
Кроме того, наблюдалась значимая эмоция по взаимодействию взгляда с силой эмоции (F 6,138 = 4,3, p<0,01), объясняемая более сильным влиянием взгляда на категоризацию эмоций при слабой силе эмоции (взгляд по взаимодействию эмоций для уровней с 1 по 4, Ф 1,23 = 23,8, p<0,001), чем при высокой силе эмоций (взгляд по взаимодействию эмоций для уровней 5-7, F 1,23 = 5,1, p<0,05). Анализ времени реакции (ВР) выявил снижение правильных ВР с силой эмоции (повторные измерения ANOVA, F 6,138 = 54,5, p<0,001), более быструю реакцию на сердитые лица по сравнению с испуганными лицами (F 1,23 = 12). , p<0,01), и более быстрая реакция на прямой взгляд по сравнению с отводом (F 1,23 = 7,7, p<0,05). Кроме того, наблюдалась эмоция при взаимодействии взгляда (F 1,23 = 8, p<0,01), что соответствует более быстрому времени реакции на прямой по сравнению с отведенным взглядом только в состоянии гнева (t 23 = -3,9, p<0,001).
Чтобы охарактеризовать механизм, лежащий в основе улучшенного распознавания эмоций, сигнализирующих об угрозе, мы подобрали поведение участников, используя семейство вложенных моделей выбора, которые предполагают, что решения формируются на основе зашумленного сравнения отображаемой эмоции и критерия, по следующей рецептуре (подробнее см. Материалы и методы):
P(гнев)=Φ[w·x+b]·(1-ε)+0,5·ε
где P(гнев) соответствует вероятности оценки лица как гневного, Ф[. ] к кумулятивной нормальной функции, w к перцептивной чувствительности к отображаемой эмоции, x к свидетельству (силе эмоции) в пользу гнева или страха в каждом испытании (от -7 для интенсивного выражения страха до + 7 для сильного выражения гнева), b к аддитивному независимому от стимула смещению в пользу одной из двух реакций/эмоций и ε к доле ошибок (случайных предположений) в испытаниях.
Мы сравнили «нулевую» модель, которая не учитывала контекстуальное влияние направления взгляда на процесс принятия решения, с двумя дополнительными моделями, реализующими два разных механизма, которые могут объяснить наблюдаемое повышение точности распознавания для комбинаций взгляда и эмоций «УГРОЗА+». : 1. первый вариант, в котором направление взгляда смещает критерий принятия решения в пользу эмоции, сигнализирующей о более высокой угрозе, и 2. второй вариант, в котором направление взгляда усиливает чувствительность к эмоциям, сигнализирующим о более высокой угрозе. Выбор байесовской модели показал, что повышение чувствительности для комбинаций УГРОЗА + значительно лучше объясняет поведенческие данные, чем изменение критерия (коэффициент Байеса ≈ 10 8 , вероятность превышения p вместо > 0,74). Оценки максимального правдоподобия параметра перцептивной чувствительности w , извлеченные из модели-победителя, были значительно повышены для комбинаций УГРОЗА+ взгляда и эмоций (9). 0141 т 23 = 3,9, р<0,001; Рисунок 3б,в). Доля срывов не различалась между комбинациями УГРОЗА+ и УГРОЗА- (t 23 = 0,4, p>0,5).
Расширенное нейронное кодирование эмоций, сигнализирующих об угрозе
Чтобы подтвердить обнаружение повышенной чувствительности к эмоциям, сигнализирующим об угрозе, и идентифицировать ее нейронные субстраты, мы затем исследовали, как выражения лица модулируют ЭЭГ-активность скальпа, зарегистрированную во время задачи категоризации эмоций. Вместо вычисления средних значений, связанных с событием, мы полагались на подход, основанный на параметрической регрессии, заключающийся в регрессии сигналов ЭЭГ в одной попытке в зависимости от силы отображаемой эмоции на каждом электроде и в момент времени после предъявления лица (Wyart et al., 2012а, 2015). Общая линейная регрессионная модель (GLM) была адаптирована к данным ЭЭГ, где сила эмоции (от 0 для нейтрального/безэмоционального выражения до 7 для интенсивного выражения страха/гнева) была введена в качестве предиктора широкополосных сигналов ЭЭГ от одного испытания к другому. на каждом электроде и в момент времени после начала стимула (от 0,2 с до 1,0 с после начала стимула). Полученный временной ход на каждом электроде представляет собой степень, в которой активность ЭЭГ «кодирует» (ковариируется с) силу эмоции, обеспечиваемую измененными чертами лица.
Оценки параметров наклона регрессии выявили значимые корреляции между силой эмоции и активностью ЭЭГ, пик которой начинается примерно через 280 мс после предъявления лица в височной (t-тест против нуля, t 23 = -12,7, p<0,001) и лобной электродов ( t 23 = 8,7, p<0,001), а затем около 500 мс и по времени отклика на центрально-теменных ( t 23 = 10,2, p<0,001) и лобных электродах ( t 23 = -7,9, p<0,001) (рис. 4a–c). Временные точки и электроды, где оценки параметров значительно отличаются от нуля, указывают на нейронное кодирование информации об эмоциях. Сила этого нейронного кодирования, индексированная по амплитуде оценки параметра, обеспечивает меру чувствительности нейронов к информации об эмоциях.
Усовершенствованное нейронное кодирование эмоций, сигнализирующих об угрозе.
( a ) Средняя панель: топография скальпа нейронного кодирования на 280 мс, соответствующая его первому пику кодирования силы эмоции, усредненной по условиям (пик отклонения от нуля) и выраженной как средние оценки параметров в произвольных единицах ( а.е.). Точки указывают интересующие электроды, где нейронное кодирование было максимальным. Левая и правая панели: кодирование временной динамики состояний УГРОЗА+ и УГРОЗА- на интересующих электродах. Заштрихованные столбцы погрешностей указывают на sem. Толстые оранжевые и зеленые линии указывают значимость по сравнению с нулем при кластерно-скорректированном p-значении 0,05. Заштрихованные серые области указывают на значительные различия между условиями УГРОЗА+ и УГРОЗА- при p <0,05. ( b ) Те же соглашения, что и ( a ) для второго пика нейронного кодирования на 500 мс. ( c ) Те же соглашения, что и ( a ) для третьего пика нейронного кодирования во время отклика. ( d ) Предполагаемые корковые источники кодирования разницы между условиями УГРОЗА+ и УГРОЗА- во время значительной разницы между условиями на 170 мс. ( e ) То же, что ( d ) при 500 мс. ( f ) То же, что и ( d ) во время отклика. FG: веретеновидная извилина, pSTS: задняя верхняя височная борозда, SMG: надмаргинальная извилина, ANG: угловая извилина, STG: верхняя височная извилина, MTG: средняя височная извилина, OCG: затылочная извилина, aINS: передняя часть островка, IFS: нижняя лобная борозда, TP: височный полюс, OFC: орбитофронтальная кора, OP: затылочный полюс, TPJ: височно-теменное соединение, dlPFC: дорсолатеральная префронтальная кора.
Чтобы проверить нейронную сигнатуру повышенной чувствительности к эмоциям, сигнализирующим об угрозе, мы сравнили оценки параметров, извлеченные отдельно для УГРОЗА+ (прямой гнев и предотвращенный страх) и УГРОЗА- (предотвращенный гнев и прямой страх) комбинаций взгляда и эмоций. Этот контраст выявил повышенные оценки параметров для комбинаций УГРОЗА+ сначала на 170 мс во времени (парный t-критерий, t 23 = -2,5, p<0,05) и лобные электроды ( t 23 = 2,2, p<0,05), а затем через 500 мс и время отклика на центрально-теменных ( t 23 = 2,2, p<0,05) и фронтальные электроды ( t 23 = -2,4, p<0,05) (рис. 4a–c). Это открытие указывает на то, что нейронное усиление кодирования эмоций было усилено в эти моменты времени и электроды для эмоций, сигнализирующих об угрозе. Это улучшение, зависящее от угрозы, оставалось значительным при рассмотрении только правильных ответов (временно: т 23 = -2,1, р<0,05; центрально-теменной т 23 = 4,2, р<0,001). Интересно, что комбинации УГРОЗА+ не были связаны с повышенными средними значениями, связанными с событиями, при классической пиковой латентности (P1, N170, P2, P3: все t 23 <1,95, p>0,07). Чтобы оценить, какие области мозга генерируют сигналы ЭЭГ, записанные на скальпе, мы вычислили корковые источники этого усиленного кодирования эмоций, сигнализирующих об угрозе, применив тот же регрессионный подход к оценкам тока минимальной нормы, распределенным по поверхности коры. Оценки параметров в интересующие моменты времени (где наблюдались различия между комбинациями УГРОЗА+ и УГРОЗА-) затем сравнивались между двумя условиями (см. Материалы и методы). Увеличение наклона регрессии для комбинаций УГРОЗА+ сместилось от вентральных зрительных областей, селективных к выражению эмоций на лице (веретенообразная извилина и верхняя височная борозда) около 170 мс, к ассоциативным областям мозга, охватывающим теменную, височную и лобную кору (верхняя и средняя височная, височный полюс, и орбитофронтальная кора) через 500 мс, а затем в сенсомоторные области вокруг начала ответа (дорсальная центральная, теменная и лобная области) (рис. 4d–f).
Эти нейронные эффекты сходятся с поведенческим моделированием в пользу устойчивого повышения перцептивной чувствительности к эмоциям, сигнализирующим об угрозе, начиная со 170 мс после предъявления лица и продолжаясь до начала реакции. Дополнительные данные подтверждают нашу гипотезу о том, что усиление нейронной чувствительности к комбинациям УГРОЗА+ конкретно связано с увеличением подразумеваемой угрозы для этих комбинаций взгляда и эмоций. Отдельная группа участников оценила идентичности, использованные в задаче категоризации эмоций, с точки зрения воспринимаемой угрозы и надежности (см. Материалы и методы), а оценки на уровне группы для каждой идентичности были регрессированы по сравнению с сигналами ЭЭГ, полученными в одной попытке, в качестве дополнительных регрессоров. Эта регрессия показала, что воспринимаемая угроза, но не достоверность, значительно коррелировала с височной и центрально-теменной активностью ЭЭГ через 500 мс после предъявления лица в том же направлении, что и контраст между комбинациями УГРОЗА+ и УГРОЗА- (угроза: т 23 > 3,6, р<0,01; достоверность: t 23 < 0,7, p>0,48).
Независимое от внимания улучшение нейронной обработки с помощью угрозы
Анализ нейронных данных до сих пор подтвердил гипотезу о том, что контекстуальная информация о взгляде влияет на категоризацию эмоций, увеличивая перцептивную чувствительность к эмоциям, сигнализирующим об угрозе. Такой эффект может быть опосредован повышенным нисходящим вниманием к эмоциям, сигнализирующим об угрозе, т. Е. Комбинациям УГРОЗА + (прямой гнев и предотвращенный страх). Чтобы проверить эту возможность, мы исследовали, влияют ли остаточные флуктуации в сигналах ЭЭГ в одиночной попытке, необъяснимые изменениями в силе эмоций (измеренными с помощью предыдущих регрессий), на точность последующего категорического решения, т. е. перцептивную чувствительность к отображаемой эмоции. Этот подход напоминает меры «вероятности выбора», применяемые в электрофизиологии для измерения корреляции между нейронной активностью и поведением выбора (Britten et al., 19).96; Шадлен и др., 1996; Parker and Newsome, 1998) – путем оценки того, насколько флуктуации записанных нейронных сигналов «считываются» последующим решением (Wyart et al., 2012a, 2015). Независимые от стимулов улучшения корреляций нейронного выбора классически интерпретируются как увеличение «считываемых» весов, то есть повышенное нисходящее внимание к этим нейронным сигналам (Nienborg and Cumming, 2009, 2010). Здесь повышенная нервная модуляция выбора для условий УГРОЗА+ может указывать на увеличение нисходящего внимания к эмоциям, сигнализирующим об угрозе, что, в свою очередь, может объяснить наблюдаемое увеличение перцептивной и нервной чувствительности к этим комбинациям взгляда и эмоций.
Чтобы проверить эту гипотезу, мы ввели остатки ЭЭГ из предыдущей регрессии в зависимости от силы эмоций в качестве дополнительного предпочтительного предиктора «посредничества» — как средства проверки того, меняются ли эти нейронные сигналы совместно с перцептивной чувствительностью (подробности см. в разделе «Материалы и методы»). . На практике мы оценили параметры b mod и w mod этих терминов нейронной модуляции в каждый момент времени после представления лица с помощью ЭЭГ-информированной регрессии выбора, для которой нейронные остатки от пробы к пробе e из регрессии против силы эмоции вводились либо отдельно (аддитивное влияние, параметр b mod ), либо как их взаимодействие с силой эмоции (мультипликативное влияние, параметр w mod ) в качестве дополнительных предикторов последующего выбор:
P(гнев)=ϕ[(w+wmod· e) x+b+bmod·e]
Временной ход и пространственное распределение этой нервной модуляции перцептивной чувствительности ( wmod ) качественно соответствовали нейронной кодирование силы эмоции (рис. 5a–c) с отрицательным временным компонентом, достигающим максимума при 270 мс ( t 23 = -4,2, p<0,001), затем положительный центрально-теменной с пиком около 600 мс ( t 23 = 8,0, p<0,001) и затем во время ответа ( t 23 = 7,6, р<0,001). Мы использовали выбор байесовской модели, чтобы подтвердить, что остаточные значения ЭЭГ мультипликативно коррелируют с перцептивной чувствительностью ( wmod ) последующего решения, а не аддитивно как систематическая ошибка ( bmod ) в силе эмоций, как во времени (фактор Байеса ≈ 10 3,4 , p exc = 0,79) и центрально-теменных электродов (коэффициент Байеса ≈ 10 8,9 , p exc = 0,99). Важно отметить, что не наблюдалось разницы в силе модуляции между комбинациями УГРОЗА+ (прямой гнев и предотвращенный страх) и УГРОЗА- (предотвращенный гнев и прямой страх) (височные: t 23 = -0,4, p>0,5; центрально-теменные: т 23 = 0,1, р>0,5). Чтобы определить, связано ли это отсутствие значимой разницы с подлинным отсутствием эффекта (а не с отсутствием статистической чувствительности), мы рассчитали коэффициенты Байеса при тех же параметрических предположениях, что и обычная статистика (см. Материалы и методы). Мы получили коэффициенты Байеса ниже 10–4 на височных и центрально-теменных электродах, что свидетельствует об отсутствии увеличения «считываемых» весов для состояний «УГРОЗА+». Этот нулевой эффект предполагает, что наблюдаемое усиление перцептивной и нервной чувствительности к этим сигнализирующим об угрозе комбинациям взгляда и эмоций не вызывается косвенно усилением нисходящего внимания в этих условиях.
Отсутствие зависящего от угрозы усиления корреляций нейронного выбора.
( a ) Средняя панель: топография скальпа корреляций нейронного выбора, выраженная как модуляция перцептивной чувствительности с помощью остатков кодирования ЭЭГ на 280 мс, тот же момент времени, показанный на рисунке 4a. Интересующие электроды, указанные точками, такие же, как на рисунке 4а. Левая и правая панели, временная динамика модуляции перцептивной чувствительности остатками кодирования ЭЭГ, выраженными в условных единицах (а.е.). Те же соглашения, что и на рисунке 4a. ( b ) Те же соглашения, что и ( a ) при 500 мс. ( c ) Те же соглашения, что и ( a ) во время отклика. Изменение силы модуляции во времени согласуется с изменением оценки параметра кодирования. Никакой разницы между УГРОЗА+ и УГРОЗА- не наблюдается.
Раннее нейронное кодирование эмоций, сигнализирующих об угрозе, при двигательной подготовке
Мы пришли к выводу, что угроза может влиять не только на нейронную репрезентацию отображаемой эмоции в зрительной и ассоциативной коре, но и на подготовку предстоящего ответа в эффекторно-селективных структурах (Conty et al. , 2012). Для измерения сигналов подготовки к ответу в нейронных данных мы рассчитали спектральную мощность в мю- и бета-диапазонах частот (8–32 Гц) (Donner et al., 2009; de Lange et al., 2013). Выполнение и подготовка движений конечностей совпадают с подавлением низкочастотной (8–32 Гц) активности, более выраженной в моторной коре, контралатеральной по сравнению с ипсилатеральной по отношению к движению. Таким образом, ожидается, что вычитание контралатеральной активности из ипсилатеральной моторной коры приведет к положительному показателю моторной подготовки. Контраст между ответами левшей и правшей во время ответа выявил латеральные центральные электроды, связанные с фокальными источниками в моторной коре (рис. 6а). Вычитание контралатеральных сигналов из ипсилатеральных относительно руки, назначенной для реакции «страха» (уравновешенной между участниками), дало индекс моторной латерализации, знак которого значительно предсказывает предстоящий выбор (гнев или страх) за 360 мс до начала реакции (парный t-критерий, т 23 = 4,6, р<0,001; Рисунок 6б).
Кодирование эмоций, сигнализирующих об угрозе, в мерах латерализации двигательных реакций.
( a ) Верхняя панель, топография скальпа до ответа мощности частоты времени в диапазоне 8–32 Гц за последние 100 мс до ответа, для испытаний, в которых испытуемые реагировали левой рукой, за вычетом испытаний, в которых они реагировали их правая рука. Точки соответствуют выбранным электродам, где эффект был максимальным. Нижняя панель: соответствующие нейронные источники. ( b ) Динамика латерализации ответа во времени (мощность частотно-временной активности от контралатеральных электродов минус ипсилатеральные электроды к руке, используемой для ответа на «страх») в сторону гнева и страха, когда выбор был между гневом (красный) или страхом (синий). Заштрихованные столбцы погрешностей указывают на sem. Заштрихованная серая область указывает на значительную разницу в моторной латерализации между реакциями гнева и страха. ( c ) Кодирование силы эмоции в индексе латерализации реакции для состояний УГРОЗА+ (оранжевый) и УГРОЗА- (зеленый). Различия между условиями наблюдаются через 200 мс после начала стимула (стимул заблокирован, верхняя панель) и во время ответа (заблокирован ответ, нижняя панель). Условные обозначения те же, что и на рис. 4. ( d ) Динамика корреляций нейронного выбора во времени, выраженная как модуляция аддитивного смещения за счет моторной латерализации, кодирующей остатки в произвольных единицах (а.е.) с блокировкой стимула (верхняя панель) и блокировкой реакции (нижняя панель). Условные обозначения те же, что и на рисунке 4.
Мы применили предыдущий подход нейронного кодирования, регрессировав этот индекс моторной латерализации по сравнению с подписал силы эмоции (от 0 для нейтрального выражения до ± 7 для сильного выражения гнева/страха) на основе испытания за испытанием. Оценки параметров наклона регрессии значительно отклонялись от нуля через 400 мс после начала стимула (t-тест против нуля, t 23 = 5,1, p<0,001) и во время ответа ( t 23 = 5,2, p<0,001) – отражает более сильную подготовку к реакции на более сильные (т.е. более диагностические) эмоции. Вычисление наклонов регрессии отдельно для комбинаций УГРОЗА+ (прямой гнев и предотвращенный страх) и УГРОЗА- (предотвращенный гнев и прямой страх) показало, что комбинации УГРОЗА+ вызывают более сильное кодирование силы эмоции при двигательной подготовке в конце начала реакции (9).0141 t 23 = 2,9, p<0,01), но также рано, примерно через 200 мс после предъявления лица ( t 23 = 3,2, p<0,01). Это раннее двигательное усиление, зависящее от угрозы, оставалось значимым при рассмотрении только правильных ответов ( t 23 = 3,0, p<0,01). В то время как УГРОЗА- комбинации взгляда и эмоций не были связаны со значительным нейронным кодированием при двигательной подготовке до 440 мс после предъявления лица ( t 23 <0,8, p>0,4), комбинации УГРОЗА+ приводили к значимому нейронному кодированию между 100 и 320 мс с пиком при 200 мс ( t 23 = 3,2, p<0,01; рис. 6c).
Чтобы определить, влияет ли это раннее нейронное кодирование эмоций, сигнализирующих об угрозе, при двигательной подготовке на скорость последующих ответов, мы пересчитали и сравнили параметры регрессии, оцененные отдельно для быстрых и медленных ответов на комбинации УГРОЗА+ (прямой гнев и предотвращенный страх), на на основе медианного разделения времени отклика, основанного на силе эмоций. Это сравнение выявило одиночное постепенное нейронное кодирование силы эмоции при двигательной подготовке, предшествующей быстрым, но не медленным реакциям, возникающее уже через 150 мс (при пороге p -значение 0,05) после предъявления лица (разница в начале кодирования между быстрой и медленной реакцией, складной нож (Kiesel et al., 2008, см. Материалы и методы) t 23 = 5,2, p< 0,001; рис. 7а). Этот эффект указывает на то, что раннее нейронное кодирование комбинаций УГРОЗА+ при двигательной подготовке характерно для эффективных (быстрых) ответов. Мы подтвердили, что этот латентный сдвиг в нейронном кодировании был избирательным в отношении сигналов двигательной подготовки, выполнив такое же сравнение нейронного кодирования силы эмоций на центрально-теменных электродах. Этот контраст выявил только разницу в пиковой амплитуде, а не в латентном периоде начала, между быстрыми и медленными ответами (пиковая амплитуда: т 23 = 5,1, р<0,001; латентность начала: складной нож t 23 = -1,3, p>0,2; Рисунок 7б).
Кодирование силы эмоции как функции времени реакции (ВР) в двигательных и теменных структурах.
( a ) Нейронное кодирование силы эмоции для условий УГРОЗА+ при моторной латерализации для быстрого и медленного времени реакции (RT): когда RT были быстрыми, кодирование силы эмоции становилось значимым через 150 мс и постепенно возрастало до ответа; напротив, когда RT были медленными, кодирование силы эмоций стало значимым позже, на 540 мс. Заштрихованные столбцы погрешностей указывают на sem. Толстые темно- и светло-серые линии указывают значимость по сравнению с нулем при кластерно-скорректированном p-значении 0,05. Заштрихованные серые полосы указывают на значительные различия между быстрыми и медленными ответами. Задержка кодирования значительно различается между быстрыми и медленными RT, ***: p<0,001 ( b ) Кодирование силы эмоций в теменных электродах. Соглашение такое же, как ( a ). Быстрые ответы связаны с более сильным нейронным кодированием силы эмоций, но без каких-либо изменений в задержке кодирования.
Наконец, мы провели анализ корреляций нейронного выбора, чтобы оценить, влияет ли раннее нейронное кодирование эмоций, сигнализирующих об угрозе, при двигательной подготовке не только на скорость, но и на содержание (гнев или страх) последующих реакций. Описанный выше нейронный анализ «посредничества» в разных условиях показал, что независимые от стимула колебания индекса моторной латерализации ковариируются как смещение аддитивного выбора в предстоящем ответе через 400 мс после предъявления лица (9). 0141 т 23 = 2,9, р<0,01). Действительно, в отличие от флуктуаций височной и центрально-теменной активности, влияние вариабельности моторной латерализации на категоризацию эмоций лучше описывалось как аддитивная ошибка выбора , а не как изменение перцептивной чувствительности (коэффициент Байеса ≈ 10 36,4 , p exc = 0,98) – согласуется с его предполагаемой ролью в качестве моторного представления переменной решения (Donner et al., 2009).; де Ланге и др., 2013). Никакой разницы в силе модуляции не наблюдалось между комбинациями УГРОЗА+ (прямой гнев и предотвращенный страх) и УГРОЗА- (предотвращенный гнев и прямой страх) ( t 23 <1,6, p>0,1; рис. 6d). Критически важно, что даже при рассмотрении только комбинаций остаточная вариабельность моторной латерализации, измеренная между 100 и 320 мс (где нейронное кодирование эмоций, сигнализирующих об угрозе, было значительным), не оказала значительного влияния на предстоящий выбор (90–141 t 9). 0142 23 < 1,4, р>0,17). Этот нулевой эффект был подтвержден выбором байесовской модели, которая определила подлинное отсутствие корреляции нейронного выбора как наиболее вероятное объяснение данных (коэффициент Байеса ≈ 10 2,3 , p ex = 0,96). Это открытие указывает на то, что раннее нейронное кодирование эмоций, сигнализирующих об угрозе, при двигательной подготовке происходит раньше, чем формирование предстоящего выбора.
Зависимое от тревоги нейронное кодирование эмоций, сигнализирующих об угрозе
У населения в целом тревога классически связана с повышенной чувствительностью к сигналам угрозы в социальных условиях (Bishop, 2007; Cisler and Koster, 2010). Здесь мы оценили, зависит ли усиленная нейронная обработка эмоций, сигнализирующих об угрозе, в височных и двигательных областях с уровнем тревожности у наших участников. С этой целью мы измерили уровень тревожности в начале экспериментальной сессии, до сбора данных, используя Опросник тревожных состояний Спилбергера (STAI) (Spielberger et al. 19).83). Этот вопросник для самооценки позволяет определить степень уязвимости к тревожным расстройствам (Grupe et al., 2013). Состояние тревожности участников варьировалось от 20 до 45 (среднее значение = 30,5, стандартное отклонение = 6,8). Показатели личностной тревожности варьировались от 22 до 52 (среднее значение = 38,2, стандартное отклонение = 7,8). Эти оценки сопоставимы с исходными опубликованными нормами для этой возрастной группы (Spielberger, 1983) и с французскими нормативными данными (Bruchon-Schweitzer and Paulhan, 1993). Мы проанализировали влияние тревожности на поведенческие и нейронные данные двумя взаимодополняющими способами: 1. путем разделения участников на две группы одинакового размера на основе их измеренной тревожности и 2. путем сопоставления параметров нейронного кодирования, оцененных на уровне отдельных участников. с их размеренной тревогой. Удивительно, но мы не обнаружили влияния тревожности на общие показатели производительности (9).0141 t 11 < 0,05, p>0,9), ни о разнице между УГРОЗОЙ+ (прямой гнев и предотвращенный страх) и УГРОЗОЙ- (отклоненный гнев и прямой страх) сочетаниями взгляда и эмоций ( F 1,22 < 0,4, р>0,5).
Тем не менее отсутствие эффекта тревоги на поведенческом уровне сопровождалось компенсаторной двойной диссоциацией в нейронных данных. Действительно, ситуационная тревожность значительно влияла на нейронное кодирование силы эмоций на височных электродах на пике нейронного кодирования через 280 мс после предъявления лица (среднее разделение, взаимодействие: F 1,22 = 7,3, р=0,01; Рисунок 8a): наблюдатели с высоким уровнем тревожности не показали различий в нейронном кодировании между комбинациями УГРОЗА+ и УГРОЗА- (УГРОЗА+: t 11 = -5,8, p<0,001; УГРОЗА-: t 11 = -6,1, p<0,001, разница: t 22 = 0,84, p = 0,4), тогда как малотревожные наблюдатели кодировали исключительно УГРОЗУ+ при той же латентности (УГРОЗА+: t 11 = -6,5, p<0,001; УГРОЗА-: t 11 = — 1,8, р=0,08, разница: t 22 = -3,0, р=0,01). Параметрическая оценка взаимосвязи между ситуационной тревожностью и разницей в нейронном кодировании комбинаций УГРОЗА+ и УГРОЗА- оказалась значимой (коэффициент корреляции Пирсона r = 0,51, d. f. = 22, p = 0,01; рис. 8а). Другими словами, высокая тревожность была связана со значительным и индифферентным нейронным кодированием негативных эмоций, сигнализирующих об угрозе или нет, в вентральных избирательных областях лица.
Модуляция кодирования угрозы индивидуальной тревогой.
( a ) Левая панель: корреляция (Пирсон) между состоянием тревожности и разностью оценок параметров кодирования между состояниями УГРОЗА+ и УГРОЗА- в височных электродах на 280 мс. Правая панель: оценки параметров кодирования в височных электродах, разделенные на людей с высокой и низкой тревожностью для условий УГРОЗА+ и УГРОЗА- на 280 мс. T+: УГРОЗА+, T-: УГРОЗА-. ( b ) Слева, корреляция (Пирсон) между тревогой состояния и оценками параметров кодирования в сигналах моторной латерализации для состояния УГРОЗА+ на 200 мс. Справа: оценки параметров кодирования в сигналах моторной латерализации разделены на людей с высокой и низкой тревожностью для условий УГРОЗА+ и УГРОЗА- через 200 мс. ***: р<0,001, *р<0,05.
Интересно, что в раннем временном окне (пик кодирования на 200 мс), когда в моторных сигналах кодировались только комбинации УГРОЗА+ (прямой гнев и предотвращенный страх), наблюдалась обратная картина: только у лиц с высокой тревожностью на этом этапе наблюдалось значительное кодирование. латентность (взаимодействие межсубъектной тревожности и парного взгляда F 1,22 = 4, р=0,05; рис. 8б). Чем больше люди были тревожны, тем больше они кодировали релевантные для наблюдателя сигналы угрозы в двигательных системах (корреляция между оценками параметров для условий УГРОЗА+ и коэффициентом тревожности состояния Пирсона r = 0,52, df = 22, p <0,01; Рисунок 8b). Более того, нейронное кодирование эмоций УГРОЗА+ в двигательных сигналах коррелировало с поведенческой чувствительностью к эмоциям УГРОЗА+ у высокотревожных лиц (коэффициент корреляции Пирсона r = 0,66, d. f. = 10, p=0,01), в то время как у низкотревожных людей этого не наблюдалось. (коэффициент корреляции Пирсона r = -0,42, df = 10, p>0,16, разница между коэффициентами, p<0,01). Подводя итог, можно сказать, что в то время как люди с высокой тревожностью одинаково обрабатывают все сигналы угрозы в избирательных областях лица, они избирательно кодируют сигналы угрозы, которые имеют отношение к ним, в двигательных системах, и это кодирование отражает их поведенческую чувствительность к эмоциям, сигнализирующим об угрозе.
Обсуждение
Точное декодирование эмоций у других, особенно негативных, передает адаптивные преимущества в социальной среде. Хотя типичные социальные взаимодействия не требуют явной категоризации эмоций, выражаемых другими, точное понимание нейронных механизмов, участвующих в распознавании эмоций, дает важную информацию о том, как человеческий мозг обрабатывает социально значимые сигналы. И хотя предыдущая работа выявила нейронные корреляты перцептивных решений (Gold and Shadlen, 2007; Heekeren et al. , 2008), только в нескольких исследованиях рассматривался вопрос о том, как такие решения формируются на основе социально значимых стимулов, таких как выражение лица. проявления эмоций. Как и в большинстве задач перцептивной категоризации, мы манипулировали неоднозначностью сенсорных данных — здесь, используя контролируемые морфы между гневными или испуганными выражениями и нейтральными. Но благодаря социальной природе наших стимулов мы могли бы одновременно и неявно манипулировать контекстуальной значимостью отображаемой эмоции с точки зрения подразумеваемой угрозы для наблюдателя, используя направление взгляда, и применять подход, управляемый моделью, для характеристики нейронной приоритизации угрозы. -передача информации в электрических сигналах мозга.
Направление взгляда, которое действует как контекстуальная подсказка в нашей задаче категоризации эмоций, отличается от контекстуальных подсказок, обнаруженных в перцептивных исследованиях принятия решений, которые обычно предоставляются за сотни миллисекунд до стимула, релевантного для принятия решения (Rahnev et al. , 2011; Kok et al., 2012; Wyart et al., 2012b; de Lange et al., 2013). Здесь, как и во многих социальных ситуациях, контекстуальные сигналы могут возникать одновременно со стимулом, релевантным для принятия решения, — свойство, которое сильно ограничивает их влияние на обработку стимула. Более того, значение контекстуальных сигналов (например, сигналов внимания или ожидания), используемых в перцептивных исследованиях принятия решений, обычно указывается явно и, таким образом, явно обрабатывается участниками во время выполнения задачи (Kok et al., 2012; Wyart et al., 2012б). Здесь, напротив, направление взгляда не имеет значения для задачи категоризации эмоций и, следовательно, не нуждается в явной обработке. Несмотря на эти два отличия от других контекстных сигналов, мы показываем, что направление взгляда настраивает нейронную обработку информации об эмоциях с 200 мс после появления стимула до реакции в сенсорных, ассоциативных и двигательных цепях человеческого мозга.
Предыдущие наблюдения повышения субъективных оценок и улучшения распознавания выражений гнева в сочетании с прямым взглядом и выражений страха в сочетании с отведенным взглядом интерпретировались с точки зрения контекстуальной оценки отображаемой эмоции во время ее обработки (Adams and Kleck, 2003; Сандер и др. , 2007; Адамс и др., 2012). В частности, теории «оценки» (Sander et al., 2007) подчеркивают, что сердитое выражение лица в сочетании с прямым взглядом может быть интерпретировано как поведенчески «уместное» для наблюдателя как объект словесного или физического нападения, в то время как выражение страха взгляд в сторону от наблюдателя может сигнализировать об источнике опасности в непосредственной близости от наблюдателя. Однако механизмы, реализующие предложенную контекстуальную оценку эмоций в зависимости от их подразумеваемой угрозы для наблюдателя, остались неясными. Направление взгляда может либо сместить воспринимаемую эмоцию в сторону ее наиболее релевантной (сигнализирующей об угрозе) интерпретации, т. е. гнева в сочетании с прямым взглядом или страха в сочетании с отведенным взглядом, либо повысить чувствительность к наиболее релевантной эмоции. Настоящее исследование дает прямой ответ на этот вопрос, показывая как поведенчески (путем сравнения количественных соответствий двух эффектов с поведенческими данными), так и нейронно (путем регрессии сигналов мозга в зависимости от силы эмоций), что улучшенная точность распознавания эмоций, сигнализирующих об угрозе, соответствует избирательное нейронное усиление перцептивной чувствительности к этим комбинациям взгляда и эмоций.
Сигналы ЭЭГ, модулированные информацией об эмоциях, на центрально-теменных электродах от 500 мс после предъявления лица до выполнения ответа, результат в соответствии с «супрамодальным» признаком перцептивной интеграции, о котором сообщалось в предыдущих исследованиях (O’Connell et al., 2012; Wyart и др., 2012а). Было предложено, чтобы эта центрально-теменная позитивность кодировала переменную решения «общего домена», поскольку она варьируется в зависимости от силы сенсорных свидетельств как для зрительных, так и для слуховых решений, независимо от связанной реакции (O’Connell et al., 2012) . Здесь было обнаружено, что та же центрально-теменная позитивность увеличивается с силой эмоций выражений лица, что указывает на переменную решения в нашей задаче категоризации эмоций. Важно отметить, что сила этих отношений была усилена эмоциями, сигнализирующими об угрозе. Это улучшенное нейронное представление угрожающих комбинаций взгляда и эмоций не может быть объяснено повышенным вниманием или реакциями удивления, поскольку ранее сообщалось, что центрально-теменной потенциал «P3» изменяется в зависимости от ресурсов внимания (Johnson, 19). 88) и удивление (Mars et al., 2008), не усиливались в ответ на эмоции, сигнализирующие об угрозе. Более того, мы также могли бы исключить возможность того, что это усиленное нейронное кодирование косвенно вызвано усилением избирательного внимания, которое должно было быть связано с улучшенным «расшифровыванием» решений участников из лежащих в их основе нейронных сигналов (Nienborg and Cumming, 2009). , 2010; Уайарт и др., 2015). Таким образом, мы предполагаем, что усиленная нейронная обработка сигнализирующих об угрозе эмоций происходит независимо от внимания и восходит снизу вверх.
Более ранние контекстуальные модуляции обработки эмоций также наблюдались в вентральных областях, селективных к лицу, через 170 мс после предъявления лица. Хотя эти результаты противоречат «двухэтапному» взгляду, согласно которому информация об эмоциях и взгляде будет обрабатываться независимо в течение первых сотен миллисекунд (Pourtois et al., 2010), прежде чем интегрироваться в зависимости от их значимости для наблюдателя (Клучарев и Sams, 2004), они согласуются с недавними выводами (Conty et al. , 2012; El Zein et al., 2015) о ранних взаимодействиях между эмоциями и информацией взгляда на компонентах N170 и P200. В эти ранние латентные периоды только эмоции, сигнализирующие об угрозе, кодировались селективными по лицу нейронными сигналами, отражая более быструю обработку эмоций, сигнализирующих о непосредственной угрозе наблюдателю, как функцию связанного с ними взгляда.
Что еще более поразительно, направление взгляда также модулировало кодирование эмоциональных выражений в эффекторно-селективных областях параллельно с эффектами, наблюдаемыми в вентральных избирательно-лицевых областях: только эмоции, сигнализирующие об угрозе, кодировались в сигналах подготовки к ответу, лежащих над моторной корой человека при 200 мс после презентации лица. Недавняя работа проливает свет на адаптивную функцию этого раннего представления сигналов угрозы в моторной коре. Нарушение этой двигательной репрезентации с помощью ТМС ухудшает распознавание на лице негативных (то есть потенциально угрожающих) эмоций, а не позитивных (Балкони и Бортолотти, 2012; 2013). Более того, восприятие природных сцен задействует моторную кору в очень ранние латентные периоды только тогда, когда эмоциональная валентность сцены отрицательна (Borgomaneri et al., 2014). В совокупности эти результаты подтверждают тесную связь между эмоциями и двигательными цепями (Grèzes et al., 2014), что позволяет мозгу быстро и эффективно реагировать на сигналы угрозы (Ohman and Mineka, 2001; Frijda, 2009).). Наши результаты основываются на этих более ранних наблюдениях, показывая, что мозг параметрически кодирует силу сигналов угрозы в моторной коре параллельно с их представлением в сенсорных областях, избирательных по отношению к лицу.
Наконец, наши данные показывают четкую функциональную диссоциацию между лицевыми и эффекторно-селективными областями в зависимости от индивидуальной тревожности. Повышенная чувствительность к эмоциям, сигнализирующим об угрозе, в лицевой избирательной височной коре определяется наблюдателями с низким уровнем тревожности, тогда как раннее усиление, измеренное в моторной коре, обнаруживается только у наблюдателей с высокой тревожностью. Наблюдение, что люди с высоким уровнем тревожности кодируют все негативные эмоции как одинаково (и сильно) выраженные в участках, выбранных для лиц, согласуется с более ранними сообщениями о «сверхбдительности» к потенциально угрожающим сигналам у этих людей (Bishop, 2007; Cisler and Koster). , 2010), и с их тенденцией интерпретировать неоднозначные стимулы как угрожающие (Beck et al., 1985) – оба связаны с гиперактивностью миндалины (Bishop, 2007; Etkin and Wager, 2007). Тем не менее наши результаты показывают, что сильно тревожные люди способны избирательно кодировать сигналы угрозы в моторной коре. В соответствии с идеей компенсаторного механизма, отчетливое нейронное усиление временной и двигательной активности, обнаруживаемое у людей с низким и высоким уровнем тревожности, приводит к сходным поведенческим улучшениям с точки зрения перцептивной чувствительности к сигналам угрозы. Вместе эта картина результатов предполагает, что тревога увеличивает относительный вклад двигательного пути во время обработки негативных социальных сигналов в соответствии с адаптивной функцией тревоги в эффективном обнаружении и быстром реагировании на угрозы в окружающей среде (Bateson et al. , 2011). Стоит отметить, что в настоящем исследовании участвовали только участники с показателями тревожности в диапазоне показателей здорового взрослого населения (Спилбергер, 19 лет).83), оставляя открытым вопрос о том, будут ли люди с клинической тревогой аналогичным образом задействовать свою моторную кору в ответ на угрожающие социальные стимулы. Кроме того, дальнейшие исследования должны оценить специфичность этих зависимых от тревоги эффектов в свете растущего количества доказательств в пользу коморбидности между тревогой и депрессивными расстройствами.
Применяя теоретические модели принятия решений к социально значимым стимулам, мы смогли охарактеризовать нейронные и вычислительные механизмы, лежащие в основе интеграции и интерпретации лицевых сигналов в имплицитном контексте угрозы. Эволюционное давление, возможно, заставило человеческий мозг расставлять приоритеты сигналов угрозы параллельно в сенсорной и моторной системах (Darwin, 1872; LeDoux, 2012). Такая расстановка приоритетов, которая, как выяснилось, происходит быстро, избирательно, но независимо от внимания, может повысить чувствительность восприятия к другим характеристикам сенсорной среды (Phelps et al. , 2006), чтобы обеспечить быстрые и адаптивные реакции в сложных, многомерных ситуациях опасности. .
Материалы и методы
субъектов
В эксперименте по ЭЭГ приняли участие 24 здоровых человека (12 женщин, средний возраст 22,7 ± 0,7 года). Все участники были правшами, с нормальным зрением и не имели неврологического или психиатрического анамнеза. Они предоставили письменное информированное согласие в соответствии с институциональными рекомендациями местного комитета по этике исследований (Хельсинкская декларация) и получили оплату за свое участие.
Стимулы
Посмотреть подробный протокол Стимулы состояли из 36 идентичностей (18 женщин), адаптированных из базы данных Radboud Faces Database (Langner et al., 2010), которые различались по эмоциям (нейтральные, сердитые или испуганные выражения) и направлению взгляда (прямо к участнику или отведенному на 45° к участнику). влево или вправо). Используя Adobe Photoshop CS5.1 (Adobe Systems, Сан-Хосе, Калифорния), лица были изменены, чтобы удалить все видимые волосы, изменить размер и положение, чтобы глаза, нос и рот оказались в пределах одной окружности. Все изображения были преобразованы в оттенки серого и обрезаны в овал размером 280 x 406 пикселей с центром в пределах кадра размером 628 x 429 пикселей.пиксельный черный прямоугольник.
Чтобы изменить интенсивность эмоционального выражения, лица были изменены с нейтрального на гневное выражение и с нейтрального на испуганное выражение с помощью FantaMorph (Abrosoft http://www.fantamorph.com/). Сначала мы создали 7 уровней морфов от нейтральных выражений к гневным и от нейтральных к испуганным выражениям (отдельно для стимулов прямого и отведенного взгляда) с помощью простого линейного преобразования морфинга. В результате получилось 30 условий для каждой идентичности: 7 уровней морфов * 2 эмоции * 2 направления взгляда = 28 и 2 нейтральных стимула с прямым и отведенным взглядом. Затем мы откалибровали переход между выражениями гнева и страха, выполнив предварительный тест оценки интенсивности эмоциональных выражений и скорректировав преобразования на основе результатов. 19испытуемым (9 женщин, средний возраст 24,7 ± 0,9 года) демонстрировали выражение лица в течение 250 мс и оценивали воспринимаемую эмоциональную интенсивность по непрерывной шкале от «совсем не интенсивной» до «очень интенсивной» с помощью мышиного устройства (с максимум 3 секунды на ответ). Мы скорректировали различия между эмоциями, линеаризовав средние кривые оцениваемой интенсивности и создав соответствующие морфы, которые были проверены на 10 новых субъектах (4 женщины, средний возраст 24,1 ± 1,9). Подводя итог, можно сказать, что стимулы включают 36 идентичностей с состоянием отведенного взгляда. и состояние прямого взгляда, каждое из которых имеет 7 уровней гнева и 7 уровней страха, уравновешенных по воспринимаемой эмоциональной интенсивности, и нейтральное состояние, что дает в общей сложности 1080 элементов (примеры стимулов см. на рис. 2а).
Экспериментальная процедура
Запросить подробный протокол Используя набор инструментов Psychophysics-3 (Brainard, 1997; Pelli, 1997), стимулы проецировались на черный экран. Каждое испытание начиналось с белого овала, ограничивающего лица, который сохранялся на протяжении всего испытания. Белый овал появился примерно на 500 мс, после чего появилась белая точка фиксации на уровне глаз примерно на 1000 мс (чтобы сохранить естественную фиксацию на приближающихся лицах и избежать движения глаз от центра овала к областям глаз). , затем стимулы появлялись в течение 250 мс. Задача участников состояла в том, чтобы решить, выражали ли лица Гнев или Страх, нажимая одну из двух кнопок, локализованных на двух внешних устройствах, которые держат в правой и левой руках, правым или левым указательным пальцем соответственно (рис. 2б). Использовалась карта «Гнев/Страх» (например, «Гнев: левая рука», «Страх: правая рука»), сохраняемая постоянной для каждого субъекта, уравновешенная по всем субъектам. Все стимулы предъявлялись однократно, всего было проведено 1080 проб. Эксперимент был разделен на 9экспериментальные блоки, каждый из которых состоит из 120 испытаний, сбалансированных по количеству эмоций, направлениям взгляда, полу и уровням морфов. После каждого блока участникам показывали процент правильных ответов, чтобы поддерживать их мотивацию.
Анализ поведенческих данных
Запросить подробный протокол ANOVA с повторными измерениями был выполнен для процента правильных ответов и среднего времени реакции с направлением взгляда (прямой/отраженный), эмоциями (гнев/страх) и интенсивностью (7 уровней морфов) в качестве внутрисубъектных факторов.
Выбор модели
Запросить подробный протокол Мы выполнили анализ поведенческих данных с помощью модели, чтобы охарактеризовать наблюдаемое повышение точности распознавания комбинаций взгляда и эмоций УГРОЗА+. Мы использовали выбор байесовской модели на основе модельных свидетельств (оцененных с помощью 10-кратной перекрестной проверки оценки логарифмической правдоподобия модели, которая неявно наказывает за сложность модели, не полагаясь на конкретные приближения, такие как байесовский информационный критерий или информационный критерий Акаике). . Мы применяли статистику фиксированных и случайных эффектов, ранее описанную в литературе. Сравнение с фиксированными эффектами предполагает, что все участники использовали одну и ту же базовую модель для создания своего поведения, так что общее свидетельство модели для данной модели пропорционально произведению свидетельства модели для модели для всех участников. Основываясь на данных этой модели, мы сравнили различные модели, вычислив их байесовский фактор как отношение модельных данных сравниваемой модели (Jeffreys, 1961; Касс и Рафтери, 1995). Сравнение случайных эффектов является более консервативным, позволяя разным участникам использовать разные модели для создания своего поведения, и направлено на вывод о распределении по моделям, которые участники используют (Penny et al., 2010). Для этого сравнения мы рассчитали поддержку победившей модели с помощью вероятности превышения ( p exc ), которая представляет собой вероятность того, что участники с большей вероятностью выберут эту модель для создания поведения по сравнению с любой альтернативной моделью.
Мы начали с простейшей модели (модель 0), которая могла бы объяснить решения каждого испытуемого, используя зашумленную психометрическую модель, подобную «обнаружению сигнала», в которую мы включили скорость отклонения, тем самым учитывая, что испытуемые угадывали случайным образом определенную долю испытаний:
P(гнев)=ϕ[w*x+b]*(1-ε)+0,5*ε
где P(гнев) соответствует вероятности оценки лица как гневного, Ф[.] до кумулятивная нормальная функция w к перцептивной чувствительности к отображаемой эмоции, x до пробного массива значений доказательств в пользу гнева или страха (сила эмоции, от -7 для сильного выражения страха до +7 для сильного выражения гнева), b до аддитивного, независимого от стимула смещение в сторону одной из двух эмоций, а ε — доля случайных предположений среди вариантов выбора. Мы сравнили «нулевую» модель, которая не учитывала влияние контекста направления взгляда на процесс принятия решения, с двумя дополнительными моделями, реализующими два разных механизма, которые могли бы объяснить наблюдаемое повышение точности распознавания комбинаций взгляда и эмоций «УГРОЗА+». Первая возможность (модель 1) заключается в том, что направление взгляда искажает распознавание эмоций в пользу интерпретации, сигнализирующей о более высокой угрозе (гнев при прямом взгляде, страх при отведенном взгляде). В качестве альтернативы (модель 2) направление взгляда может избирательно повышать чувствительность к эмоциям, сигнализируя о более высокой угрозе в этом контексте (смоделировано различной чувствительностью к эмоциям в условиях УГРОЗА+ и УГРОЗА-).
Регистрация и предварительная обработка ЭЭГ
Запросить подробный протокол Колпачок ЭЭГ из 63 спеченных кольцевых электродов Ag/AgCl (Easycap) использовался для записи активности ЭЭГ. Активность ЭЭГ записывалась с частотой дискретизации 1000 Гц с использованием усилителя BRAINAMP (Brain Products, BRAINAMP MR PLUS) и онлайн-фильтра нижних частот на частоте 250 Гц. Опорный канал располагался у них на носу, и использовалась площадка на лбу. Полное сопротивление поддерживалось ниже порога 10 кОм.
Необработанные данные ЭЭГ были пересчитаны в среднее значение, подвергнуты понижению дискретизации до 500 Гц, низкочастотному фильтру на частоте 32 Гц и эпохе от 1 с до до 4 с после начала лицевого стимула с использованием EEGLAB (Delorme and Makeig, 2004) . Во-первых, визуальные данные ЭЭГ были проверены для удаления мышечных артефактов и выявления зашумленных электродов, которые были интерполированы в среднее значение соседних электродов. Во-вторых, анализ независимых компонентов (ICA), который исключал интерполированные электроды, был выполнен для данных эпохи, а компоненты ICA, фиксирующие артефакты моргания, были вручную отклонены. Наконец, была проведена визуальная проверка полученных отдельных эпох, чтобы исключить любые оставшиеся испытания с артефактами. После пробных отказов в среднем 9Осталось 99 ± 10 испытаний на субъекта.
Частотно-временной анализ был выполнен с использованием набора инструментов Fieldtrip для MATLAB (Oostenveld et al. , 2011). Нас особенно интересовали моторные мю-диапазоны (8–32 Гц), поэтому мы оценили спектральную мощность колебаний ЭЭГ в мю-бета-диапазоне с помощью «многоступенчатого» частотно-временного преобразования (снижение Слепяна, частотный диапазон 8–32 Гц, пять циклов, три конусов на окно). Целью этого подхода с множественным сужением является получение более точных оценок мощности путем сглаживания частот. Обратите внимание, что это частотно-временное преобразование использует постоянное количество циклов на окно по частотам, следовательно, временное окно, продолжительность которого уменьшается обратно пропорционально увеличению частоты.
Анализ ЭЭГ
Временная частота: меры латерализации двигателя
Запросить подробный протокол Поскольку подавление мю-бета-активности в полушарии, противоположном руке, используемой для ответа, является маркером двигательной подготовки к ответу (Donner et al., 2009; de Lange et al., 2013), спектральная мощность от 8 до 32 Гц рассчитывали на каждом электроде и в момент времени для всех субъектов. Затем для каждого субъекта, чтобы получить показатели латерализации, спектральная мощность от 8 до 32 Гц для испытаний, в которых субъекты реагировали правой рукой, вычиталась из испытаний, в которых субъекты реагировали левой рукой. После усреднения по всем испытуемым были выбраны электроды, в которых моторная латерализация была максимальной от 200 мс до времени ответа: «П3», «КП3», «С3» для левого полушария и «П4, «КП4», «С4» для левого полушария. правое полушарие. Моторная латерализация, характерная для реакций «гнев» или «страх», была получена путем учета используемой карты «Гнев/Страх» и вычитания спектральной активности руки «Гнев» из спектральной активности руки «Страх» (среднее значение для «P3, «CP3», «C3» минус среднее значение «P4», «CP4», «C4», если участники ответили «Гнев» левой рукой, и наоборот, если они ответили «Гнев» правой рукой).
Регрессионный анализ: кодирование эмоциональной информации
Запросить подробный протокол В нашей задаче категоризации эмоций сила доказательства соответствует интенсивности отображаемой эмоции. Поэтому на основе недавних исследований (Wyart et al., 2012b, 2015) мы выполнили однократную регрессию сигналов ЭЭГ в зависимости от этой переменной. Была использована общая модель линейной регрессии (GLM), в которой сила эмоции (от 0 для нейтрального/безэмоционального выражения до 7 для сильного выражения страха/гнева) была введена в качестве предиктора широкополосных сигналов ЭЭГ в каждом испытании в каждый момент времени. после начала действия стимула (от 200 мс до и через 1 с после начала действия стимула) на каждом электроде. Соответствующие оценки параметров регрессии, представленные в произвольных единицах, были измерены для каждого участника, а затем усреднены по участникам для получения средних значений на уровне группы. Временной ход оценок параметров описывает нейронное «кодирование» релевантной (эмоциональной) информации, предоставляемой представленным выражением лица. Электроды и временные точки, в которых оценки параметров регрессии были максимальными, были выбраны для дальнейшего сравнения между интересующими условиями: прямой гнев и предотвращенный страх против прямого страха и предотвращенного гнева.
Подобные общие линейные регрессии были также выполнены для латерализованной активности мю-бета. Еще раз, интенсивность эмоционального выражения была введена как регрессор, чтобы предсказать активность двигательной латерализации от попытки к попытке (рассчитанную, как описано выше) для каждой временной точки после начала действия стимула. Единственное важное отличие заключается в том, что из-за «знакового» характера индекса моторной латерализации (положительного для контралатеральной активности) мы выражали интенсивность эмоционального выражения, как знак отображаемой эмоции, от -7 для интенсивного выражения. страха до +7 за сильное выражение гнева.
Корреляционный анализ нейронного выбора
Запросить подробный протокол Мы определили, модулируют ли остаточные флуктуации сигналов ЭЭГ в однократной попытке, необъяснимые изменениями в силе эмоции (измеряемой с помощью предыдущих нейронных регрессий в зависимости от силы эмоции), распознавание последующей эмоции. Этот подход напоминает меры «вероятности выбора», применяемые в электрофизиологии для измерения корреляции между нейронной активностью и поведением выбора (Britten et al., 19).96; Шадлен и др., 1996; Parker and Newsome, 1998) – путем оценки того, насколько флуктуации записанных нейронных сигналов «считываются» последующим решением (Wyart et al., 2012a, 2015). Преимущество измерения корреляций нейронного выбора в рамках нашей вычислительной модели состоит в том, что мы могли не только установить или , но и то, как нейронные флуктуации повлияли на последующее поведение — либо аддитивно как независимое от стимула смещение, либо мультипликативно как изменение перцептивной чувствительности.
На практике мы оценивали параметры b mod и w mod этих терминов нейронной модуляции в каждый момент времени после представления лица с помощью ЭЭГ-информированной регрессии выбора, для которой нейронные остатки e из регрессии против силы эмоции вводились либо отдельно (аддитивное влияние, параметр b mod , модель 1), либо как их взаимодействие с силой отображаемой эмоции (мультипликативное влияние, параметр w mod , модель 2) в качестве дополнительного предиктора последующего категориального выбора следующим образом:
1. p(гнев)=ϕ(w·x+b+bmod·e)
2. p( гнев)=ϕ(w·x+b+wmod·e·x)
Мы применили выбор байесовской модели для сравнения этих двух возможных модуляций процесса принятия решений нейронными флуктуациями с использованием как фиксированных, так и случайных эффектов, описанных выше. выше.
Статистические процедуры ЭЭГ
Запросить подробный протокол Все регрессионные анализы данных ЭЭГ проводились независимо для каждого субъекта, а затем проводился анализ второго уровня на уровне группы для оценки значимости наблюдаемых эффектов у участников. Анализ второго уровня опирался на стандартные параметрические тесты (t-тесты, дисперсионный анализ с повторными измерениями) с явным контролем частоты ошибок типа 1, возникающей в результате множественных сравнений в разные моменты времени, с помощью непараметрической статистики на уровне кластера, как описано в (Марис и Оостенвельд, 2007 г.). Спаривание между экспериментальными условиями и сигналами ЭЭГ перемешивалось псевдослучайно 1000 раз, и максимальная статистика на уровне кластера (сумма t-значений в смежных значимых моментах времени на пороговом уровне 0,05) извлекалась для каждого перемешивания для вычисления «нулевое» распределение размера эффекта во временном окне [-200, +1000] мс вокруг предъявления стимула или [-1000, +200] вокруг начала ответа. Для каждого значимого кластера в исходных (не перемешанных) данных мы вычислили долю кластеров в нулевом распределении, статистика которых превышала полученную для рассматриваемого кластера, что соответствует его «скорректированному по кластеру» p-значению. Мы применили второй метод начальной загрузки, чтобы проверить значительные сдвиги в латентности нейронного кодирования между состояниями, используя процедуру «складного ножа», описанную в (Kiesel et al., 2008).
Факторы Байеса были рассчитаны для критического отсутствия наблюдаемых эффектов, чтобы отличить недостаточную чувствительность тестов от подлинного отсутствия различий (Dienes, 2011). Байесовский фактор случайных эффектов на групповом уровне был рассчитан при тех же предположениях, что и стандартный тест t , который утверждает, что распределение наблюдаемого эффекта среди отдельных лиц может быть аппроксимировано нормальным распределением среднего значения (µ) и стандартного отклонения ( о). Мы рассчитали максимальное логарифмическое правдоподобие модели в пользу «нулевой» гипотезы, которая предполагает, что µ=0, и модель в пользу гипотезы «эффекта», для которой и µ, и σ могут быть свободно приспособлены к наблюдаемым данные. Затем мы использовали байесовский информационный критерий для сравнения двух моделей и вычисления соответствующего коэффициента Байеса. Коэффициент Байеса ниже 1/3 обеспечивает существенные доказательства в пользу нулевой гипотезы, тогда как коэффициент Байеса > 3 свидетельствует в пользу гипотезы эффекта.
Мы провели контрольный анализ, чтобы подтвердить надежность корреляции тревоги и угрозы у людей, наблюдаемых в височных и двигательных областях. Для этой цели мы выполнили процедуру перекрестной проверки с исключением одного, в которой мы вычислили «перекрестно проверенные» интервалы прогнозирования для линии регрессии на уровне группы по показателю тревоги участника № n, когда данные для участника № n был исключен из регрессии на уровне группы. Эта процедура была повторена для 24 участников для корреляции тревоги и угрозы в двигательных областях и показала, что нервные эффекты у двух участников немного снизились (<20%) за пределами перекрестно проверенных 95% интервалы предсказания. Повторный расчет корреляции на уровне группы в двигательных областях после исключения двух выбросов, оставив в анализе 22 участника, привел к значимому эффекту ( r = 0,48, d.f. = 20, p = 0,02). Этот исключающий анализ не выявил каких-либо отклонений для корреляции тревоги и угрозы в височных областях.
Анализ реконструкции источника
Запросить подробный протокол Анализ источника был выполнен с помощью Brainstorm (Tadel et al., 2011). Исходная модель, состоящая из 15 002 токовых диполей, использовалась для расчета матриц инверсии ядра для каждого субъекта на основе всех испытаний субъекта. Ориентация диполей была ограничена корковой мантией общей модели мозга, взятой из стандартного шаблона мозга Монреальского неврологического института (MNI), предоставленного в ходе мозгового штурма. Отдельные модели скальпа, записанные с помощью устройства Zebris, использовались для деформации этой шаблонной модели головы в колпачки датчиков ЭЭГ. Используя модель OpenMEEG BEM (Kybic et al. , 2005; Gramfort et al., 2010), для каждого субъекта была рассчитана прямая модель ЭЭГ. Затем были извлечены отдельные инверсионные матрицы (15002 вершины * 63 электрода) для выполнения однократной пробной регрессии на исходном уровне.
Эксперимент по рейтингу угроз и надежности
Запросить подробный протокол В эксперименте приняли участие 20 человек (10 женщин, средний возраст = 22,7 ± 0,6). 36 идентичностей, использованных в эксперименте, предъявлялись только в нейтральном состоянии. Каждая идентичность предъявлялась дважды, один раз с прямым и один раз с отведенным взглядом. На экране на 2 секунды появлялись лица, после чего они исчезали и на экране рисовались 2 сплошные шкалы. Участники оценивали личности по этим шкалам с точки зрения угрозы и надежности от «совсем нет» до «очень сильно» (в верхней части шкалы появился текст, напоминающий инструкцию: Насколько это лицо опасно/надежно?). Порядок весов был рандомизирован среди испытуемых. Шкалы оставались на экране до тех пор, пока не были даны два ответа, однако испытуемых проинструктировали отвечать интуитивно, не тратя слишком много времени на принятие решения.
Каталожные номера
- Адамс РБ
- Франклин RG
- Кверага К
- Амбади N
- Клецк RE
- Уэлен PJ
- Хаджихани N
- Нельсон AJ
(2012) Реакция миндалевидного тела на страх отведенного и прямого взгляда варьируется в зависимости от скорости презентации
Социальная когнитивная и аффективная неврология 7 :568–577.
https://doi.org/10.1093/scan/nsr038
- Google ученый
- Адамс РБ
- Клецк RE
(2003) Воспринимаемое направление взгляда и обработка выражений эмоций на лице
Психологическая наука 14 :644–647.
https://doi.org/10.1046/j.0956-7976.2003.psci_1479.x
- Google ученый
- Адамс РБ
- Клецк RE
(2005) Влияние прямого и отведенного взгляда на восприятие эмоций, передаваемых по лицу
Эмоция 5 :3–11.
https://doi.org/10.1037/1528-3542.5.1.3
- Google ученый
- Авиезер Н
- Бентин С
- Дударев В
- Хассин RR
(2011) Автоматия интеграции эмоционального лица и контекста
Эмоция 11 :1406–1414.
https://doi.org/10.1037/a0023578
- Google ученый
- Балкон М
- Бортолотти А
(2012) Обнаружение выражения эмоций на лице и показатели самоотчета в эмпатических ситуациях зависят от торможения сенсомоторной цепи с помощью низкочастотной рТМС
Стимуляция мозга 5 :330–336.
https://doi.org/10.1016/j.brs.2011.05.004
- Google ученый
- Балкон М
- Бортолотти А
(2013) «Моделирование» мимического выражения эмоций при кратковременном и длительном воздействии стимула.
эффект торможения премоторной коры с помощью rTMSBrain and Cognition 83 : 114–120.
https://doi.org/10.1016/j.bandc.2013.07.003
- Google ученый
- Барретт ЛФ
- Кенсингер EA
(2010) Контекст обычно кодируется во время восприятия эмоций
Психологическая наука 21 :595–599.
https://doi.org/10.1177/0
7610363547- Google ученый
- Бейтсон М
- Брилот Б
- Крапива D
(2011)
Тревога: эволюционный подход
Canadian Journal of Psychiatry 56 :707–715.
- Google ученый
Книга
- Бек AT
- Эмери Г
- Гринберг RL
(1985)
Тревожные расстройства и фобии: когнитивный взгляд
Нью-Йорк: Основные книги.
- Google ученый
- Биндеманн М
- Майк Бертон A
- Лэнгтон SRH
(2008) Как взаимодействуют взгляд и выражение лица?
Зрение 16 :708–733.
https://doi.org/10.1080/13506280701269318
- Google ученый
- Епископ SJ
(2007) Нейрокогнитивные механизмы тревоги: интегративный отчет
Trends in Cognitive Sciences 11 :307–316.
https://doi.org/10.1016/j.tics.2007.05.008
- Google ученый
- Боргоманери S
- Газзола V
- Авенанти А
(2014) Временная динамика возбудимости моторной коры при восприятии естественных эмоциональных сцен
Социально-когнитивная и аффективная неврология 9 :1451–1457.
https://doi.org/10.1093/scan/nst139
- Google ученый
- Брейнард DH
(1997) Набор инструментов психофизики
Пространственное зрение 10 :433–436.
https://doi.org/10.1163/156856897X00357
- Google ученый
- Бриттен KH
- Ньюсом WT
- Шадлен MN
- Селебрини S
- Мовшон JA
(1996) Взаимосвязь между поведенческим выбором и зрительными реакциями нейронов макаки MT
Визуальная неврология 13 :87–100.
https://doi.org/10.1017/S0
380000715X- Google ученый
Книга
- Брюхон-Швейцер М
- Полхан I
(1993)
Le Manuel Du STAI-Y De CD Spielberger, Французская адаптация
Париж: ECPA.
- Google ученый
- Цислер JM
- Костер EH
(2010) Механизмы смещения внимания к угрозе при тревожных расстройствах: интегративный обзор
Clinical Psychology Review 30 :203–216.
https://doi.org/10.1016/j.cpr.2009.11.003
- Google ученый
- Конти Л
- Дезекеш G
- Хьювиль L
- Грез J
(2012) Раннее связывание взгляда, жестов и эмоций: нейронный ход времени и корреляты
Журнал неврологии 32 :4531–4539.
https://doi.org/10.1523/JNEUROSCI.5636-11.2012
- Google ученый
Книга
- Дарвин С
(1872 г.) Выражение эмоций у человека и животных (1-е издание)
Лондон: Джон Мюррей.
https://doi.org/10.1037/10001-000
- Google ученый
- де Ланге FP
- Рахнев Д.А.
- Доннер TH
- Лау Х
(2013) Предстимульная колебательная активность моторной коры отражает перцептивные ожидания
Журнал неврологии 33 :1400–1410.
https://doi.org/10.1523/JNEUROSCI.1094-12.2013
- Google ученый
- Делорм А
- Макейг S
(2004) EEGLAB: набор инструментов с открытым исходным кодом для анализа динамики ЭЭГ в одной попытке, включая анализ независимых компонентов
Journal of Neuroscience Methods 134 :9–21.
https://doi.org/10.1016/j.jneumeth.2003.10.009
- Google ученый
- Диенес Z
(2011) Байесовская статистика против ортодоксальной: на чьей вы стороне?
Перспективы психологической науки 6 : 274–290.
https://doi.org/10.1177/17456
406920
- Google ученый
- Доннер TH
- Сигель М
- Фри P
- Энгель АК
(2009) Нарастание предиктивно-выборочной активности в моторной коре человека при перцептивном принятии решений
Современная биология 19 :1581–1585.
https://doi.org/10.1016/j.cub.2009.07.066
- Google ученый
- Эль Зейн М
- Гамонд Л
- Конти Л
- Грез J
(2015) Влияние избирательного внимания на раннюю интеграцию социальных сигналов: одинаковое время, модулированные нейронные источники
NeuroImage 106 : 182–188.
https://doi.org/10.1016/j.neuroimage.2014.10.063
- Google ученый
- Эткин А
- Ставка ТД
(2007) Функциональная нейровизуализация тревоги: метаанализ обработки эмоций при посттравматическом стрессовом расстройстве, социальном тревожном расстройстве и специфической фобии
Американский журнал психиатрии 164 :1476–1488.
https://doi.org/10.1176/appi.ajp.2007.07030504
- Google ученый
- Юбэнк MP
- Фокс Е
- Колдер AJ
(2010) Взаимодействие между взглядом и выражением лица в миндалине и расширенной миндалине модулируется тревогой
Frontiers in Human Neuroscience 4 :56.
https://doi.org/10.3389/fnhum.2010.00056
- Google ученый
- Фрижда NH
(2009) Переживание эмоций и их разновидности
Обзор эмоций 1 :264–271.
https://doi.org/10.1177/1754073
3595
- Google ученый
- Золото JI
- Шадлен MN
(2007) Нейронная основа принятия решений
Annual Review of Neuroscience 30 :535–574.
https://doi.
org/10.1146/annurev.neuro.29.051605.113038- Google ученый
- Грэм Р
- ЛаБар КС
(2007) Интерференция Гарнера выявляет зависимость между эмоциональным выражением и взглядом при восприятии лица
Эмоция 7 :296–313.
https://doi.org/10.1037/1528-3542.7.2.296
- Google ученый
- Грамфорт А
- Пападопуло Т
- Оливи Е
- Клерк М
(2010) OpenMEEG: программное обеспечение с открытым исходным кодом для квазистатической биоэлектромагнетики
Biomedical Engineering Online 9 :45.
https://doi.org/10.1186/1475-925Х-9-45
- Google ученый
Книга
- Зеленый ДМ
- Свет JA
(1966)
Теория обнаружения сигналов и психофизика
Нью-Йорк: John Wiley and Sons.
- Google ученый
- Группа DW
- Ничке JB
(2013) Неопределенность и ожидание при тревоге: комплексная нейробиологическая и психологическая перспектива
Nature Reviews Neuroscience 14 :488–501.
https://doi.
org/10.1038/nrn3524- Google ученый
- Грез J
- Валабрег R
- Голипур B
- Шевалье C
(2014) Прямая миндалевидно-моторная цепочка для эмоциональных проявлений, влияющих на действие: исследование диффузионно-тензорной визуализации
Картирование человеческого мозга 35 :5974–5983.
https://doi.org/10.1002/hbm.22598
- Google ученый
- Хекерен HR
- Маррет S
- Унгерлейдер LG
(2008) Нервные системы, которые опосредуют перцептивное принятие решений человеком
Обзоры природы.
Неврология 9 : 467–479.https://doi.org/10.1038/nrn2374
- Google ученый
Книга
- Джеффрис Х
(1961)
Теория вероятностей
Оксфорд, Великобритания: Издательство Оксфордского университета.
- Google ученый
Книга
- Джонсон Р
(1988)
Амплитуда компонента P300 связанного с событием потенциала: обзор и синтез
В: Ackles P.K., Jennings J.R., Coles M.G.H, editors. Достижения в области психофизиологии . Гринвич, Коннектикут: JAI Press.
стр. 693–137.- Google ученый
- Касс RE
- Стропила AE
(1995) Факторы Байеса
Журнал Американской статистической ассоциации 90 :773–795.
https://doi.org/10.1080/01621459.1995.10476572
- Google ученый
- Кизель А
- Миллер Дж
- Жоликер P
- Бриссон Б
(2008) Измерение различий в задержке ERP: сравнение методов оценки с одним участником и методом складного ножа
Психофизиология 45 :250–274.
https://doi.org/10.1111/j.1469-8986.2007.00618.x
- Google ученый
- Ключарев В
- Самс М
(2004) Взаимодействие направления взгляда и обработки выражений лица: исследование ERP
НейроОтчет 15 :621–625.
https://doi.org/10.1097/00001756-200403220-00010
- Google ученый
- Кок П
- Джехи JF
- де Ланге FP
(2012) Чем меньше, тем лучше: ожидание обостряет представления в первичной зрительной коре
Нейрон 75 : 265–270.
https://doi.org/10.1016/j.neuron.2012.04.034
- Google ученый
- Кибик Дж
- Клерк М
- Аббуд Т
- Фогерас О
- Керивен Р
- Пападопуло Т
(2005) Общий формализм для интегральных формулировок прямой задачи ЭЭГ
IEEE Transactions on Medical Imaging 24 : 12–28.
https://doi.org/10.1109/TMI.2004.837363
- Google ученый
- Лангнер О
- Дотч Р
- Бийлстра G
- Вигболдус DHJ
- Ястреб СТ
- фургон Книппенберг A
(2010) Презентация и проверка базы данных лиц radboud
Познание и эмоции 24 :1377–1388.
https://doi.org/10.1080/02699930
5076
- Google ученый
- Леду J
(2012) Переосмысление эмоционального мозга
Нейрон 73 : 653–676.
https://doi.org/10.1016/j.neuron.2012.02.004
- Google ученый
Книга
- Макмиллан Н.А.
- Крилман CD
(2004)
Теория обнаружения: Руководство пользователя
Нью-Йорк: Psychology Press.
- Google ученый
- Марс РБ
- Дебенер S
- Гладвин TE
- Харрисон LM
- Хаггард П
- Ротвелл JC
- Бестманн S
(2008) Колебания событийной электроэнцефалограммы от одной попытки к другой отражают динамические изменения степени неожиданности
Журнал неврологии 28 :12539–12545.
https://doi.org/10.1523/JNEUROSCI.2925-08.2008
- Google ученый
- Нинборг H
- Камминг Б
(2010) Корреляции между активностью сенсорных нейронов и поведением: как много они говорят нам о причинно-следственной связи нейрона?
Текущее мнение в области нейробиологии 20 : 376–381.
https://doi.org/10.1016/j.conb.2010.05.002
- Google ученый
- Нинборг H
- Камминг БГ
(2009) Активность сенсорных нейронов, связанная с принятием решений, отражает больше, чем причинный эффект нейрона
Природа 459 :89–92.
https://doi.org/10.1038/nature07821
- Google ученый
- Н’Диай К
- Сандер Д
- Вийомье P
(2009) Обработка релевантности в миндалевидном теле человека: направление взгляда, выражение лица и интенсивность эмоций
Эмоция 9 :798–806.
https://doi.org/10.1037/a0017845
- Google ученый
- О’Коннелл RG
- Докри PM
- Келли СП
(2012) Супрамодальный сигнал накопления-в-связке, который определяет перцептивные решения у людей
Nature Neuroscience 15 :1729–1735.
https://doi.org/10.1038/nn.3248
- Google ученый
- Оман А
- Минека S
(2001) Страхи, фобии и готовность: к усовершенствованному модулю страха и обучения страху
Психологический обзор 108 :483–522.
https://doi.org/10.1037/0033-295X.108.3.483
- Google ученый
- Остенвельд R
- Фри P
- Марис Э
- Шоффелен JM
(2011) FieldTrip: программное обеспечение с открытым исходным кодом для расширенного анализа данных МЭГ, ЭЭГ и инвазивных электрофизиологических данных
Computational Intelligence and Neuroscience 2011 :156869.
https://doi.org/10.1155/2011/156869
- Google ученый
- Паркер AJ
- Ньюсом WT
(1998) Чувство и отдельный нейрон: исследование физиологии восприятия
Annual Review of Neuroscience 21 : 227–277.
https://doi.org/10.1146/annurev.neuro.21.1.227
- Google ученый
- Пелли ДГ
(1997) Программное обеспечение VideoToolbox для визуальной психофизики: преобразование чисел в фильмы
Spatial Vision 10 :437–442.
https://doi.org/10.1163/156856897X00366
- Google ученый
- Пенни WD
- Стефан К.Е.
- Донизо J
- Роза MJ
- Фристон KJ
- Шофилд ТМ
- Лефф АП
(2010) Сравнение семейств динамических причинно-следственных моделей
PLoS Вычислительная биология 6 :e1000709.
https://doi.org/10.1371/journal.pcbi.1000709
- Google ученый
- Фелпс EA
- Линг С
- Карраско М
(2006) Эмоции облегчают восприятие и усиливают перцептивные преимущества внимания
Психологическая наука 17 : 292–299.
https://doi.org/10.1111/j.1467-9280.2006.01701.x
- Google ученый
- Портуа G
- Спинелли L
- Сек М
- Вийомье P
(2010) Модуляция обработки лица эмоциональным выражением и направлением взгляда во время внутричерепных записей в правой веретенообразной коре
Журнал когнитивной неврологии 22 :2086–2107.
https://doi.org/10.1162/jocn.2009.21404
- Google ученый
- Рахнев Д
- Лау Х
- де Ланге FP
(2011) Предшествующее ожидание модулирует взаимодействие между сенсорными и префронтальными областями человеческого мозга
Журнал неврологии 31 :10741–10748.
https://doi.org/10.1523/JNEUROSCI.1478-11.2011
- Google ученый
- Ригарт Р
- де Гелдер B
(2008) Эмоциональная суть сцены влияет на распознавание выражений лица
Когнитивная, эмоциональная и поведенческая неврология 8 :264–272.
https://doi.org/10.3758/CABN.8.3.264
- Google ученый
- Сандер Д
- Гранжан Д
- Кайзер S
- Верле Т
- Шерер КР
(2007) Эффекты взаимодействия воспринимаемого направления взгляда и динамического выражения лица: доказательства теории оценки эмоций
Европейский журнал когнитивной психологии 19 :470–480.
https://doi.org/10.1080/0
- 40600757426
- Google ученый
- Сато W
- Йошикава С
- Котияма Т
- Мацумура М
(2004) Миндалевидное тело обрабатывает эмоциональную значимость выражений лица: фМРТ-исследование с использованием взаимодействия между выражением и направлением лица
NeuroImage 22 :1006–1013.
https://doi.org/10.1016/j.neuroimage.2004.02.030
- Google ученый
- Шадлен MN
- Бриттен KH
- Ньюсом WT
- Мовшон JA
(1996)
Компьютерный анализ взаимосвязи между нейронными и поведенческими реакциями на визуальное движение
The Journal of Neuroscience 16 :1486–1510.
- Google ученый
Книга
- Спилбергер CD
- Горсуч RL
- Лушене Р
- Вагг PR
- Джейкобс Джорджия
(1983)
Руководство по инвентаризации тревожных состояний
Пало-Альто, Калифорния: Consulting Psychologists Press.
- Google ученый
- Тадель Ф
- Байет S
- Мошер JC
- Пантазис Д
- Лихи RM
(2011) Brainstorm: удобное приложение для анализа МЭГ/ЭЭГ.
https://doi.org/10.1155/2011/879716
- Google ученый
- Вайарт В
- де Гардель V
- Шолль J
- Саммерфилд C
(2012) Ритмические колебания накопления доказательств при принятии решений в мозгу человека
Нейрон 76 :847–858.
https://doi.org/10.1016/j.neuron.2012.09.015
- Google ученый
- Вайарт В
- Майерс NE
- Саммерфилд C
(2015) Нейронные механизмы перцептивного выбора человека при сфокусированном и разделенном внимании
Журнал неврологии 35 :3485–3498.
https://doi.org/10.1523/JNEUROSCI.3276-14.2015
- Google ученый
- Вайарт В
- Нобре AC
- Саммерфилд C
(2012) Диссоциируемые априорные влияния вероятности и релевантности сигнала на визуальную контрастную чувствительность
Труды Национальной академии наук Соединенных Штатов Америки 109 : 3593–3598.
https://doi.org/10.1073/pnas.1120118109
- Google ученый
Письмо-решение
Благодарим вас за отправку вашей работы под названием «Тревога диссоциирует адаптивные функции сенсорных и моторных реакций на социальные угрозы» для экспертной оценки в eLife . Ваш материал был положительно оценен Тимоти Беренсом (старшим редактором) и тремя рецензентами, один из которых, Пегги Мейсон, является членом нашего Совета редакторов-рецензентов.
Рецензенты обсудили обзоры друг с другом, и редактор-рецензент подготовил это решение, чтобы помочь вам подготовить исправленное представление.
Взаимодействие эмоционального выражения с направлением взгляда в зависимости от чувствительности усиления элегантно, хорошо сделано и ново. Зависимый от тревоги анализ и связанные с ним результаты интригуют. Решение следующих вопросов сделало бы это ценным вкладом.
Самое главное, Методы должны быть включены в Результаты. Методы слишком важны для понимания результатов, чтобы представлять их отдельно. Пожалуйста, напишите рукопись так, чтобы она была понятна широкому читателю. Ясность важнее стандартного формата, и все рецензенты согласились с тем, что методологические детали должны быть включены в презентацию результатов.
Могут ли авторы обосновать, что они не учитывают реакцию участников в своем анализе ЭЭГ? Или, возможно, анализ данных стандартным способом – по стимулу – а затем дополнительно по классификационному ответу?
Есть опасения, что один выброс определяет корреляцию тревоги и угрозы для двигательных областей. Не могли бы авторы решить эту проблему?
Укажите, как учитывались множественные сравнения.
Обследовались ли пациенты на социальную тревожность или депрессию?
Вышеуказанные пункты являются основными, которые повлияли на решения рецензентов. Полные обзоры представлены ниже.
Рецензент №1:
Это интересное исследование с еще более интересным ответом, если добавить немного беспокойства. У меня не так много комментариев отчасти потому, что методы ЭЭГ непрозрачны для меня. Меня больше всего беспокоит, имеет ли ценность средняя часть результатов, касающаяся всех тем вместе. Похоже, что результаты на самом деле обусловлены двумя разными сильно значимыми результатами в двух разных когортах испытуемых — с высокой и низкой тревожностью. Таким образом, общая повышенная чувствительность к угрозе+ состоит из низкой тревожности — восприятия и высокой тревожности = моторных частей.
Рецензент №2:
В целом я думаю, что это хорошая работа, и авторы явно приложили значительные усилия, чтобы предоставить полный отчет о своем наборе данных, и им следует приветствовать объем анализа. проводится, чтобы лучше понять нейронную реакцию на угрожающие социальные стимулы. Однако я обнаружил, что часть анализа трудно разобрать, а в некоторых случаях важная информация отсутствовала (или была неочевидной).
Особенно меня беспокоило выборочное представление поведенческих результатов. Где результаты времени реакции (как уже упоминалось, они были проанализированы в методах)? Не было ли взаимодействия между производительностью категоризации и уровнем морфинга при изучении взаимодействия производительности взгляд-эмоция? Графики на рис. 2В показывают, что, как и следовало ожидать, эффект взгляда сильнее для более слабых сигналов эмоций (ближе к нейтральным). В качестве более второстепенного вопроса, почему участники не на 100% правы в отношении конечных точек морфов (100% гнев, 100% страх) — некоторые личности постоянно классифицируются неправильно?
Переходим к анализу ЭЭГ. Авторы используют передовые и относительно специализированные методы для изучения взаимосвязи сигнала ЭЭГ с модуляциями эмоциональной силы, и их результаты интересны и кажутся убедительными. Хотя я думаю, что они проделали разумную работу по объяснению этих методов и того, как их следует интерпретировать, мне хотелось бы получить более подробную информацию о статистических тестах, которые они использовали. Конечно, они выполнили некоторую загрузку, чтобы исправить проблемы с множественными сравнениями, с которыми они сталкиваются. На это намекает подпись к рисунку 3 (пороговое значение, скорректированное по кластеру), но никаких подробностей, которые я могу видеть, нигде не приводится.
Кроме того, я думаю, что авторы использовали все испытания (правильные и неправильные) в своем анализе ЭЭГ, но мне неясно, почему это может быть оправдано, особенно для более слабых эмоциональных стимулов (где участники верны только в 70% случаев в классифицировать их). Например, испытание «угроза + страх, предотвращенное страхом» могло быть классифицировано участником как лицо гнева (и, таким образом, рассматриваться участником как угроза-). Ожидали ли авторы, что нейронное кодирование будет одинаковым в обоих случаях? Если нет, то как они могут оправдать неучет ответов участников в своем анализе ЭЭГ?
Наконец-то мне понравился анализ зависимости от тревожности, и его результаты ясны и интересны.
Рецензент №3:
Wyart et al. исследовать взаимодействие эмоционального выражения и направления взгляда и протестировать две модели: мультипликативное усиление угрозы восприятия эмоций и аддитивное смещение реакции. Две модели четко объясняются со ссылкой на поведенческие данные, после чего авторы проверяют эти модели на данных одиночных пробных нейронных сетей. Авторы сообщают о временной зависимости чувствительности к условиям высокой и низкой угрозы. Они также проверяют модели смещения по сравнению с чувствительностью, используя остатки регрессионного анализа на обоих височных частичных электродах, и обнаруживают повышение перцептивной чувствительности, а также проверяют модели по индексу моторной латерализации и обнаруживают эффект смещения.
Эта часть статьи элегантна, хорошо сделана и, насколько мне известно, оригинальна. Раздел «Методы» действительно ясен — я бы предпочел, чтобы некоторые объяснения моделей были перенесены в основную часть документа, поскольку эта часть, хотя и является «технической», действительно необходима для понимания смысла последующего анализа. и в его нынешнем виде введение гораздо менее ясно, чем более подробное объяснение.
В последнем разделе статьи утверждается связь с уровнем тревожности у отдельных субъектов, поскольку угроза+ и — по-разному модулируют височные области у людей с низкой тревожностью), а угроза+ модулирует двигательные области у людей с высокой тревожностью.
У меня есть оговорки по поводу второй части диссоциации у лиц с высокой тревожностью, в основном связанной с рисунком 7Б – мне кажется, это главная слабость.
1) Тревога сочетается с депрессией и социальной тревожностью. Были ли участники проверены на предмет оценки других факторов, которые могут быть связаны с тревожностью?
2) Все участники кроме одного лежат в самом центре нормального диапазона. На рисунке 7B похоже, что оценка 40+ может быть выбросом. Значения R вычисляются с помощью Пирсона (я предполагаю, хотя я заметил, что этого нет на рисунке; это должно быть). Если удаленный субъект исключен, сохраняется ли корреляция?
Исходя из этого, мне кажется, что наблюдается увеличение изменчивости в группе «более тревожных», что может быть движущей силой корреляции, тем более что разница между угрозой и отсутствием угрозы в группе с высокой тревожностью группа в 7B не имеет значения.
3) Эффект на рисунке 7B кажется слабым эффектом в один момент времени. Нет обоснования/объяснения выбора ни окна в 7А, ни одиночной точки в 7Б (если это так).
4) Роль времени реакции: авторы говорят, что не обнаружили различий в поведенческих реакциях между более и менее тревожными группами. Это включало время реакции?
[Примечание редактора: дополнительные изменения были запрошены до принятия, как описано ниже.]
Благодарим вас за повторную отправку вашей работы под названием «Тревога диссоциирует адаптивные функции сенсорных и моторных реакций на социальные угрозы» для экспертной оценки по адресу . eLife . Ваше представление было положительно оценено Тимоти Беренсом (старшим редактором) и Пегги Мейсон (редактором-рецензентом).
Отличная версия. Вы использовали обзоры, чтобы сделать эту сложную статью настолько понятной, насколько того заслуживают ее интересные выводы. У редактора есть три очень незначительных предложения по уточнению (достаточно 1-2 предложений на выпуск). Вот вопросы:
Рисунок 1 очень полезен за исключением Я не понимал ось эмоций на рисунке 1 (закрашена серым цветом), пока не увидел нижнюю часть рисунка 3, где она обозначена как эмоциональная чувствительность. Пожалуйста, обозначьте на рисунке 1 и поясните в легенде (в дополнение к пояснению в тексте).
Как было выбрано время на рисунке 4? Время в B и C имеет для меня некоторый смысл, но я не понимаю, как было выбрано 280, когда оно так далеко от области существенных различий. Также, пожалуйста, объясните, почему карты в D-F не выглядят так, как если бы они были получены из мозга, нанесенного на карту в середине A-C (особенно B-C). Перечитав это, я думаю, что понимаю проблему синхронизации — вы отображаете отклонения от 0, а не пиковые различия между условиями. Может быть, сделать акцент на последнем так же, как и на первом, превратив серые прямоугольники в столбцы вверху, чтобы они соответствовали зеленым и оранжевым столбцам? Все одинаково подчеркнули именно так. Путаница в отношении выравнивания карты или его отсутствия остается.
Не могли бы вы для общего читателя добавить несколько слов о различиях в латерализации, показанных на рис. 6В и обсуждавшихся в подразделе «Раннее нейронное кодирование эмоций, сигнализирующих об угрозе, при двигательной подготовке»? Это кажется устойчивым отличием — известно ли оно и ожидается ли оно? Удивительно в любом случае? Помните, что eLife читательская аудитория широка и не обязательно хорошо осведомлена о большом внутреннем опыте.
https://doi.org/10.7554/eLife.10274.011
Ответ автора
Взаимодействие эмоционального выражения с направлением взгляда в зависимости от чувствительности усиления является элегантным, хорошо выполненным и новым. Зависимый от тревоги анализ и связанные с ним результаты интригуют. Решение следующих вопросов сделало бы это ценным вкладом.
Самое главное, Методы должны быть включены в Результаты. Методы слишком важны для понимания результатов, чтобы представлять их отдельно. Пожалуйста, напишите рукопись так, чтобы она была понятна широкому читателю. Ясность важнее стандартного формата, и все рецензенты согласились с тем, что методологические детали должны быть включены в презентацию результатов.
Мы последовали совету рецензентов, включив описание методов в основной текст. Мы стремились сделать результаты понятными для неспециалистов, не требуя от них подробного прочтения раздела «Методы». При этом мы решили оставить методологические подробности в разделе «Методы», а в основном тексте предоставить обоснование регрессионного анализа. Мы считаем, что эти изменения значительно улучшили читабельность рукописи для неспециалистов, и благодарим рецензентов за это полезное предложение.
Могут ли авторы обосновать отсутствие учета ответов участников в их анализе ЭЭГ? Или, возможно, анализ данных стандартным способом – по стимулу – а затем дополнительно по классификационному ответу?
Мы считаем, что этот комментарий тесно связан с предыдущим пунктом — описание методов в основном тексте должно прояснить, почему мы не проводили прямых сопоставлений на основе ответов участников (например, правильные ответы против ошибок). Мы не проводили таких контрастов, потому что соответствующие эффекты могут быть косвенно (и тривиально) опосредованы изменениями в самом стимуле, а не в процессе принятия решения. Например, как сигналы ЭЭГ, так и точность поведенческих реакций растут вместе с силой эмоции, отображаемой стимулом лица, так что классификация сигналов ЭЭГ в отношении точности реакции может показать значительную разницу, которая просто связана с тем фактом, что правильные ответы в среднем были связаны с более сильными эмоциями, т. е. внешними различиями в стимуляции. Выполнение анализа классификации ответов, не смешиваемого с изменениями в стимуляции, потребовало бы условия «постоянного стимула», например, рассмотрение безэмоциональных/нейтральных стимулов изолированно (только 1/15 всех испытаний в нашем экспериментальном плане). Другой подход, которого мы придерживались на протяжении всей рукописи и который позволяет анализировать все испытания, — это метод, основанный на регрессии, заключающийся в изучении того, как испытание за испытанием Остаточные значения регрессии характеристик стимула (здесь — силы эмоции) по отношению к сигналам ЭЭГ зависят от точности ответа. Другим преимуществом регрессионного анализа на основе моделей, который действительно принимает во внимание ответы участников и который мы более подробно подчеркиваем в пересмотренной рукописи, является то, что они позволяют не только оценить, коррелируют ли сигналы ЭЭГ с поведенческими реакциями, но также и то, как они делают это: через модуляции перцептивной чувствительности и/или предвзятости.
С теоретической точки зрения стоит отметить, что согласно теориям принятия решений, вдохновленным классической теорией обнаружения сигналов, процесс принятия решений, лежащий в основе правильных ответов и ошибок, одинаков — единственная разница заключается в положении зашумленной переменной решения по отношению к критерий принятия решения по конкретным испытаниям. Таким образом, определение чувствительности и предвзятости основывается как на правильных ответах, так и на ошибках. Следуя этим рассуждениям, мы решили провести регрессионный анализ данных ЭЭГ без искусственного отделения правильных ответов от ошибок. Таким образом, полученный паттерн указывает на усиление нервной чувствительности к эмоциям, сигнализирующим об угрозе 9.0141 независимо от точности последующего ответа . Тем не менее, даже при рассмотрении только правильных проб в анализе «кодирования», показанном на рис. 4, существенные различия между эмоциями УГРОЗА+ и УГРОЗА- появляются на височных электродах через 170 мс после начала стимула ( t 23 = -2,1, p < 0,05), теменные электроды на 500 мс ( t 23 = 4,2, p < 0,001) и двигательные сигналы на 200 мс ( t 23 = 3, p < 0,01). Мы добавили эти результаты в раздел «Результаты»:
«Это усиление, зависящее от угрозы, оставалось значимым при рассмотрении только правильных ответов (временные: t 23 = -2,1, p < 0,05; центрально-теменной t 23 = 4,2, p < 0,001)”,
и:
0141 t 23 = 3,0, p < 0,01).
Мы надеемся, что эти пояснения, отраженные в дополнительных описаниях регрессионных методов по всему тексту, сделают основные выводы более понятными для широкого круга читателей eLife .
Есть опасения, что корреляция тревога-угроза для двигательных областей определяется одним выбросом. Не могли бы авторы решить эту проблему?
Мы провели дополнительный контрольный анализ, чтобы исключить возможность того, что корреляция тревоги и угрозы, наблюдаемая в двигательных областях, обусловлена одним или несколькими выбросами. Во-первых, важно отметить, что показатели тревожности наших участников (от 20 до 45) хорошо попадают в диапазон нормальной популяции (< 60), и, таким образом, нет априорной причины исключать кого-либо из наших участников исключительно на основании по его/ее шкале тревожности. Кроме того, именно вариабельность показателей тревожности между участниками обусловила их корреляцию с нервными эффектами. В качестве первого контрольного анализа мы рассчитали интервалы прогнозирования для линии регрессии на уровне группы по показателю тревожности каждого участника. Ни один участник не попал за пределы 95% интервалов прогнозирования как для височной (рис. 8А), так и для двигательной области (рис. 8В) — и, таким образом, могут быть статистически помечены как выбросы.
Мы провели второй, более строгий контрольный анализ, чтобы дополнительно подтвердить надежность корреляции тревоги и угрозы, наблюдаемой в височных и двигательных областях. Мы выполнили процедуру перекрестной проверки с исключением одного исключения, в которой мы вычислили «перекрестно проверенные» интервалы прогнозирования для линии регрессии на уровне группы по показателю тревожности участника № n, когда данные для участника № n были исключены из регрессия на уровне группы. Эта процедура была повторена для 24 участников для корреляции тревоги и угрозы в двигательных областях и показала, что нервные эффекты у двух участников немного снизились (<20%) за пределами 9.0141 перекрестная проверка 95% интервалов прогнозирования, включая участника № 4, чья оценка тревожности составила 45. Тем не менее, повторное вычисление корреляции на уровне группы в двигательных областях после исключения двух выбросов, в результате чего в анализе осталось 22 участника, привело к значительному эффекту ( r = 0,48, ф.р. = 20, p = 0,02). Этот исключающий анализ не выявил каких-либо отклонений для корреляции тревоги и угрозы в височных областях.
Вместе мы считаем, что эти контрольные анализы подтверждают надежность корреляций тревоги и угрозы как для височных, так и для двигательных областей, и теперь мы опишем их в разделе «Методы» пересмотренной рукописи.
Укажите, как учитывались множественные сравнения.
Приносим свои извинения за то, что не предоставили достаточно статистических данных о поправке на множественные сравнения в разделе «Методы». Мы применили стандартные поправки, описанные в (Maris and Oostenveld, 2006), отраженные в «скорректированных по кластеру» p-значениях, представленных в разделе «Результаты». Теперь мы прямо указываем, как учитывались множественные исправления, в разделе «Методы» пересмотренной рукописи в новом подразделе «статистические процедуры» («Все регрессионные анализы […] соответствуют его «кластерно-скорректированному» p-значению»). .
Обследовались ли пациенты на социальную тревожность или депрессию?
Этот вопрос связан с растущим количеством данных в пользу коморбидности между тревогой (связанной с шестью тревожными расстройствами, среди которых социальная тревога) и депрессивными расстройствами, что требует выявления как специфических, так и общих нейрокогнитивных маркеров тревоги и депрессии.
Наши результаты выдвинули адаптивную и положительную функцию умеренной (неклинической) тревожности в ответ на социальные угрозы (Bateson et al., 2011). Наши участники заполнили анкету STAI, наиболее часто используемую в экспериментальных исследованиях тревожных признаков в неклинических выборках (см., например, Sylvers et al., 2011), но они не были обследованы ни на депрессию, ни на социальную тревожность. Как недавно подчеркнули Grupe and Nitschke (2013): «Несмотря на отсутствие специфичности, актуальность исследований с использованием STAI подчеркивается его чувствительностью в качестве маркера риска тревожных расстройств». Кроме того, что касается коморбидности депрессии и тревоги, было показано, что у пациентов с клинически выраженной тревогой и депрессией уровень смертности был ниже, чем у пациентов только с депрессией (Mykletun et al., 2009).). Таким образом, это открытие предполагает, что как неклиническая, так и клиническая тревога могут иметь специфическую «защитную» функцию, связанную с адаптивной ролью тревоги в выживании, которая не является общей для депрессии.
Тем не менее, мы согласны с рецензентами и признаем вытекающее из этого ограничение в отношении специфичности наблюдаемых зависимых от тревоги эффектов. В связи с этим мы добавили предложение в раздел «Обсуждение»:
«Дальнейшие исследования должны оценить специфичность этих зависимых от тревоги эффектов в свете растущего количества доказательств в пользу коморбидности между тревогой и депрессивными расстройствами».
Рецензент №1:
Это интересное исследование с еще более интересным ответом, если добавить немного беспокойства. У меня не так много комментариев отчасти потому, что методы ЭЭГ непрозрачны для меня. Меня больше всего беспокоит, имеет ли ценность средняя часть результатов, касающаяся всех тем вместе. Похоже, что результаты на самом деле обусловлены двумя разными сильно значимыми результатами в двух разных когортах испытуемых — с высокой и низкой тревожностью. Так что общая повышенная чувствительность до угроза + состоит из низкой тревожности — восприятия и высокой тревожности = моторных частей.
Благодарим рецензента за положительный отзыв. Мы согласны с рецензентом в том, что усиление сенсорной и моторной реакции сильно модулируется, вдвойне диссоциативным образом, оценкой тревожности участников. Тем не менее, мы считаем, что средняя часть раздела «Результаты» важна для данной области, поскольку она проливает свет на специфический нейронный процесс, ответственный за улучшения, зависящие от угрозы: повышенная нейронная чувствительность к эмоциям, сигнализирующим об угрозе, а не к восприятию или принятию решения. предвзятость, которая подтверждает наш основанный на модели анализ поведенческих данных и которая, насколько нам известно, ранее не проверялась. Более поздняя диссоциация между участниками с высокой и низкой тревожностью основывается на этих первых выводах, подчеркивая избирательный вклад сенсорных и моторных областей в повышенную чувствительность к эмоциям, сигнализирующим об угрозе, в двух группах с высокой и низкой тревожностью.
Чтобы сделать наш анализ ЭЭГ более четким, как указано в коллективных ответах выше, мы последовали совету рецензента, смешав описание методов с основным текстом. Мы стремились сделать результаты понятными для неспециалистов, не требуя подробного чтения раздела «Методы». Мы надеемся, что рецензент согласится с тем, что эти изменения значительно улучшили читабельность рукописи для неспециалистов.
Рецензент №2:
В целом, я думаю, что это хорошая работа, и авторы явно приложили значительные усилия, чтобы предоставить полный отчет о своем наборе данных, и им следует приветствовать объем проведенного анализа. чтобы лучше понять нейронную реакцию на угрожающие социальные стимулы. Однако я обнаружил, что часть анализа трудно разобрать, а в некоторых случаях важная информация отсутствовала (или была неочевидной). Особенно меня беспокоило выборочное представление поведенческих результатов.
Мы благодарим рецензента за положительную оценку нашей аналитической основы и результатов. Мы понимаем озабоченность рецензента по поводу нашего выборочного описания поведенческих результатов. Первоначально мы решили отдать предпочтение краткости, а не полноте поведенческих результатов, чтобы облегчить передачу основного сообщения, но ретроспективно мы можем видеть, как недостающие детали могут ухудшить читабельность этого раздела. Мы последовали предложению рецензента, предоставив недостающую информацию о точности распознавания и времени реакции в исправленной рукописи.
Где находятся результаты времени реакции (как уже упоминалось, они были проанализированы в методах)?
Результаты времени реакции были добавлены в раздел «Результаты»:
«Анализ времени реакции (ВР) выявил снижение правильного ВУ с силой эмоции (повторные измерения ANOVA, F 6,138 = 54,5, p < 0,001) , более быстрые реакции на сердитые лица по сравнению с испуганными (F 1,23 = 12, p < 0,01) и более быстрые реакции на прямой по сравнению с отведенным взглядом (F 1,23 = 7,7, р < 0,05). Кроме того, наблюдалась эмоция при взаимодействии взглядом (F 1,23 = 8, p < 0,01), что соответствует более быстрому времени реакции на прямой по сравнению с отведенным взглядом только в состоянии гнева (t 23 = -3,9, р < 0,001)».
Обратите внимание, что эти результаты времени реакции обеспечивают независимое повторение нашего основного вывода о взаимодействии между направлением взгляда и эмоциями в отношении точности распознавания эмоций — показывая ту же закономерность в отношении скорости распознавания эмоций.
Отсутствовало ли взаимодействие между производительностью категоризации и уровнем морфинга при изучении взаимодействия производительности взгляд-эмоция? Графики на рис. 2В показывают, что, как и следовало ожидать, эффект взгляда сильнее для более слабых сигналов эмоций (ближе к нейтральным).
Действительно существует взаимодействие между угрозой+/- и уровнем трансформации при категоризации, и эти результаты были добавлены в раздел «Результаты»:
6,138 = 4,3, p < 0,01), что объясняется более сильным влиянием взгляда на категоризацию эмоций при слабой силе эмоции (взгляд по взаимодействию эмоций для уровней 1–4, F 1,23 = 23,8, p < 0,001), чем при высокой силе эмоций (взгляд по взаимодействию эмоций для уровней с 5 по 7, F 1,23 = 5,1, p < 0,05)».
Тем не менее, обратите внимание, что это взаимодействие само по себе не является специфическим ни для одной из наших двух моделей (разница между предвзятостью и чувствительностью между условиями угроза+ и угроза-). Разница между двумя моделями заключается в том, является ли эффект взгляда самым сильным для безэмоциональных/нейтральных стимулов (эффект предвзятости) или для слабых эмоций (эффект чувствительности, см. новый рисунок 1, показывающий предсказания модели для обоих типов эффектов). Это наблюдение было одной из основных причин для анализа на основе моделей, который может быть арбитром между этими двумя альтернативными объяснениями этого взаимодействия при категоризации эмоций.
В качестве более второстепенного вопроса, почему участники не на 100% правы в отношении конечных точек морфов (100% гнева, 100% страха) — некоторые личности постоянно классифицируются неправильно?
Это очень хороший момент, и, к сожалению, мы не объяснили, как мы учитывали это наблюдение при моделировании поведения участников. В каждую модель, которую мы подогнали к выборам участников, мы добавили дополнительный параметр «промежуток», который соответствует доле случайных предположений среди вариантов. Этот параметр обычно используется в психофизике для моделирования несовершенных асимптотических характеристик, отмеченных рецензентом. Мы исправили раздел «Методы», чтобы описать полную модель, включая параметр «промежуток»:
«Мы начали с простейшей модели (модель 0), которая могла бы объяснить решения каждого испытуемого, используя зашумленную, похожую на «обнаружение сигнала» психометрическую модель, в которую мы включили скорость отклонения, тем самым учитывая, что испытуемые угадывали случайным образом определенную пропорцию число испытаний:
P(гнев) = Ф[w* x + b]*(1− ɛ)+ 0,5*ɛ
, где P(гнев) соответствует вероятности оценки лица как гневного, Ф[ .] к кумулятивной нормальной функции, w к перцептивной чувствительности к отображаемой эмоции, x до пробного массива значений доказательств в пользу гнева или страха (сила эмоции, от -7 для сильного выражения страха до +7 для сильного выражения гнева), b до аддитивного, независимого от стимула предвзятость к одной из двух эмоций, а ε — доля случайных предположений среди вариантов».
Обратите внимание, что среднее значение на уровне группы для скорости отклонения ɛ, подобранное одновременно с другими параметрами w и b, равно 16,2 ± 1,8% (среднее ± стандартная ошибка среднего), значение, которое, что важно, не различается между условиями угрозы+ и угрозой- ( t 23 = 0,4, p > 0,5).
Тем не менее, мы серьезно отнеслись к альтернативной гипотезе рецензента и провели дополнительный анализ, чтобы исключить возможность того, что некоторые личности постоянно неправильно классифицировались участниками. Во-первых, мы подтвердили, что все идентичности, использованные в исследовании, были классифицированы значительно выше случайных на обоих крайних точках оси морфинга эмоций. Так оно и было на самом деле (парные t-тесты, все t 23 > 6,4, все p < 0,001). Во-вторых, мы установили отдельные сигмовидные функции, чтобы описать кривую среднего психометрического ответа участников для каждой личности, взятой отдельно. Этот анализ «фиксированных эффектов» подтвердил, что все идентичности были последовательно отнесены к гневу и страху в зависимости от уровня морфа, о чем свидетельствуют значительно положительные наклоны связанных психометрических функций (логистическая регрессия, все t -значения> 12,8 , все р < 0,001). Кроме того, распределение точки субъективной «нейтральности» (при которой психометрическая функция пересекает 0,5) по идентичностям сосредоточено на 0,01 (по шкале от -1 для самого сильного выражения страха до +1 для самого сильного выражения гнева). со стандартным отклонением 0,20, что указывает на отсутствие крайних предубеждений в категоризации участников для идентичности, использованной в исследовании. Вместе мы считаем, что эти дополнительные результаты полностью подтверждают наш выбор моделировать несовершенные асимптотические характеристики с помощью независимых от стимулов промахов, а не неправильно распознанных эмоций, проявляемых определенными личностями.
Переходим к анализу ЭЭГ. Авторы используют передовые и относительно специализированные методы для изучения взаимосвязи сигнала ЭЭГ с модуляциями эмоциональной силы, и их результаты интересны и кажутся убедительными. Хотя я думаю, что они проделали разумную работу по объяснению этих методов и того, как их следует интерпретировать, мне хотелось бы получить более подробную информацию о статистических тестах, которые они использовали. Конечно, они выполнили некоторую загрузку, чтобы исправить проблемы с множественными сравнениями, с которыми они сталкиваются. На это намекает подпись к рисунку 3 (пороговое значение, скорректированное по кластеру), но никаких подробностей, которые я могу видеть, нигде не приводится.
Как указано в нашем ответе на коллективные вопросы выше, мы добавили параграф «статистические процедуры» в раздел «Методы», где мы более подробно описываем статистический анализ данных. Что касается проблемы множественных сравнений, мы применили обычные поправки, описанные в (Maris and Oostenveld, 2006), отраженные в «скорректированных по кластеру» p-значениях, представленных в разделе «Результаты». Теперь мы прямо указываем, как учитывались множественные исправления, в разделе «Методы» пересмотренной рукописи в новом подразделе «статистические процедуры» («Все регрессионные анализы […] соответствуют его «кластерно-скорректированному» p-значению»). .
Кроме того, я думаю, что авторы использовали все испытания (правильные и неправильные) в своем анализе ЭЭГ, но мне неясно, почему это было бы оправдано, особенно для более слабых эмоциональных стимулов (где участники верны только примерно в 70% случаев). при их классификации). Например, испытание угроза + предотвращение страха могло быть классифицировано участником как лицо гнева (и, таким образом, обработано участником как угроза- ). Ожидали ли авторы, что нейронное кодирование будет одинаковым в обоих случаях? Если нет, то как они могут оправдать неучет ответов участников в своем анализе ЭЭГ?
Как утверждалось в нашем ответе на коллективные вопросы выше, важно помнить, что согласно теориям принятия решений, вдохновленным классической теорией обнаружения сигналов, процесс принятия решений, лежащий в основе правильных ответов и ошибок, одинаков — единственная разница заключается в положении зашумленная переменная решения по отношению к критерию принятия решения в конкретных испытаниях. Таким образом, определение чувствительности и предвзятости основывается как на правильных ответах, так и на ошибках. Исходя из этого, мы решили провести регрессионный анализ данных ЭЭГ без искусственного отделения правильных ответов от ошибок. Таким образом, результирующий паттерн отражает усиление нейронной чувствительности к эмоциям, сигнализирующим об угрозе, независимо от точности последующей реакции.
Тем не менее, мы не имеем в виду, что нейронное кодирование силы эмоции должно быть одинаковым при анализе отдельно правильных ответов и ошибок. И, как и ожидалось, нейронное кодирование силы эмоций сигналами ЭЭГ на височных, центрально-теменных и двигательных электродах не выявило каких-либо значимых кластеров при рассмотрении ошибок по отдельности (с поправкой на кластер p > 0,1). Напротив, рассмотрение только правильных испытаний в анализе кодирования, показанном на рис. 4, не изменило значимых различий между условиями «угроза+» и «угроза-» на височных электродах через 170 мс после начала стимула (9). 0141 T 23 = -2,1, P <0,05), теменные электроды при 500 мс ( T 23 = 4,2, P <0,001) и моторные сигналы на 200 м. = 3, р < 0,01).
Кроме того, обратите внимание, что корреляционный анализ нейронного выбора, выполненный в разделе «Результаты», действительно учитывает ответы участников, и теперь это более явно подчеркивается в исправленной рукописи. Важным преимуществом примененного нами модельного подхода по сравнению с безмодельным разделением правильных и ошибочных ответов является то, что он позволяет оценить не только , изменяются ли сигналы ЭЭГ с поведенческими реакциями, а также как они это делают: через изменения чувствительности и/или смещения.
Наконец, мне понравился анализ зависимости от тревоги, и его результаты ясны и интересны.
Мы рады, что рецензент счел наш анализ нейронных эффектов, зависящих от тревоги, важным и интересным.
Рецензент №3:
[…] Эта часть статьи элегантна, хорошо сделана и, насколько мне известно, оригинальна. Раздел «Методы» действительно ясен — я бы предпочел, чтобы некоторые объяснения моделей были перенесены в основную часть документа, поскольку эта часть, хотя и является «технической», действительно необходима для понимания смысла последующего анализа. и в его нынешнем виде введение гораздо менее ясно, чем более подробное объяснение.
Мы благодарим рецензента за его положительные комментарии к исследованию. Как указано в ответ на вышеприведенные коллективные замечания, мы попытались более подробно рассказать о методах, основанных на регрессии и управляемых моделями, которые мы применяли к нейронным данным в разделе «Результаты» пересмотренной рукописи. Мы надеемся, что рецензент найдет переработанную рукопись более ясной в этом отношении.
В заключительном разделе статьи утверждается связь с уровнем тревожности у отдельных субъектов, как угроза+ и — по-разному модулируют височные области у лиц с низкой тревожностью), а угроза+ модулирует моторные области у лиц с высокой тревожностью.
У меня есть оговорки по поводу второй части диссоциации у лиц с высокой тревожностью, в основном связанной с рисунком 7Б – мне кажется, это главная слабость.
Мы считаем, что прояснили и усилили наблюдаемую зависимую от тревоги диссоциацию между сенсорными и моторными улучшениями в ответ на угрозу + комбинации взгляда и эмоций в исправленной рукописи — как указано в ответ на общие пункты выше и конкретные вопросы, поднятые рецензент ниже.
1) Тревога сочетается с депрессией и социальной тревожностью. Были ли участники проверены на предмет оценки других факторов, которые могут быть связаны с тревожностью?
Участники не подвергались скринингу на другие параметры, такие как социальная тревожность или депрессия, а только на один аспект, относительно которого у нас были прогнозы относительно восприятия угрозы: тревожность. Рецензент прав в том, что показатели тревожности, измеренные с помощью опросника STAI, обычно коррелируют с показателями депрессии и социальной тревожности у разных людей, и мы обновили раздел «Обсуждение», чтобы признать вытекающее из этого ограничение специфичности наблюдаемых эффектов: «Дальнейшие исследования должны оценить специфичность этих зависимых от тревоги эффектов в свете растущих данных в пользу коморбидности между тревогой и депрессивными расстройствами».
Тем не менее, как было указано в ответ на вышеизложенные коллективные вопросы и в отношении сопутствующей патологии между депрессией и тревогой, было показано, что у пациентов с клинически выраженной тревогой и депрессией уровень смертности был ниже, чем у пациентов только с депрессией (Mykletun et al., 2009). ). Таким образом, это открытие предполагает, что клиническая тревога может иметь специфическую «защитную» функцию, связанную с адаптивной ролью тревоги в выживании, которая не является общей для депрессии. Несмотря на свои ограничения (общие для большинства существующих исследований тревоги в этой области), наши результаты предоставляют эмпирические доказательства в пользу такой положительной функции неклинической тревоги в общей популяции.
2) Все участники, кроме одного, лежат в самом центре нормального диапазона. На рисунке 7B похоже, что оценка 40+ может быть выбросом. Значения R вычисляются с помощью Пирсона (я предполагаю, хотя я заметил, что этого нет на рисунке; это должно быть). Если удаленный субъект исключен, сохраняется ли корреляция?
Силы корреляции, указанные в рукописи, действительно являются значениями линейной корреляции Пирсона, и это теперь указано в подписях к тексту и рисункам исправленной рукописи. Как было указано в ответ на коллективные замечания выше, мы провели несколько дополнительных анализов данных, показанных на рис. 7 (теперь рис. 8 в исправленной рукописи), которые должны убедить рецензента в надежности зависимых от тревоги эффектов в сенсорных и двигательные области.
Во-первых, важно отметить, что показатели тревожности наших участников (от 20 до 45) находятся в пределах диапазона нормальной популяции (< 60), и поэтому нет априорной причины исключать кого-либо из наших участников. исключительно на основе его/ее оценки тревожности. Кроме того, именно вариабельность показателей тревожности между участниками обусловила их корреляцию с нервными эффектами. В качестве первого контрольного анализа мы рассчитали интервалы прогнозирования для линии регрессии на уровне группы по показателю тревожности каждого участника. Ни один участник не попал за пределы 95% интервалов прогнозирования как для височной (рис. 8А), так и для двигательной области (рис. 8В) — и, таким образом, могут быть статистически помечены как выбросы. Тем не менее, если исключить 90 141 произвольно 90 142 участника № 4, чей показатель тревожности равнялся 45, моторная корреляция с тревожностью остается минимально значимой (90 141 r 90 142 = 0,35, d.f. = 21, двусторонний 90 141 p 90 142 = 0,09), а также взаимодействие между состоянием тревожности и уровнем угрозы (смешанный дизайн ANOVA, F 1,22 = 3,1, p = 0,09). А у высокотревожных участников с разбивкой по медиане нейронное кодирование силы эмоции в состоянии угроза+ остается значимым на двустороннем уровне (t-тест против нуля, t 10 = 2,4, двусторонний p = 0,03), а также наблюдаемая разница между условиями угроза+ и угроза- (парный t-критерий, t 10 = 2,6, двусторонний p = 0,02).
Вместо того, чтобы произвольно исключить одного участника, мы выполнили процедуру перекрестной проверки с исключением одного, в которой мы вычислили «перекрестные» интервалы прогнозирования для линии регрессии на уровне группы по показателю тревожности участника № n, когда данные для участника № n были исключены из регрессии на уровне группы. Эта процедура была повторена для 24 участников для корреляции тревоги и угрозы в двигательных областях и показала, что нервные эффекты у двух участников немного снизились (<20%) за пределами 9.0141 перекрестная проверка 95% интервалов прогнозирования, включая участника №4. Тем не менее, пересчет корреляции на уровне группы в двигательных областях после исключения двух выбросов, оставив в анализе 22 участника, привел к значимому эффекту ( r = 0,48, d.f. = 20, p = 0,02). Этот исключающий анализ не выявил каких-либо отклонений для корреляции тревоги и угрозы в височных областях.
Вместе мы считаем, что эти контрольные анализы подтверждают надежность корреляций тревоги и угрозы как для височных, так и для двигательных областей, и теперь мы опишем их в разделе «Методы» пересмотренной рукописи.
Исходя из этого, мне кажется, что наблюдается увеличение изменчивости в группе «более тревожных», что может быть движущей силой корреляции, тем более, что похоже, что разница между угрозой и отсутствием угрозы в высокой тревожная группа в 7B не имеет значения.
В ответ на это конкретное замечание мы можем заверить рецензента, что существует значительная разница в нейронном кодировании моторной латерализации для группы с высокой тревожностью между состояниями «угроза+» и «угроза-» (парный t-тест, т 11 = 3,0, р = 0,01). Нейронное кодирование состояния угрозы+ даже значительно положительное (t-тест против нуля, t 11 = 2,6, p = 0,02). Обратите также внимание, что оба теста остаются значимыми, даже когда участник № 4 произвольно исключается из группы с высокой тревожностью (см. наш ответ на предыдущий пункт). Наконец, использование процедуры исключения одного для выявления двух выбросов в отношении корреляции, показанной на рисунке 7B (теперь рисунок 8B), оставило корреляцию значимой, тем самым подтвердив, что корреляция не определяется посторонними точками данных.
3) Эффект на рисунке 7B кажется слабым эффектом в один момент времени. Нет обоснования/объяснения выбора ни окна в 7А, ни одиночной точки в 7Б (если это так).
Мы полностью согласны с рецензентом в этом вопросе. В то время как мы решили измерить корреляцию с тревогой в момент времени, соответствующий пику эффекта кодирования в двигательных сигналах (200 мс), мы выбрали временное окно, соответствующее значимому кластеру для кодирования на височных электродах. Это действительно было несоответствие, которое мы исправили, также выбрав пик нейронного кодирования для височных электродов (280 мс) для выполнения корреляции. Мы соответственно изменили рисунок (ранее рисунок 7, теперь рисунок 8) и текст:
«Действительно, ситуационная тревожность значительно влияла на нейронное кодирование […] между комбинациями угроза + и угроза- оказалась значимой (коэффициент корреляции Пирсона r = 0,51, df = 22, p = 0,01; рисунок 8A)».
Обратите внимание, что обе корреляции с тревожностью остаются значимыми, если брать все временное окно, соответствующее значимому кластеру, как в височной (170-400 мс, r = 0,53, d.f. = 22, p < 0,01), так и в моторной электроды (130-300 мс, r = 0,43, ф. р. = 22, р < 0,05). Мы считаем, что эти дополнительные анализы еще больше усиливают корреляции, зависящие от тревоги, о которых сообщается в рукописи.
4) Роль времени реакции: авторы говорят, что не обнаружили разницы в поведенческих реакциях между более и менее тревожными группами. Это включало время реакции?
Анализы действительно включали время реакции, которое не различается между группами с высокой и низкой тревогой и не коррелирует с тревогой. Не было измеримой разницы между двумя тревожными группами только с точки зрения поведения. В разделе «Результаты» сообщается об одном зависимом от тревоги эффекте, который значительно влияет на поведение в нашем исследовании. Сила нейронного кодирования эмоций «угроза+» на двигательных электродах значительно коррелировала с поведенческой чувствительностью у участников с высокой тревожностью, но не у участников с низкой тревожностью, и значительно больше у участников с высокой тревожностью, чем у участников с низкой тревожностью:
«Более того, нейронное кодирование эмоций угрозы+ в моторных сигналах коррелировало с поведенческой чувствительностью к эмоциям угрозы+ у сильно тревожных людей (коэффициент корреляции Пирсона r = 0,66, d. f. = 10, p = 0,01; рисунок 8B), в то время как это не было случай для малотревожных лиц (коэффициент корреляции Пирсона r = -0,42, df = 10, p > 0,16, разница между коэффициентами, p <0,01)».
[Примечание редактора: перед принятием были запрошены дополнительные изменения, как описано ниже.]
Рисунок 1 очень полезен кроме Я не понимал ось эмоций на рисунке 1 (закрашена серой областью), пока не увидел нижнюю часть рисунка 3, где она обозначена как эмоциональная чувствительность. Пожалуйста, обозначьте на рисунке 1 и поясните в легенде (в дополнение к пояснению в тексте).
Приносим извинения за то, что не уточнили, что изображено на рис. 1, закрашенное серым цветом. Ось эмоций представляет свидетельство эмоционального выражения от самого испуганного лица слева к самому злому лицу справа. Заполненная серая область представляет собой разницу между двумя построенными кривыми (одна для прямого взгляда, другая для отведенного взгляда). Мы добавили эту серую область, чтобы подчеркнуть тот факт, что различия между прямым и отведенным взглядом (т. е. разница между двумя кривыми) будут максимальными для нейтрального уровня для изменения смещения между условиями прямого и отведенного взгляда и максимальны для низких эмоций. сильные стороны для изменения чувствительности между состояниями угроза+ и угроза-. Теперь мы добавили к рисунку серую рамку, которая описывает заполненную серую область: «разница прямая/предотвращенная», и мы добавили в легенду:
«Максимальные эффекты проявляются для нейтральных (безэмоциональных) выражений, выделенных заштрихованной серой областью на оси эмоций, которая представляет разницу между двумя психометрическими функциями для прямого и отведенного взгляда.»
Как были выбраны моменты времени на рисунке 4? Время в B и C имеет для меня некоторый смысл, но я не понимаю, как было выбрано 280, когда оно так далеко от области существенных различий. Также, пожалуйста, объясните, почему карты в D-F не выглядят так, как если бы они были получены из мозга, нанесенного на карту в середине A-C (особенно B-C). Перечитав это, я думаю, что понимаю проблему синхронизации — вы отображаете отклонения от 0, а не пиковые различия между условиями. Может быть, сделать акцент на последнем так же, как и на первом, превратив серые прямоугольники в столбцы вверху, чтобы они соответствовали зеленым и оранжевым столбцам? Все одинаково подчеркнули именно так. Путаница в отношении выравнивания карты или его отсутствия остается.
Топографии ЭЭГ, представленные в середине рис. 4a-c, взяты при латентности пиковых отклонений от нуля, а не пиковых различий между состояниями. В то время как первый пик кодирования независимо от условий появляется через 280 мс, разница между двумя интересующими нас условиями (угроза+ и угроза-) проявляется раньше (170 мс). Это не относится к двум другим пикам (500 мс и время отклика), поскольку разница между условиями соответствовала отклонениям пиков от нуля. Мы наносим эти топографии, чтобы обосновать, как мы выбрали электроды, на которых мы сравниваем два интересующих условия, т. Е. Используя контраст (t-критерий против нуля), ортогональный контрасту между двумя условиями. Теперь мы добавили к рисунку 4 поверх этих топографий: «Пиковое кодирование в разных условиях».
Более того, по предложению редактора мы заменили серые прямоугольники на серые столбцы поверх зеленых и оранжевых столбцов, чтобы в равной степени подчеркнуть отклонения от нуля и различия между условиями (пересмотренный рисунок 4). Тем не менее, наша главная цель состояла в том, чтобы рассмотреть когнитивные и нейронные различия между интересующими нас состояниями (угроза+ и угроза-), а не изучать кодирование эмоций как таковое. Поэтому мы по-прежнему считаем, что было бы полезно визуально различать эти два типа статистических различий, уделяя больше внимания разнице между условиями «угроза+» и «угроза-».
Карты мозга, представленные на рис. 4d-f, представляют разницу в кодировании между состояниями «угроза+» и «угроза-» на исходном уровне в моменты времени, когда между этими состояниями существовала значительная разница (серая полоса). Теперь мы добавили к рисунку 4 поверх этих карт мозга: «Источник различий в кодировании T+/T-». Анализ реконструкции источника локализует области мозга, которые генерируют сигналы ЭЭГ, записанные на скальпе. Таким образом, аналогичные усредненные сигналы ЭЭГ, записанные на скальпе, могут исходить из разных источников мозга. Соответственно, было показано, что височные компоненты ЭЭГ (N170) имеют разные источники в головном мозге, включая веретенообразную извилину (например, Itier and Taylor, 2002) и верхнюю височную борозду (например, Batty and Taylor, 2003). Здесь мы сообщаем об обоих этих регионах как об источниках различий между кодированием угрозы+ и угрозы-. Кроме того, мозговые источники теменной ЭЭГ-активности (P300) включают широко разнесенные области мозга, в пределах которых лобные и височные области (Nieuwenhuis et al., 2005), подобно тем, о которых сообщалось в настоящем исследовании при вычислении источников различий между угроза+ и угроза- связанный с теменной скальп-записанной ЭЭГ. Наконец, обнаружение дорсальных центральных двигательных областей в f (связано с топографией в c во время ответа), а не e (связано с топографией в b через 500 мс) неудивительно во время ответа.
Мы также добавили это предложение в раздел «Результаты»:
«Чтобы оценить, какие области мозга генерируют сигналы ЭЭГ, записанные на скальпе, мы вычислили корковые источники этого усиленного кодирования эмоций, сигнализирующих об угрозе, выполнив тот же регрессионный подход к минимуму. -нормовые текущие оценки, распределенные по поверхности коры».
Не могли бы вы для общего читателя добавить несколько слов о различиях в латерализации, показанных на рисунке 6B и обсуждавшихся в подразделе «Раннее нейронное кодирование эмоций, сигнализирующих об угрозе, при двигательной подготовке»? Это кажется устойчивым отличием — известно ли оно и ожидается ли оно? Удивительно в любом случае? Помните, что eLife Читательская аудитория широка и не обязательно хорошо осведомлена о большом объеме информации внутри отрасли.
Следуя совету редактора, мы добавили некоторую информацию о различиях в латерализации, показанных на рис. 6В, в подразделе «Раннее нейронное кодирование эмоций, сигнализирующих об угрозе, при двигательной подготовке»:
«Выполнение и подготовка движений конечностей совпадают с подавлением низкочастотная (8-32 Гц) активность, которая сильнее в моторной коре, контралатеральной по сравнению с ипсилатеральной по отношению к движению. Таким образом, ожидается, что вычитание контралатеральной активности из ипсилатеральной моторной коры приведет к положительному показателю двигательной подготовки».
https://doi.org/10.7554/eLife.10274.012
Статья и информация об авторе
Сведения об авторе
Marwa El Zein
- Laboratoire de Neurosciences Cognitives, Département d’Etudes Cognitives, Ecole Normale Supérieure, Исследовательский университет PSL, Париж, Франция
- Университет Пьера и Марии Кюри, Париж, Франция
Взнос
МЭЗ, Концепция и дизайн, Сбор данных, Анализ и интерпретация данных, Составление или редактирование статьиДля корреспонденции
marwaelzein@gmail. comКонкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.Валентин Вайарт
Laboratoire de Neurosciences Cognitives, Департамент когнитивных исследований, Ecole Normale Supérieure, Исследовательский университет PSL, Париж, ФранцияВклад
VW, Концепция и дизайн, Анализ и интерпретация данных, Составление или редактирование статьиВклад в равной степени с
Жюли ГрезКонкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов. «Этот ORCID iD идентифицирует автора этой статьи:» 0000-0001-6522-7837Жюли Грез
Laboratoire de Neurosciences Cognitives, Департамент когнитивных исследований, Ecole Normale Supérieure, Исследовательский университет PSL, Париж, ФранцияВклад
JG, Концепция и дизайн, Анализ и интерпретация данных, Составление или редактирование статьиВклад в равной степени с
Валентин ВайартКонкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Финансирование
Национальное агентство исследований (ANR-11-EMCO-00902)
- Жюли Грез
Национальное агентство исследований (ANR-11-0001-02 PSL*)
- Валентин Вайарт
- Жюли Грез
Национальное агентство исследований (ANR-10-LABX-0087)
- Валентин Вайарт
- Жюли Грез
Фонд Роджера де Сполберха
- Жюли Грез
Национальный институт здравоохранения и медицинских исследований
- Валентин Вайарт
- Жюли Грез
Спонсоры не участвовали в разработке исследования, сборе и интерпретации данных или принятии решения о представлении работы для публикации.
Благодарности
Эта работа была поддержана грантами французского национального исследовательского агентства ANR-11-EMCO-00902, ANR-11-0001-02 PSL*, ANR-10-LABX-0087, Фонда ROGER DE SPOELBERCH и INSERM. Мы хотим поблагодарить Laurent Hugueville за его полезную техническую помощь.
Этика
Субъекты-люди: все участники предоставили письменное информированное согласие в соответствии с институциональными рекомендациями местного комитета по этике исследований (в соответствии с Хельсинкской декларацией) и получили оплату за свое участие. В этом согласии они согласились, что их данные будут храниться анонимно, анализироваться и что соответствующие результаты будут представлены на конференциях и опубликованы в рецензируемых журналах. Этическое одобрение было предоставлено местным комитетом по этике (Comite de Protection des Personnes, Ile-de-France VI, одобрение Inserm № C07-28, одобрение DGS № 2007-0569)., одобрение IDRCB № 2007-A01125-48), в соответствии со статьей L1121-1 Кодекса общественного здравоохранения Франции.
Редактор-рецензент
- Пегги Мейсон, Чикагский университет, США
История публикаций
- Поступило: 22 июля 2015 г.
- Принято: 25 ноября 2015 г.
- Версия записи опубликована: 29 декабря 2015 г. (версия 1)
Авторское право
© 2015, Эль Зейн и др.
Эта статья распространяется в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование и распространение при условии указания автора и источника.
Метрики
Счетчик цитирования статей, полученный путем опроса самого высокого числа из следующих источников: Crossref, PubMed Central, Scopus.
Ссылки для скачивания
Список ссылок, состоящий из двух частей, для загрузки статьи или частей статьи в различных форматах.
Загрузки (ссылка для скачивания статьи в формате PDF)
- Статья PDF
Открытые цитаты (ссылки для открытия цитат из этой статьи в различных онлайн-сервисах управления ссылками)
- Менделей
- ЧитатьКуб»>
Процитировать эту статью (ссылки для загрузки цитат из этой статьи в форматах, совместимых с различными инструментами управления ссылками)
- Марва Эль Зейн
- Валентин Вайарт
- Жюли Грез
(2015)
Тревога диссоциирует адаптивные функции усиления сенсорной и двигательной реакции на социальные угрозы
eLife 4 :e10274.